Строение листа дерева: Состав листа дерева – Стройка и Ремонт
Лист дерева Строение функции и виды листьев
Лист дерева. Строение, функции и виды листьев.
В ботанике листья являются неотъемлемой частью стволовой системы растения. Лист дерева состоит из листовой пластинки (уплощенная часть листа), черешка (стебель) и прилистников (придатки у основания листа). Листья на деревьях бывают различных форм и размеров. Все крона дерева должна занимать достаточно большую площадь поверхности дерева, это важно для поглощения света хлорофиллом в процессе фотосинтеза и углекислого газа (СО2) для производства органических молекул.
Такие разные листья
Как правило, лист дерева состоит из широкой лопасти (пластинки), прикрепленной к стеблю. Листья бывают разные по размеру, форме и некоторым другим характеристикам, включая тип жилкования (расположение вен). Разные типы жилкования характерны для разных видов растений, например, двудольные имеют сетчатое жилкование, у однодольных растений жилкование листьев параллельное. Листья также могут быть простые и составные.
Строение и функции листа
Листья деревьев выполняют ряд важнейших функций, а также содержат воду, которая необходима для преобразования энергии света в глюкозу в процессе фотосинтеза. Листья имеют две структуры, которые сводят к минимуму потери воды — кутикулу и устьица. Кутикула является восковым налетом на верхней и нижней части листьев, которая предохраняет воду от испарения в атмосферу.
Основная функция листа – это производства продуктов питания для растения путем фотосинтеза. Хлорофилл, вещество, которое придает растениям их характерный зеленый цвет, поглощает световую энергию. Внутреннее строение листа находится под защитой эпидермиса. Центральный лист, или мезофилл, состоит из мягкой стенки, его клетки известны как паренхимы. На одну пятую часть мезофилл состоит из хлорофилла, содержащего хлоропласты. Они поглощают солнечный свет, чтобы выделять затем кислород, и, в сочетании с определенными ферментами, добывают из воды водород.
Кислород, освобожденный из зеленых листьев, используется для дыхания растений и животных. Водород, полученный из воды, в сочетании с углекислым газом участвует в ферментативных процессах фотосинтеза в виде сахаров, которые являются основой растительного и животного мира. Кислород попадает в атмосферу через специальные поры на поверхности листьев.
Водород, полученный из воды, в сочетании с углекислым газом участвует в ферментативных процессах фотосинтеза в виде сахаров, которые являются основой растительного и животного мира. Кислород попадает в атмосферу через специальные поры на поверхности листьев.
Хотя кутикула выполняет важную функцию защиты от чрезмерной потери влаги, листья не могут быть непроницаемыми, потому что они должны также позволить впитываться углекислому газу. После того как CO2 проникает в лист через устьица, он перемещается в клетки мезофилла, где и происходит фотосинтез с последующим производством глюкозы.
От чего зависит цвет листьев?
За цвет отвечают хлорофиллы, зеленые пигменты, которые обычно присутствуют в гораздо большем количестве, чем другие. Осенью производство хлорофилла замедляется, так как дни становятся короче и прохладнее. Постепенно хлорофилл разрушается и исчезает, и начинают проявляться цвета других пигментов. Они включают каротин (желтый), ксантофилл (бледно-желтый), антоцианин (красный, сине-фиолетовый) и бетацианин (красный). Танины придают например, дубовым листьям их темно-коричневый оттенок.
Танины придают например, дубовым листьям их темно-коричневый оттенок.
Жизнь листа
Лист дерева является в основном короткоживущей структурой. Даже когда они сохраняются в течение двух или трех лет, например, хвойные и широколиственные вечнозеленые растения, то после первого года приносят всему дереву не такую большую пользу, как в начале. Опадать начинают листья у основания черешка листа. Обычно это происходит осенью, хотя на этот естественный биологический процесс могут оказывать влияние и другие факторы, например, опадение может быть обусловлено повреждениями, связанными с насекомыми, болезнями или засухой.
Ближе к осени лист дерева претерпевает некоторые возрастные изменения, так как дни становятся все короче, а солнечного света все меньше. В результате зона черешка начинает смягчаться до тех пор, пока листик не отпадет. На стебле образуется целительный слой, который затягивает рану, оставляя своеобразный шрам.
Составные части листа
Основной лист покрытосеменных растений состоит из основания листьев, прилистников, черешка, и лезвия (пластины). Основание листьев слегка расширено в том месте, где лист прикрепляется к стеблю. Парные прилистники, при их наличии, находятся на каждой стороне листа основания и напоминают чешуйки, колючки, или структуры, напоминающие сам лист. Черешок представляет собой стебель, который соединяет лезвие с основанием листьев. Лезвие является основной фотосинтетической поверхностью растения.
Основание листьев слегка расширено в том месте, где лист прикрепляется к стеблю. Парные прилистники, при их наличии, находятся на каждой стороне листа основания и напоминают чешуйки, колючки, или структуры, напоминающие сам лист. Черешок представляет собой стебель, который соединяет лезвие с основанием листьев. Лезвие является основной фотосинтетической поверхностью растения.
Виды и формы листьев
Форма листьев деревьев может быть различной. В природе могут встречаться простые и сложные листья. Когда только одно лезвие соединено с черешком, то лист называется простым, он также, в свою очередь, может быть рассечен по краям самыми разными способами. Такие листья могут быть целые и ровные, а также они могут иметь зубчатые или пильчатые поля. Также края могут быть закругленные или фестончатые. Большое разнообразие встречаются на вершине и в основании листа. Есть листья, у которых нет черешка и они прикрепляются непосредственно к стеблю, а некоторые листья могут не иметь прилистников.
По типу расположения виды листьев деревьев можно выделить следующие: очередные, парные (напротив) и мутовчатые. При очередном расположении листья равномерно распределяются по стеблю, попеременно образуя восходящую спираль. В парном расположении листья растения находятся друг напротив друга. Растение имеет мутовчатый тип расположения, когда три или более листьев исходят из одного узла.
Иголки – это тоже листья
Форма листьев является основным инструментом для идентификации видов растений. Хвойные виды растений, такие как ель, пихта, и сосна, произрастающие в холодных условиях, имеют листья в виде иголок. Игловидные листья помогают в снижении потерь воды. В жарком климате такие растения, как кактусы, имеют суккулентные листья, которые также помогают экономить воду. Многие водные растения имеют листья с широкой пластинкой, плавающей на поверхности воды, при этом густая воскообразная кутикула на поверхности листьев отталкивает воду.
Касательно распределения растений на Земле климат является определяющим фактором, именно поэтому зоны растительности почти всегда соответствуют климатическим зонам. От особенностей климата и окружающей среды в полной мере зависит многообразие видов и форм растительности. Листья, которые в первую очередь являются фотосинтетические органами, также приспосабливаются к климатическим условиям наиболее оптимальным способом.
От особенностей климата и окружающей среды в полной мере зависит многообразие видов и форм растительности. Листья, которые в первую очередь являются фотосинтетические органами, также приспосабливаются к климатическим условиям наиболее оптимальным способом.
Источник
Название деревьев и их листья
Каждый день мы с детьми бываем на улице, гуляем в парках, проходим мимо самых разнообразных деревьев, собираем букеты из листьев. Но порой возникает ситуация, даже у взрослых, когда не можем определить тот или иной листик с какого дерева. Предлагаем вашему вниманию подборку материалов с картинками деревьев, а также их листьев, плодов. Этот материал поможет при подготовке к урокам по окружающему миру. Да и в олимпиадах любят вопросы вроде какому дереву принадлежит тот или иной листик.
Деревья и их листья на фото и картинках
Хвойные деревья
Сосна
Сосна Лиственница Кедр Ель
Лиственные деревья
Береза
Береза
Береза — лиственное дерево, стройное, с густой кроной, широко растущей вверх. Белый ствол березы с темными горизонтальными полосами заметно выделяет ее среди других деревьев.
Белый ствол березы с темными горизонтальными полосами заметно выделяет ее среди других деревьев.
Ствол березы
Листочки березы насыщенно зелёный и напоминают по своей форме — ромб, закругленный с одного края и с зубчиками с другой.
Берёзовый лист
С приходом осени береза одна из первых меняет свой окрас на желтый.
Осенние березы
С приходом весны веточки дерева покрываются «сережками» — продолговатыми пушистыми скоплениями крохотных цветков березы,
которые созревая выбрасывают по ветку многочисленные семена.
Сережки березы
Осина
Осина
Дуб
Липа
Клен
Ольха
Ольха
Ива
Тополь
Тополь
Каштан
Каштан
Источник
Виды листьев у деревьев и растений
Листья являются важнейшими частями большинства растений. Благодаря им, происходит движение воды по растительной массе, превращение солнечного света в энергию роста и очищение окружающего воздуха.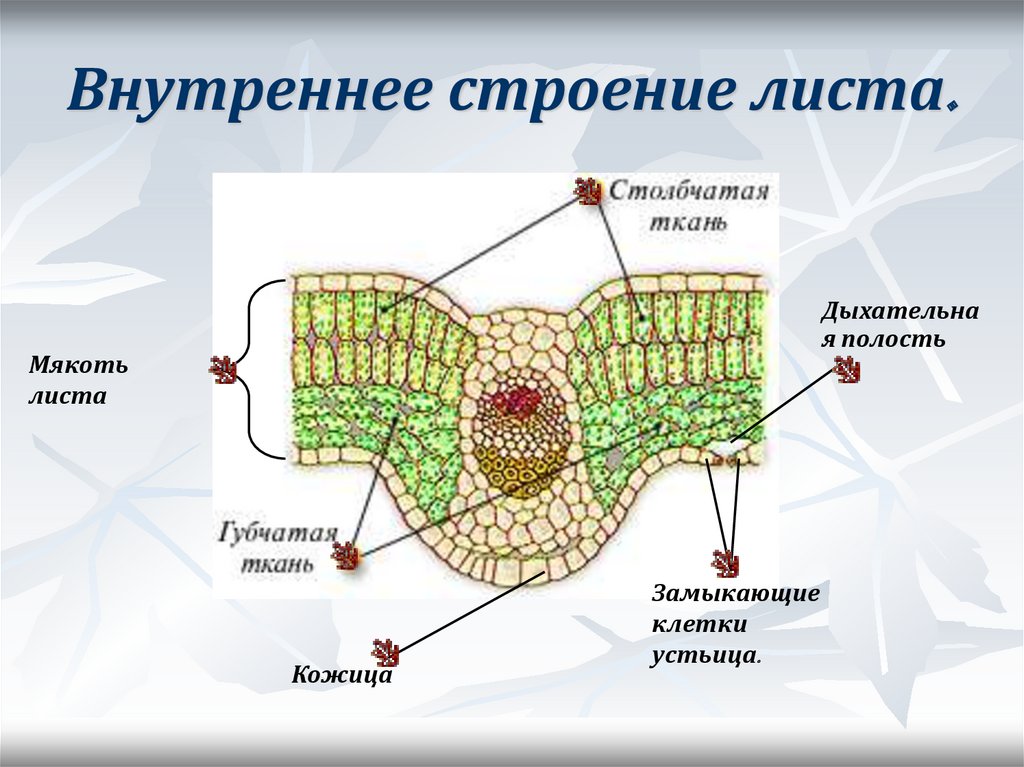 Существует множество биологических классификаций листьев на основе различных признаков. В рамках этой статьи рассмотрим основные из них.
Существует множество биологических классификаций листьев на основе различных признаков. В рамках этой статьи рассмотрим основные из них.
Что такое лист?
Лист – это наружная часть растения, которая отвечает за фотосинтез, испарение воды и газообмен между растением и окружающей средой. Они есть у подавляющего большинства растений, от едва заметной травы до огромных деревьев. При слове «лист» воображение сразу рисует классический листочек, наподобие березового. Однако существует огромное количество вариаций форм и конструкций, любая из которых служит для одних и тех же целей.
Основные виды листьев
Самая простая классификация листьев растений осуществляется по их форме. Согласно ей, существуют листовидные отростки (например, у папоротников), листья цветковых растений (классическая форма с черешком и листовой пластинкой), хвоя и обверточные листья (распространены у трав).
Виды, определяющиеся по расположению на стебле
Очередное или последовательное расположение означает, что листья начинают расти на стебле по одному на каждый узел.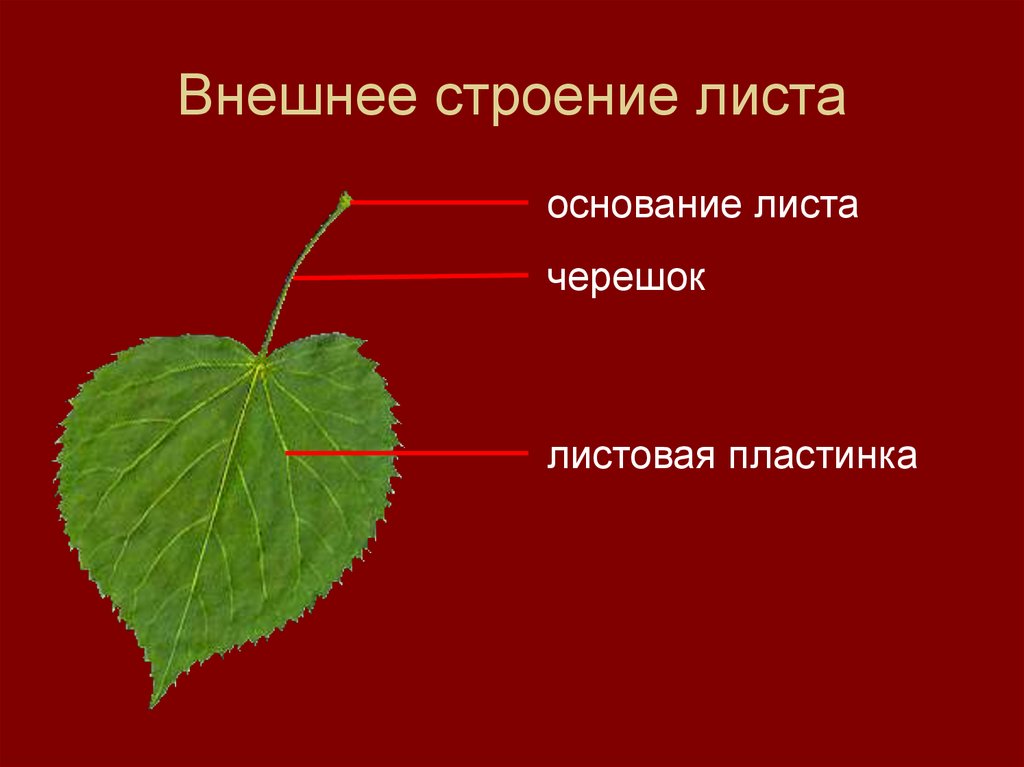 Термином «узел» обозначается место на стебле, которое используется под образование нового листа.
Термином «узел» обозначается место на стебле, которое используется под образование нового листа.
Супротивное расположение означает, что на каждом узле ветки или стебля вырастает два листка. Причем во многих случаях каждый последующий узел повернуть на 90 градусов относительно предыдущего.
Розеточное размещение листьев подразумевает их расположение на одной высоте и ориентацию по кругу. Грубо говоря, все листки такого растения растут из одной точки (корня) и образуют красивый раскидистый куст.
Также существует мутовчатое расположение. Оно похоже на супротивное, но имеет по три листа на узле. При этом узлы называются мутовками и могут быть так же последовательно развернуты на 90 градусов.
Классификация по типу листовых пластинок
В основе данной классификации лежит количество и разделение листочков, растущих на одном черенке, либо из одного узла стебля (ствола). В соответствии с этим, самый простой тип – простой лист. Он характеризуется наличием только одной листовой пластинки и одного черешка. Пластинкой называется сама поверхность листа, то есть, его «полотно» с прожилками. У простого листа оно может иметь любую форму, но вырезы никогда не доходят до черешка. Листья простого типа всегда опадают вместе с черешком, не оставляя на дереве ни одной своей части.
Пластинкой называется сама поверхность листа, то есть, его «полотно» с прожилками. У простого листа оно может иметь любую форму, но вырезы никогда не доходят до черешка. Листья простого типа всегда опадают вместе с черешком, не оставляя на дереве ни одной своей части.
Следующий тип – сложный лист. Здесь на одном черешке крепится сразу несколько листьев. При этом каждый из них может иметь свой собственный дополнительный черешок.
Виды листьев по их форме
Классификация по форме листа очень обширна. Ведь существует огромное количество растений с самой разнообразной листвой. В данный список входит более 30 названий, каждое из которых описывает определенную форму. Перечислять их все мы не будем, скажем только о наиболее распространенных.
Пожалуй, самой знакомым типом по данной классификации, является щитовидный. Листья такой формы имеет, например, береза. Они похожи на маленький щит и при этом имеют классические очертания листа. Есть и необычные, наподобие «обратносердцевидного». Этот тип имеет форму вытянутого сердечка, нижним, острым концом, примыкающим к черешку.
Этот тип имеет форму вытянутого сердечка, нижним, острым концом, примыкающим к черешку.
Также интересными являются мутовчатые листья. Этот тип, как правило, встречается у различных полевых трав и многих цветов. Вид «перисторассеченный» знаком каждому с детства – листья такой формы имеет одуванчик.
Нестандартная эволюция листьев
В ходе эволюции листва деревьев и растений претерпевала различные изменения. У большинства представителей флоры они не привели к серьезным изменениям, однако листья определенных растений начали выполнять специфические функции.
Ловчие листья
Пожалуй, самыми «узкоспециализированными» являются ловчие листья. Они присутствуют на хищных растениях, питающихся насекомыми. Яркий пример – росянка или венерина мухоловка. Главная задача такого листа – поймать насекомое, обеспечить его удержание, и переварить при помощи специальных ферментов. Способ поимки различен: в одних случаях лист вырабатывает липкий сок (росянка), в других резко закрывается (венерина мухоловка), в третьих в действие вступают особые пузырьки с клапанами (пузырчатка).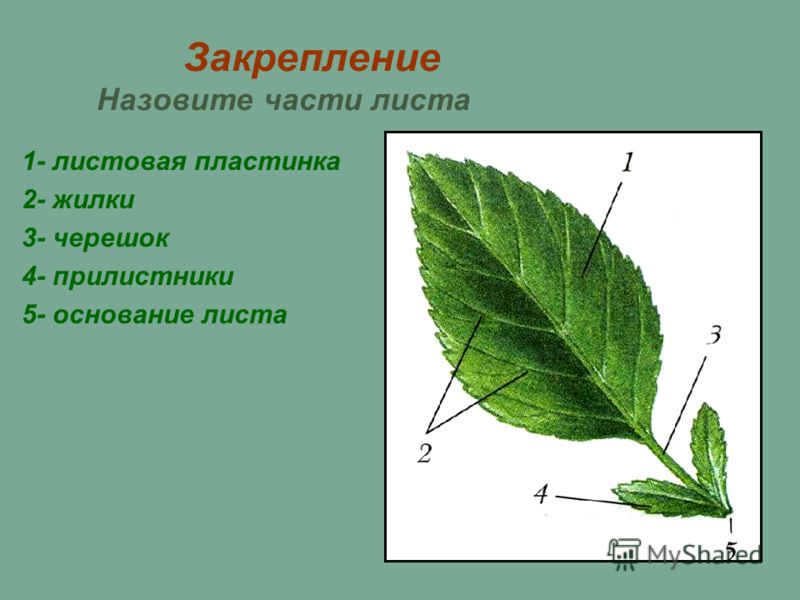
Суккулентные листья
Такой тип листьев предназначен для создания запасов воды. Самое знакомое нам растение, обладающее ими, – алоэ. Толстые и мясистые, они содержат внутри большое количество влаги, так как подобные цветы произрастают в засушливых регионах с малым количество осадков.
Мешковидные листья
Данный тип также хранит воду, но делает это не за счет толстого слоя мякоти, а при помощи воронки. Воронка же образуется самим листом, который закручивается особым образом и удерживает накопившуюся дождевую воду.
Колючки
В целях защиты, листья некоторых растений эволюционировали в колючки. Они могут быть видоизмененной листовой пластинкой, отвердевшей и заостренной, а могут образовываться из побегов.
Листья-усы имеются на стелющихся растениях, которым нужна опора. Представляют собой продолжение верхних частей обычных листьев в виде длинных, вьющихся отростков. Они цепляются за окружающие предметы, благодаря чему растение обвивает их. Такой тип листьев имеется у обычного садового гороха, огурца, тыквы.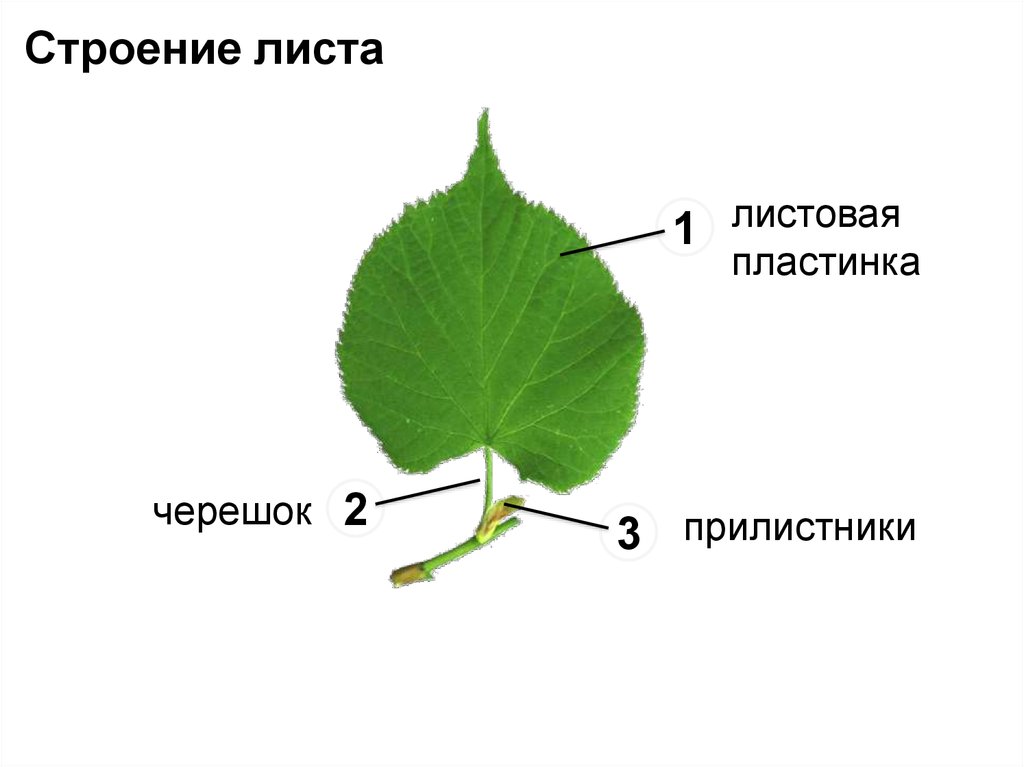
Филлодии
Филлодии являются частным случаем эволюции черешка. Такой черешок похож по форме на лист и способен выполнять фотосинтез. При этом настоящий лист, расположенный дальше, имеет упрощенное строение и деградирует.
Прицветники
Данный тип листьев характеризуется их полукруговой или круговой формой, зачастую с образованием небольшой воронки. В образованном углублении, как правило, располагаются листья другого типа или соцветия.
Источник
Листья растений
Люди пишут о них стихи и песни, любуются ими весной, летом и осенью, с нетерпением ждут их появления зимой. Они – символ жизни и возрождения природы, нежное одеяние, радующее глаз и дарящее чистый кислород всему живому на земле. Это листья – то, что мы видим каждый день и то, без чего не сможет прожить ни одно растение, да и вся наша планета.
— Листья желтые над городом кружатся, с тихим шорохом нам под ноги ложатся…
— Кленовый лист, кленовый лист, ты мне среди зимы приснись…
— Листьев зеленый звон всем тем, кто был влюблен…
Что такое листья, зачем они нужны, почему желтеют осенью и вновь вырастают зимой, какого цвета и формы бывают – всё это и многое другое вы узнаете из этой публикации.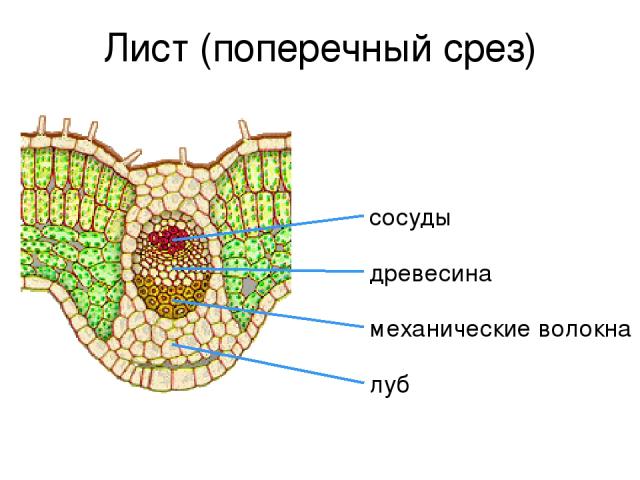
Функции листьев, их роль в жизнедеятельности растений
Если говорить сухим научным языком, лист – один из важнейших органов растения, главная функция которого – участие в процессе фотосинтеза.
[!] Фотосинтез – преобразование солнечной энергии в органические соединения внутри растения. Проще говоря, благодаря фотосинтезу, растения получают пищу от солнечных лучей.
Кроме этого, с помощью листьев растение дышит и испаряет влагу (выделяет росу).
Как видите, без зеленых покровов жизнь растения была бы невозможна, но не только растения зависят от листьев. С помощью этих своеобразных легких растение нейтрализует углекислый газ и выделяет кислород, необходимый и людям, и животным, и насекомым, то есть всему живому на планете.
Части листа
В целом, лист состоит из нескольких частей:
- Основание – место крепления к стеблю;
- Прилистник – листовидные элементы у основания, в некоторых случаях опадают после того, как лист полностью раскроется;
- Черешок – продолжение главной жилки листовой пластины, соединяющий лист и стебель;
- Листовая пластина – широкая часть листа, выполняющая его основные функции.

Так как каждое растение индивидуально, а листья бывают самыми разными, каких-то частей может и не быть. Например, часто отсутствуют прилистники, иногда не бывает черешка (в этом случае листья называются сидячими или пронзенными). Кроме того все части могут быть самой различной формы, длины и строения.
Классификация и разделение основных частей помогает ученым-ботаникам правильно идентифицировать растение и определить, к какому семейству, роду и порядку оно относится.
Строение, виды и формы листовой пластины
Листовая пластина состоит из верхнего эпидермиса, покрытого кутикулой, палисадного слоя, губчатого слоя и нижнего эпидермиса, также покрытого кутикулой. Каждый из слоев выполняет определенную функцию:
- Кутикула и эпидермис защищают пластину от внешних воздействий, препятствуют излишнему испарению воды.
[!] За процесс задержки необходимой влаги внутри листа отвечают устьица – парные клетки, способные замыкаться и не давать влаге испаряться.
Устьица начинают свою работу в засуху, спасая растение от обезвоживания.
- Палисадный слой, также называемый столбчатой тканью, отвечает за процесс фотосинтеза. Здесь собраны и хлоропласты, клетки окрашивающие поверхность листа в зеленый цвет.
- Губчатая ткань – основа листовой пластины. Её функции – газообмен, поглощение углекислоты и выделение кислорода, и фотосинтез.
Вся пластина пронизана проводящими пучками, называемыми жилками, по которым осуществляется доставка органических веществ от корня к листу (вода и минералы) и наоборот (раствор сахара). Кроме того, жилки образуют твердый скелет, защищающий мягкую ткань от разрывов.
Формы пластины
В целом все формы листьев делятся на простые и сложные, а сложные на пальчатые, перистые, двуперистые, трехлистные, перистонадрезанные, которые, в свою очередь, разделяются еще на несколько типов. Всего ботаника насчитывает не менее тридцати пяти разновидностей форм.
Простые листья состоят из одной листовой пластины, при этом она может быть самой разной формы: круглой, овальной, ромбовидной, удлиненной и так далее. Очертание кончика пластины и места крепления черешка также различаются.
Очертание кончика пластины и места крепления черешка также различаются.
Сложными называются те листья, которые состоят из нескольких частей, как сочлененных на общем черешке (лопастные, рассеченные, раздельные), так и имеющих свой отдельный черешочек (пальчатые, перистые, трехлистные).
[!] Одним из признаков сложных листьев является их опадание в разное время.
Кроме общей конфигурации листа различаются его основание (круглое, сердцевидное, треловидное, неравнобокое и т.д.), и верхушка (заостренная, выемчатая, усиковидная, притупленная и т.д.).
Формы края
Край листа, так же как и его общая форма, подсказывает ботаникам принадлежность растения к тому, или иному виду. В зависимости от глубины рассеченности, края делятся на пальчатые или зубчатые (неглубокие выемки), лопастные, рассеченные и раздельные (глубокие выемки). Гладкие края носят название цельнокрайние.
Виды жилкования
Рисунок жилкования листовой пластины может быть весьма разнообразным и зависит от вида растения.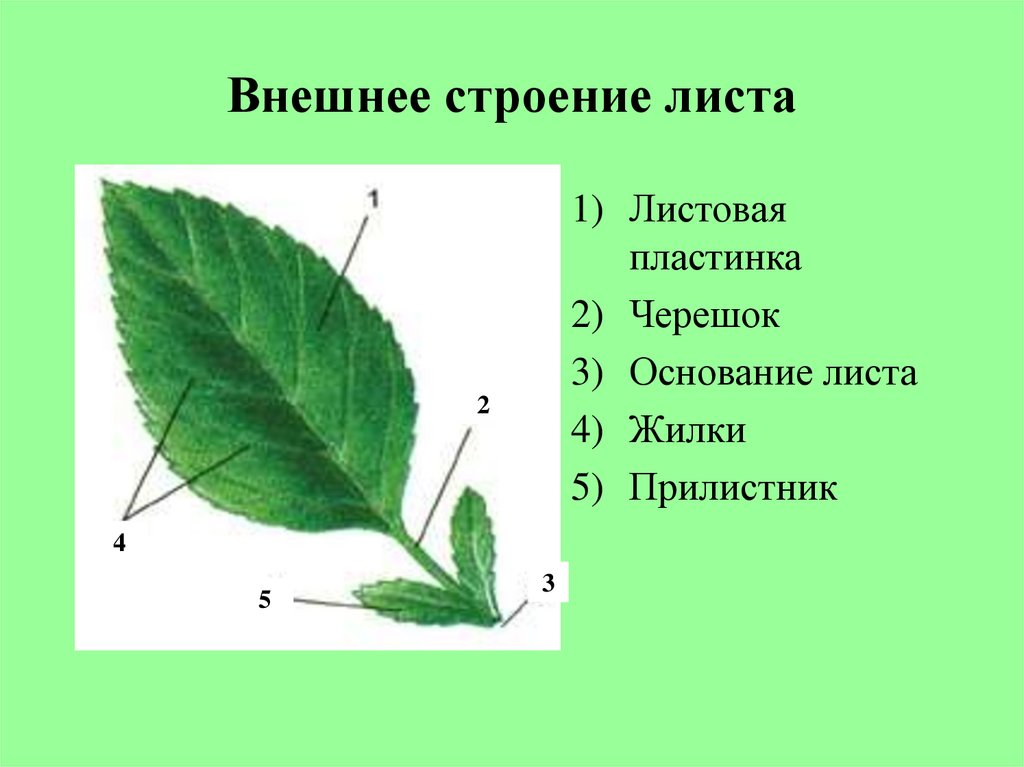 В целом все типы жилкования делятся на две части:
В целом все типы жилкования делятся на две части:
- сквозь листовую пластину проходит несколько параллельных жилок, центральная жилка при этом отсутствует (параллельное жилкование),
- есть главная (центральная) жилка, от которой ответвляются боковые (сетчатое жилкование),
- несколько выгнутых жилок, расходящихся в середине листа и сходящихся к краю (дуговидное жилкование).
В свою очередь сетчатое жилкование разделяется на несколько подвидов.
Виды прилистников и черешков
Прилистник обычно похож на маленький недоразвитый листик, располагающийся у основания листа. Они могут опадать после того, как лист полностью раскроется, или оставаться на растении. В зависимости от способа крепления к черешку, прилистники бывают свободные, сросшиеся с черешком, межчерешковые, раструбовидные или опоясывающие основание черешка.
Черешки могут различаться по форме среза: цилиндр, полуцилиндр, с выемкой и другие. Кроме того, как говорилось выше, черешка может не быть совсем, в этом случае лист прикрепляется непосредственно к стеблю.
Как видите, растительный мир демонстрирует удивительное многообразие форм, при этом существуют миллионы их сочетаний.
Итак, научно-ботаническая часть окончена, самое время перейти к удивительным фактам о листьях.
Это интересно
Как растения приспосабливаются к климату и другим условиям обитания с помощью листьев
Каждое растение вынуждено акклиматизироваться к погодным условиям, а также защищаться от внешних воздействий. Все части растения: корни, побеги, цветы и, конечно, листья, приспособились к различным климатическим явлениям: высокой или низкой температуре, засухе или излишней влажности, недостатку или избытку солнечных лучей. Кроме того, растениям угрожают люди и животные, поэтому многие из них в процессе эволюции научились отражать атаки.
Рассмотрим, как с помощью своего зеленого покрова растение противостоит неблагоприятной среде.
Засушливый или влажный климат:
- Небольшой размер листьев и, соответственно, маленькая площадь листовой пластины препятствует излишнему испарению воды;
- Листья обычно толстые, сочные – таким образом в них накапливается необходимая влага;
- Листовые пластины многих растений покрыты волосками, что также препятствует испарению;
- Той же цели служит гладкий восковой налет на поверхности.

- Крупные листья – признак растений тропического климата, за счет большого размера пластины процесс испарения происходит гораздо более интенсивно.
- Рассеченная, изрезанная форма края свободно пропускает воздушные потоки, благодаря этому порывы ветра не травмируют лист.
Места с избыточным или недостаточным солнечным светом:
- Если солнечного света недостаточно многие растения могут разворачивать листья таким образом, чтобы как можно больше солнечных лучей попало на их поверхность;
- Листовая мозаика – явление, при котором меньшие по размеру листья располагаются между более крупных собратий. В этом случае каждый лист улавливает солнечные лучи и участвует в процессе фотосинтеза;
- Некоторые растения, не нуждающиеся в большом количестве солнца, фильтруют свет через специальные полупрозрачные окошки, расположенные на листьях.
Водные растения – эти представители флоры стоят особняком, ведь им, чтобы выжить, пришлось приспособиться даже не к климату, а к совершенно другой стихии – воде:
- Листья гидатофитов (растения, полностью погруженные в воду) сильно расчленены.
 Таким образом, увеличивая площадь поверхности, растение получает необходимое количество кислорода;
Таким образом, увеличивая площадь поверхности, растение получает необходимое количество кислорода; - Листья, плавающие по поверхности водоема, не имеют устьиц с обратной стороны листовой пластины;
- Большая площадь поверхности плавающих листьев не дает им утонуть за счет распределения нагрузки.
- Специальные микроскопические выступы и восковой слой препятствует проникновению воды внутрь листа, исключая заражение растения микроорганизмами и простейшими водорослями. Вода не впитывается в поверхность, а стекает по листу каплями, заодно очищая его от пыли и грязи. Это явление получило название «эффект лотоса».
Защита от животных и людей. Некоторые растения в ходе эволюции научились обороняться от посягательств:
- Листья вырабатывают сильно пахнущие феромоны и масла, отпугивающие животных;
- Листовая пластина бывает покрыта мягкими волосками или даже твердыми колючками, жалящими агрессора.
Необычные листья
Природа наделила отдельные виды растений столь экстравагантной внешностью, что иногда определить, где перед нами листья – представляется непростой задачей.
Кактусообразные обосновались на территориях с засушливым климатом, где потеря каждой капли воды равносильна гибели. Эволюционный отбор сделал свое дело — выжили экземпляры с минимальной площадью испарения. Широкие листья – непозволительная роскошь для таких условий существования. Все внешнее убранство кактусов, жителей безводных пустошей — компактные защитные листья-колючки.
Лесные кактусы пошли еще дальше – их колючки стали тонкими, почти невидимыми волосками, а шарообразный ствол превратился в цепочку плоских сегментов.
Опунция, трихоцериус, шлюмбергера
Другие растения засушливых районов, чтобы не испарять драгоценную влагу, и вовсе решили отказаться от листьев. Вернее, листья у них остались, но только в виде маленьких неразвитых чешуек. При этом форму листа и функцию фотосинтеза приобрели побеги, называемые кладодиями или филлокадиями. Филлокадии настолько приспособились к новой роли, что внешне практически не отличаются от обычного листа, однако фактически таковыми не являются.
Есть и вариант наоборот – то, что кажется побегами, на самом деле является листьями. Один из примеров – усики ползающих растений. В этом случае усики – это верхние части листьев, приноровившиеся цепляться за опору.
Иглица, аспарагус, горошек заборный
Одни их самых необычных листьев принадлежат тропическим экзотам. Жаркий влажный климат, обилие насекомых и животных вынудило растения подстроиться под непростые условия существования и даже перейти в разряд хищников. С помощью липкого секрета или специальных пузырьков на листьях, хищные растения ловят зазевавшихся насекомых, а затем высасывают из них жизненные соки.
Еще одно приспособление тропических растений – мешок, образованный сросшимися плоскостями листовой пластины. В эту ловушку собирается дождевая вода, запас которой, по необходимости, расходуется в периоды засухи.
Росянка, пузырчатка, дисхидия Раффлеза
Листья разных цветов
Какого цвета бывают листья? На первый взгляд ответ на этот вопрос очень прост – зелеными летом, желтыми и красными осенью. На самом деле, они могут быть самых разных цветов не только осенью, но и в другое время года. Можно встретить зеленые, желтые, красные, серебристые бордовые и, даже, фиолетовые оттенки окраски естественного убранства вполне здоровых растений. Кроме необычной пигментации, на листьях некоторых, особенно южных, растений есть красивейшие узоры и орнаменты.
На самом деле, они могут быть самых разных цветов не только осенью, но и в другое время года. Можно встретить зеленые, желтые, красные, серебристые бордовые и, даже, фиолетовые оттенки окраски естественного убранства вполне здоровых растений. Кроме необычной пигментации, на листьях некоторых, особенно южных, растений есть красивейшие узоры и орнаменты.
Автор всего этого великолепия не только природа, но и человек: в процессе селекции было выведено много декоративных сортов, красующихся разноцветными или узорчатыми листьями.
Зебрина, фиттония, каладиум
Съедобные листья
Листья не только радуют глаз и необходимы для жизни планеты, некоторые из листьев еще и съедобны и, более того, составляют немалую часть рациона человека. В кулинарии их используют и в качестве овощной составляющей: шпинат, мангольд, пекинская, китайская капусты, и в качестве салатных ингредиентов: рукола, щавель, латук, и, конечно, в качестве приправ: укроп, петрушка, базилик, мята и так далее.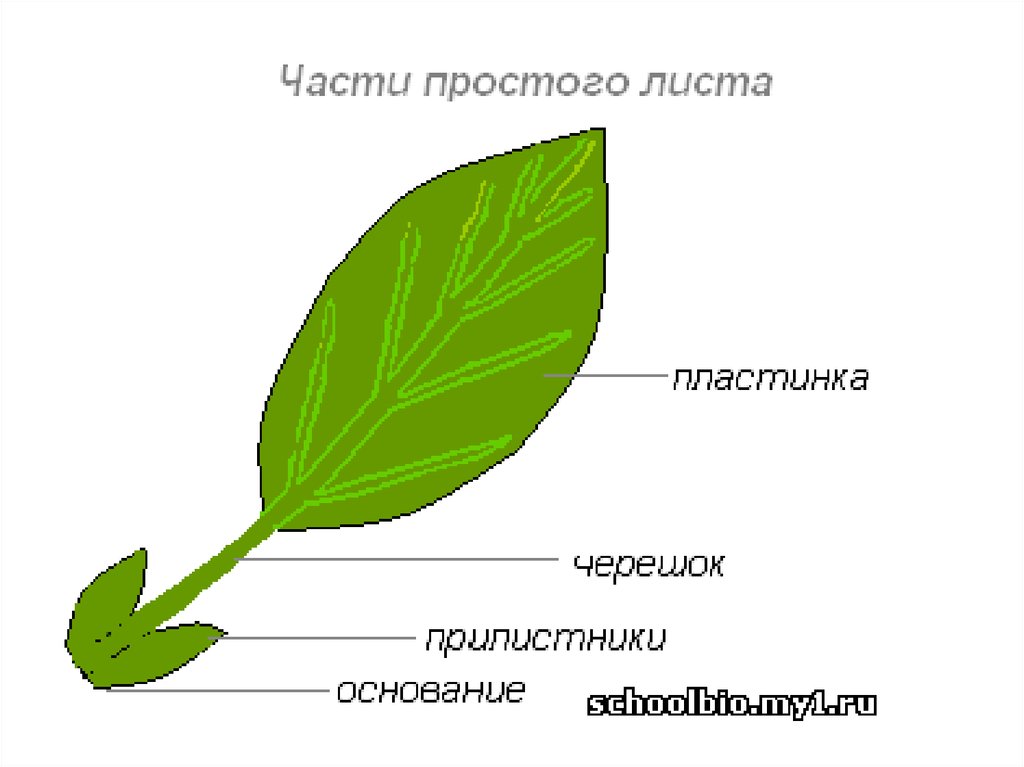
Пекинская капуста, салат-латук, базилик
Ответы на вопросы
В завершении статьи – ответы на самые популярные вопросы о листьях.
Почему лист плоский?
Такая форма увеличивает площадь листовой пластины, а, в свою очередь, большая площадь поверхности увеличивает количество клеток, участвующих в процессе фотосинтеза.
От чего зависит размер листа?
Размер и, соответственно, площадь поверхности листа, зависит от места обитания растения. Листья растений из засушливых районов обычно мелкие, а из влажных мест – крупные. Дело в том, что чем больше площадь листа, тем больше на его поверхности устьиц и тем более интенсивно происходит испарение воды. Там, где часто бывает засуха, чтобы выжить растения стараются не испарять много влаги, а в тропическом климате процесс испарения, наоборот, должен быть как можно более интенсивным.
Почему листья зеленые?
За зеленый цвет листа отвечает хлорофилл, участвующий в преобразовании углекислого газа в питательные вещества. Высокое содержание хлорофилла в листовой пластине придает одеянию растений свежий зеленый оттенок.
Высокое содержание хлорофилла в листовой пластине придает одеянию растений свежий зеленый оттенок.
[!] Хлорофилл некоторых растений окрашен в другие цвета – красный, бурый, фиолетовый, поэтому листья таких растения имеют соответствующие оттенки.
Почему листья желтеют?
Осенью хлорофилл в листьях разрушается, его становится меньше. За счет уменьшения хлорофилла постепенно уменьшается и интенсивность зеленого спектра. На первый план выступают желтые и красные пигменты (ксантофилл, каротин, антоциан), содержащиеся в клетках листа.
[!] Листья отдельных растений не меняют окраску и опадают зелеными.
Почему листья опадают осенью?
Сезонные изменения длительности светового дня и среднесуточных температур, вынудили растения приспосабливаться к переменчивым условиям существования. К наступлению зимних холодов большая часть флоры сбрасывает летнее убранство и переходит в состояние анабиоза, принятое называть зимней спячкой. Обменные процессы в жизненных системах растений практически останавливаются. Листья, столь необходимые летом для испарения излишков влаги и сбора живительного солнечного света, становятся попросту ненужными и опадают.
Обменные процессы в жизненных системах растений практически останавливаются. Листья, столь необходимые летом для испарения излишков влаги и сбора живительного солнечного света, становятся попросту ненужными и опадают.
В течение весны и лета листья добывают и перерабатывают питательные вещества необходимые для жизнедеятельности растений. В процессе такой переработки, зеленые легкие природы вырабатывают и накапливают метаболиты – лишние минеральные соли, таким образом, исполняя роль своеобразного фильтра. Со временем, отложений становится все больше и осенью растение избавляется от листа, который перестает приносить пользу.
Так уж заведено в природе, ни что не пропадает зря. Опавшие листья укрывают землю от мороза, защищая почву. В теплое время года, ковер устилающий почву постепенно разлагается, перепревает. Насекомые, бактерии и микроорганизмы перерабатывают образовавшийся перегной в питательную почву для живых растений, замыкая круговорот в природе.
[!] Именно поэтому не рекомендуется убирать опавшие листья осенью.
Будет лучше оставить их до пробуждения природы, а весной, после того, как они исполнят свою роль, собрать.
Почему листья сохнут, чернеют, откуда на листьях появляются пятна?
Листья – своеобразный индикатор состояния растения. Симптомы различных заболеваний, признаки насекомых-вредителей, как правило, в первую очередь появляются на листьях. Если листья желтеют, краснеют, на них появляются различные пятна, вздутия, сухие участки – растение болеет и нуждается в незамедлительной помощи.
Если о человеке можно судить по одежке, то об окружающей нас флоре, можно судить именно по листьям. Как правило, именно зеленое убранство является для нас символом всех растений, а главная задача человека – сберечь и сохранить природу.
Источник
Строение дерева. От клеток до корней
Строение растений мы изучали еще в школе. В этой статьей мы решили напомнить, что из себя представляет дерево, и рассказать о каждой из его частей: клетках и тканях, древесине и коре, ветвях и ветках, листьях и корнях.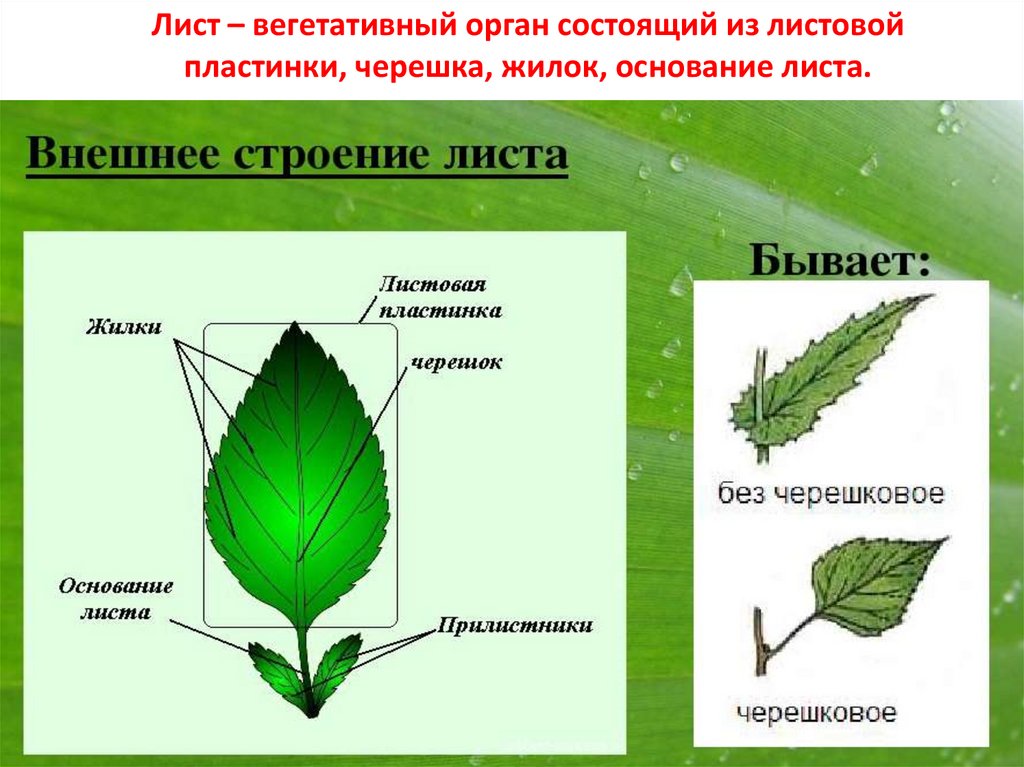
Материал был взят из первого русскоязычного издания справочника Европейского специалиста по уходу за деревьями (European Tree Worker), который пригодится как владельцам питомников и садовых участков, так и сертифицированным специалистам.
Деревья – это древесные растения большого размера. Они обладают уникальными свойствами, позволяющими им являться доминирующим видом царства растений во многих странах мира. В основе ухода за деревьями (древоводства) лежит глубокое понимание процессов роста и развития деревьев. Только с учетом данного принципа можно профессионально осуществлять уход за деревьями.
- Клетки и ткани
Для всех живых организмов характерна общая организационная структура, состоящая из клеток, тканей и органов. Клетки – это основные «строительные блоки» данной структуры. У растений новые клетки образуются путем деления существующих. Этот процесс проходит в специальных образовательных тканях – меристемах.
1 — Молодая клетка с плазмой и ядром 2 — Рост клетки 3 — Зрелая клетка с большой вакуолью
После деления клетки проходят этап дифференцировки, во время которого изменяется их структура и они приобретают способность к различным специфическим функциями. Клетки с аналогичной структурой и функциями объединяются в ткани.
Затем из тканей формируются органы, которых у растений шесть: листья, стволы, корни, почки, цветы и плоды
. И, наконец, органы образуют полностью функциональные организмы – деревья.Существует два основных типа меристематической ткани:
- первичная меристема, из которой образуются клетки, отвечающие за рост побегов и корней в длину;
- вторичная меристема, из которой образуются клетки, отвечающие за прирост в диаметре.
У деревьев есть две вторичные меристемы: камбий и феллоген.
- Камбий выполняет крайне важную роль: в процессе деления в нем образуются новые клетки, формирующие систему сосудов дерева. Из него генерируются два вида ткани: ксилема во внутренней части и флоэма снаружи.
- Феллоген
Когда дерево срубают и рассматривают в поперечном сечении, в ксилеме видны годичные кольца. В зонах умеренного климата данные кольца соответствуют годовому образованию ксилемы в камбии. Они имеют форму круга, так как относительный размер и плотность сосудистой ткани изменяются в течение вегетационного периода. По мере приближения к концу вегетационного периода клетки становятся меньше в диаметре.
По мере приближения к концу вегетационного периода клетки становятся меньше в диаметре.
Таким образом, благодаря резкой разнице между клетками, образованными в начале сезона (ранняя древесина), и клеткам, сформированными позднее (поздняя древесина), индивидуальный годовой прирост становится различимым.
В отношении древесины хвойные и лиственные породы значительно отличаются друг от друга. Кроме того, среди лиственных деревьев выделяются кольцесосудистые (например, Дуб (Quercus), Ясень (Fraxinus)) и рассеяннососудистые виды (например, Липа (Tilia), Бук (Fagus)).
В центре ствола формируется ядровая древесина. Она окружена живой заболонью. Не все проводящие элементы ксилемы служат для передвижения воды. За это отвечает только живая и активная ткань заболони, тогда как другая часть ксилемы, расположенная ближе к центру, является нефункциональной.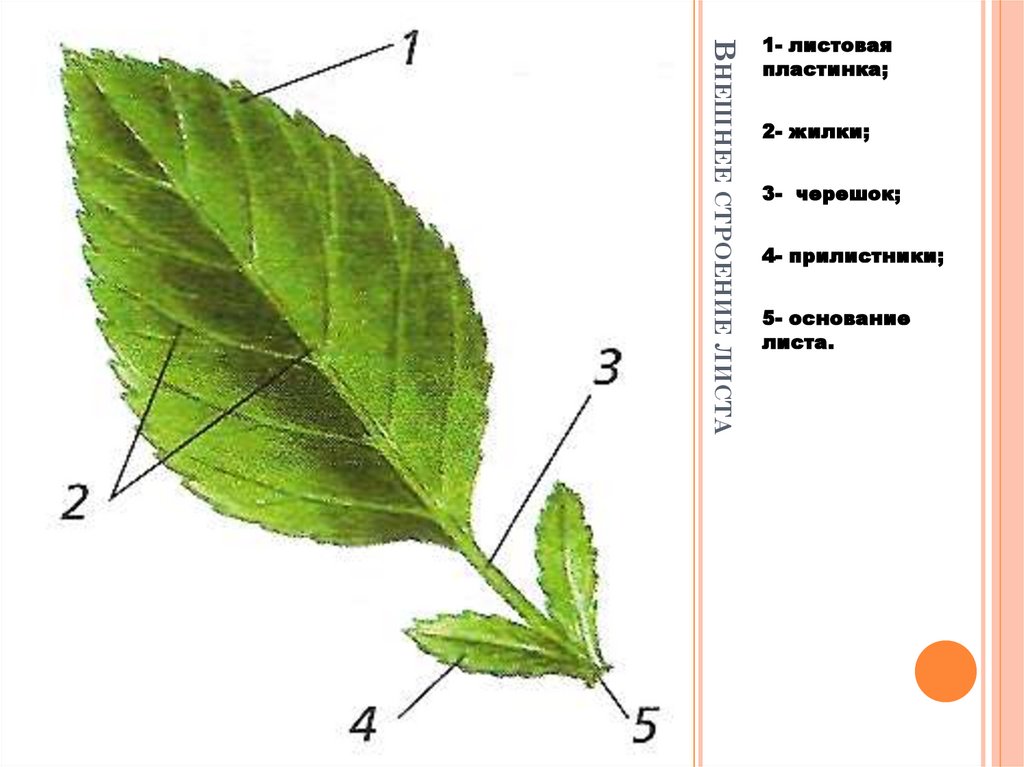 Такие мертвые клетки образуют ядро –
Такие мертвые клетки образуют ядро –
Флоэма отвечает за перемещение сахара от листьев к другим частям растения. Кроме флоэмы и ксилемы, сосудистая система дерева включает в себя лучевые клетки. Лучи расходятся в радиальном направлении от центра поперечного сечения через флоэму и ксилему и служат для транспортировки сахаров и их компонентов вдоль ствола. Они помогают ограничивать распространение гнили по древесной ткани и хранить запасы питательных веществ в виде крахмала.
Поперечный разрез стволаВнешняя часть ветвей и ствола деревьев называется корой. Это защитная ткань, поддерживающая температуру внутренней части ствола, предохраняющая растения от повреждений и уменьшающая потерю воды. Кора состоит из нефункциональной флоэмы, пробковой ткани и мертвых клеток. Для минимизации потери воды ее клетки пропитаны воском и маслами.
Газообмен между живыми тканями дерева и атмосферой происходит с помощью чечевичек, маленьких пор в коре.
Кора различных деревьев имеет разное строение и свойства. Например, кора бука очень гладкая с небольшим количеством пробковой ткани, а кора дуба, наоборот, образует толстые слои феллемы.
- Ветви и ветки
Ветки – это небольшие ветви, которые служат опорой для листьев, цветов и плодов. Ветви поддерживают ветки, а ствол поддерживает всю крону. Ветви и ветки развиваются из двух типов почек:
- терминальных или верхушечных почек на конце побега;
- боковых или пазушных почек, которые образуются вдоль ветки.
Верхушечная почка является наиболее сильной на ветви или ветке и располагается на конце побега. Она контролирует развитие вторичных почек с помощью гормонов. Обычно вторичные почки не развиваются и остаются в спящем состоянии. Как правило, верхушечная почка является наиболее активной на каждой ветви или ветке и контролирует развитие 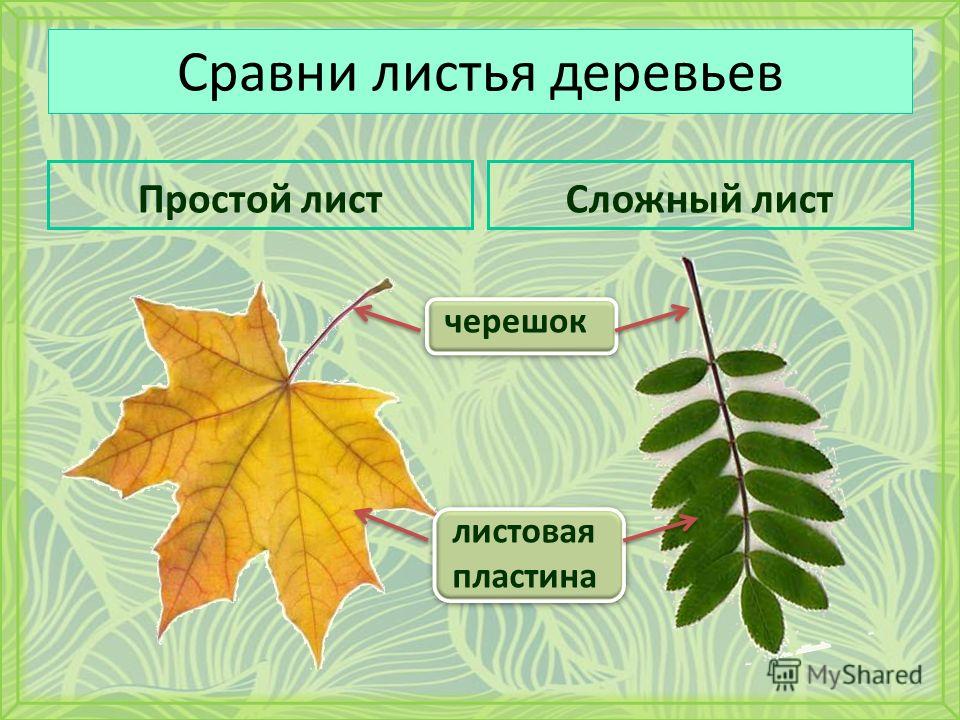
Побеги с доминирующей верхушечной почкой бывают моноподиальными или симподиальными.
Побеги без апикального доминирования являются ложнодихотомическими.
Гибель верхушечной почки в результате случайного повреждения или обрезки может привести к активизации спящих почек рядом со срезом и, как следствие, к развитию нового побега.
Некоторые побеги развивают придаточные почки, которые формируются вдоль стволов и корней. Они возникают, как правило, в ответ на потерю обычных почек в результате действия регуляторов роста.
Ежегодный прирост: 1 — 1 год; 2 — 2 года; 3 — 3 годаЛистья и почки образуются из немного утолщенной части ветки, которая называется узел. Междоузлие – это зона между узлами. На ветке видны листовые рубцы и рубцы верхушечной почки. Они помогают измерять ежегодное удлинение ветки и общий прирост. По своей структуре и функции каждая ветвь дерева сопоставима со всей кроной.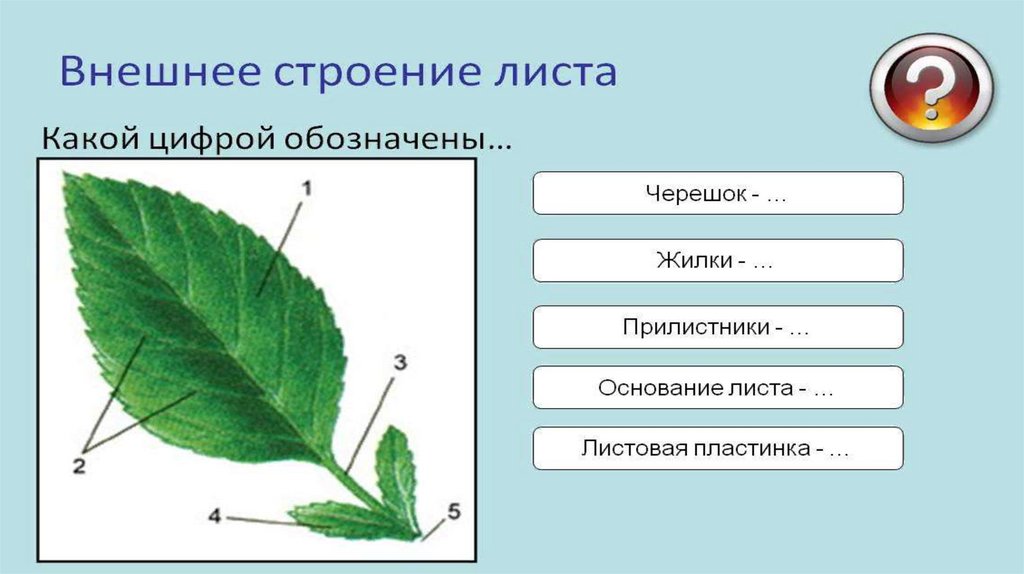 Но в то же время ветви – это не просто отростки ствола.
Но в то же время ветви – это не просто отростки ствола.
Наоборот, ветви характеризуются уникальной формой присоединения к нему, которая имеет крайне важное значение для практической деятельности в сфере ухода за деревьями, например, для обрезки.
Ветви прочно крепятся к древесине и коре, расположенной под ветвями, но над ними крепление более хрупкое. Годовой прирост слоев ткани в зоне соединения ветви и ствола хорошо заметен и формируется большую часть времени. Плечо или выпуклость вокруг основания ветви называется
- Листья
Листья отвечают за производство питательных веществ для дерева. Они содержат хлоропласт, наполненный зеленым пигментом – хлорофиллом, с помощью которого происходит фотосинтез. Еще одна функция листьев – транспирация, представляющая собой выведение воды через листву посредством испарения.
Они содержат хлоропласт, наполненный зеленым пигментом – хлорофиллом, с помощью которого происходит фотосинтез. Еще одна функция листьев – транспирация, представляющая собой выведение воды через листву посредством испарения.
5 — Клетки губчатой паренхимы
Площадь листьев достаточно большая, что позволяет им поглощать солнечный свет и углекислый газ, необходимые для фотосинтеза.
Внешняя поверхность листа покрыта воскообразным слоем, который называется кутикула. Она служит для минимизации дессикации (высушивания) листа.
Испарение воды и газообмен контролируют устьица – маленькие отверстия на поверхности листа.
Лист обладает развитой системой проводящих тканей, включающей в себя вены, или капиллярные каналы. Вены состоят из тканей как флоэмы, так и ксилемы, и отвечают за транспортировку воды и жизненно необходимых веществ, а также за перенос питательных веществ, которые вырабатываются в клетках листьев, к остальным органам дерева.
Деревья, сбрасывающие листву каждый год, называются лиственными, а те, которые сохраняют ее в течение более чем одного года, называются хвойными или вечнозелеными. Осыпание листьев обусловлено клеточными изменениями и регуляторами роста, формирующими точку отделения органа у основания черешка, или ножки листа.
Точка отделения листьев выполняет две функции:
- обеспечивает осыпание листвы осенью;
- предотвращает высыхание, распространение болезней и повреждение части растения, от которой отрывается лист.
Осенью изменение цвета листвы листопадных деревьев связано с разложением хлорофилла, позволяющим проявиться другим пигментам, содержащимся в листьях. Сокращение продолжительности светового дня в сочетании с холодными ночами приводит к усиленному накоплению сахаров и замедляет выработку хлорофилла. Этот процесс и позволяет другим пигментам, в том числе антоцианинам (красный и пурпурный) и каротиноидам (желтый, оранжевый и красный), проявиться.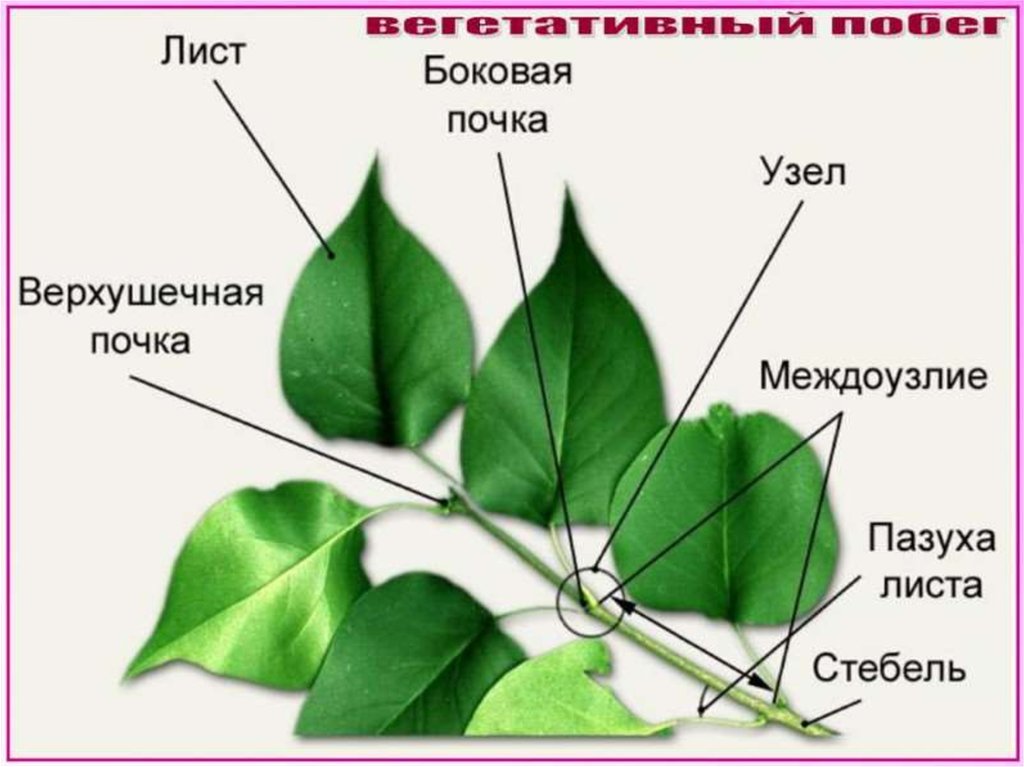
- Корни
Корни деревьев выполняют четыре основные функции:
- фиксация дерева;
- аккумуляция энергии и питательных веществ;
- поглощение веществ;
- транспортировка веществ.
1. Одревесневший корень
2. Корневой волосок
3. Корневой кончик
4. Корневой чехлик
Всасывающие корни представляют собой небольшие, волокнистые участки ткани, растущей на окончаниях основных одревесневших корней. У них есть эпидермальные клетки, модифицированные в корневые волоски, которые помогают поглощать воду и минеральные вещества. Корневые волоски живут совсем не долго (3–4 недели весной) и значительно активизируют способность к поглощению веществ с наступлением вегетационного периода весной.
Что касается корневых кончиков, они содержат меристему, где клетки делятся и растут в длину.
Корни растут там, где они могут найти воздух и кислород. Большая часть всасывающих корней находится на расстоянии 30 см от поверхности почвы. Также рядом с поверхностью располагаются горизонтальные боковые корни.
Большая часть всасывающих корней находится на расстоянии 30 см от поверхности почвы. Также рядом с поверхностью располагаются горизонтальные боковые корни.
Якорные корни растут вертикально по направлению вниз от боковых корней, обеспечивая надежную фиксацию дерева и увеличивая глубину освоения почвы корневой системой.
Корневая система:1 — Стержневая корневая система 2 — Мочковатая корневая система 3 — Поверхностная корневая система
Корни многих растений находятся в симбиозе с некоторыми грибами. Результат таких взаимоотношений называется микориза (грибокорень). Симбиоз двух организмов (дерева и грибов в нашем случае) основывается на взаимной пользе: грибы получают питательные вещества из корней и, в свою очередь, помогают корням всасывать воду и жизненно необходимые элементы.
Сожительство микоризы и растения, как правило, бывает чрезвычайно взаимовыгодно, что обусловлено объединением имеющихся у них различных способностей.
_____________________________________________________________________
Появление первого русскоязычного издания справочника Европейского специалиста по уходу за деревьями (European Tree Worker) в России стало возможным благодаря сотрудничеству НПСА «ЗДОРОВЫЙ ЛЕС» (Россия) с ведущим немецким учебным заведением в области подготовки специалистов по уходу за деревьями – Нюрнбергской школы ухода за деревьями (Германия).
Лист внешнее и внутреннее строение, жилкование, формы
Лист – внешнее и внутреннее строение, жилкование
Лист, внешнее и внутреннее строение, жилкование – без этих знаний нельзя понять, как живут зелёные растения.
Питание, дыхание, испарение и даже размножение – эти функции выполняют боковые части побегов, листья.
В процессе жизнедеятельности растения они выделяют кислород, необходимый живым организмам.
Поражает разнообразие двустороннесимметричных листовых пластинок, их размеров, видоизменения и расположение.
Главные функции листа называются фотосинтезом, газообменом и транспирацией. К дополнительным функциям листьев относятся:
- размножение, когда лист даёт корешки; запас питательных веществ луковыми, капустными листьями, алоэ;
- защита, когда листья видоизменились в колючки, поддержание, если это усики;
- листья насекомоядным растениям нужны для ловли еды;
- сбрасывание листьев, в которых накапливаются отходы жизнедеятельности.

К главным частям листа относят листовую пластинку с черешком. У сидячих экземпляров утолщённые основания пластинок называют листовыми подушками. На некоторых листьях встречаются прилистники из двойных листовидных образований внизу пластинки.
Виды прилистников: листки, плёнки, колючки или чешуйки. Это опадающие или сохраняющиеся части. Прилистники нужны для защиты пазушных боковых почек, вставочной и образовательной тканей листа.
К листовым пластинкам относятся тонкие расширенные части, где различают основу, кончик и край. По пластинкам идут центральные и ветвящиеся боковые жилки из сосудисто-волокнистых пучков. Размеры пластинок: маленьких – 1,5 мм (у Вольфии шаровидной), гигантских – 20 м (листья пальм).
Живут листья 3-8 месяцев, а некоторые – 15 с лишним лет.
Основание пластинки крепится к короткому или длинному черешку – суженной части, которая соединяет лист и стебель. Это черешковые листья.
Черешок:
поворачивает лист пластинкой к солнцу;
ослабляет удары дождевых капель или града;
проводит органические вещества в стебель, а обратно воду с минералами.
Безчерешковые листья – сидячие экземпляры, встречаются у злаков и камыша. С обхватыванием основанием листовой пластинки трубки стебля – это влагалищный вариант листьев, который выполняет защитную функцию (злаки, осока, зонтичные растения).
Внешний вид листьев характеризуют очертанием, степенью расчленённости, видом верхушки и основания. У простых листьев:
овальная форма у акации;
округлая;
ланцетная;
продолговатая;
яйцевидная у груши;
линейная;
копьевидная;
сердцевидная у липы;
обратно яйцевая и другие подобные формы.
У одуванчика листья струговидные – с треугольными сегментами, редька с лировидными листьями – у основания пластинки доли маленькие, а на верхушке – увеличенные.
Если пластинка удлинённая, то кончик острый или тупой, заострённый или остроконечный, притуплённый или выемчатый, усикообразный или усечённый.
Различаются листья и краем пластинки:
- распространённым видом считается цельный край такой, как у листьев сирени;
- много растений, у листьев которых зубчатые края – листья орешника, бука, яблони;
- у листьев барбариса колючезубчатыекрая;
- края листьев осины – пильчатые;
- с городчато-пильчатыми краями (стороны зубцов различающаяся) – листья груши;
- листья шалфея – с бородчатым краем: с острыми выемками и тупыми выступами.
Встречаются листовые пластинки, как бы разрезанные к центру. Если «разрезы» составляют:
одну треть расстояния с края до центра, речь идёт о лопастном листе, а части называют лопастями;
две трети, значит лист раздельный с частями-долями;
расстояние до центра или до нижнего края, это рассечённый лист, поделённый на сегменты.
Число «разрезов» так же влияет на форму и название: 3 – тройчатые, 5 – пальчатые, больше 5 – перистые листья.
По количеству пластинок на черешке, их относят к простым или к сложным листьям. Черешок простого листа, у берёзы, дуба, тополя – с одной листовой пластинкой и она целостная или расчленённая. Когда начинается листопад, такие листья опадают. У сложных листьев, на одном главном черешке, расположено 3 с лишним листовых пластинки и у каждой собственный черешок. Падают листочки по отдельности или листом.
Листья с тремя листочками называются тройчатыми (клевер, клубника). Если место соединения единое, а пластинка похожа на опахало – пальчато сложными листьями (люпин). Перистосложные листья с листочками, расположенными одинаково с обеих сторон черешка. Когда на конце остаётся листок без пары – непарноперистый лист (у шиповника, рябины, ясеня).
При парных листочках говорят о парноперистых листьях (у жёлтой акации). Когда на перистосложных листьях листочки так же сложные, это – дважды перистосложные листья.
Для образования жилок листа используются сосудисто-волокнистые пучки, соединяющие листовую пластинку со стеблем. Получившаяся сеть, ясно видимая с нижней стороны, называется жилкованием. У жилок 2 главные функции: опора, прочность и защита тканей от разрывов, что называется механической функцией. Транспорт веществ – воды с растворёнными минералами и органикой.
Различают 3 вида жилкования:
- Если видна главная жилка с боковыми ответвлениями, значит, речь идёт о сетчатом, перистом и пальчатом жилковании, которым наделены листья двудольных растений. Такой тип относят к открытому жилкованию.
- При дуговидном типе пучки идут дугами, начинаясь с черешка. В середине расстояние между жилками увеличивается, в конце листа жилки снова соединяются. Такой, закрытый, тип жилкования у ландыша, подорожника и других растений с расширенными и удлинёнными листьями.
-
Длинные узкие листья пшеницы, кукурузы – с параллельным жилкованием. К концу листа жилки сближаются. Дуговым и параллельным жилкованием отмечены однодольные растения.
Переходные виды жилкования:
- У листовых пластинок герани, клёна, люпина и других похожих листьев – пальчатое жилкование. Здесь основание листа даёт начало жилкам, которые идут как лучи или как веер.
- Если черешок крепится не к основанию пластинки, а к центральной части, то у такого листа звёздчатый тип жилкования (настурция).
- При дихотомическом, или вильчатом жилковании отсутствует главная жилка. Жилки делятся на 2 группы и идут к краю пластинки (гинкго и некоторые папоротники).
У листьев такое расположение на стеблях, чтобы получить больше солнечной энергии. Это называется листорасположением и передаётся по наследству. Но иногда условия освещения влияют на изменение этого признака.
Стебли растений разделены на узлы и междоузлия. Если в узлах по 1 листу, которые расположены по стеблю спиралью – очередной тип листорасположения. Встречается у яблони, ванили, пшеницы и у других подобных растений.
Если узел с 2 листами с разных сторон стебля – это супротивное расположение листьев. Пары листьев у сирени, крапивы, шалфея, клёна, занимают противоположные плоскости.
Когда узел окружён листьями больше 3-х штук как у хвоща лесного, элодеи, подмаренника, барбариса, такой тип называют мутовчатым листорасположением. При розеточном типе листья расположены вкруговую без видимого междоузлия и стебля, как у подорожника. Листовые пластинки расположены наподобие мозаики, чтобы не заслонять солнце друг для друга.
На верхней и нижней стороне листа слой кожицы, образованной покровной тканью с плотно лежащими клетками для защиты внутренней части. В этих клетках нет хлорофилла, и сквозь них свободно проходит солнечный свет. Благодаря жирообразным веществам, выделяемым кожицей, на поверхности образуется водонепроницаемая тонкая плёнка – кутикула. Кожица ни перегревается, ни повреждается и сильно не испаряет влагу, потому что покровные клетки покрыты волосками и шипиками.
Нижняя сторона пластинки «сухопутных» растений покрыта отверстиями-устьицами, которые образовали пары замыкающих клеток, похожие на подкову или боб. Плавающие листья – с устьицами наверху, у растений из влажных мест – на верхней и нижней стороне листа. На листьях подводных растений этих клеток нет.
Под кожицей листа скрывается главная ткань для образования мякоти листа с заключёнными хлоропластами. Верхняя часть мякоти – вертикальные столбчатые клетки. Ниже – слой паренхимы, образованной губчатой тканью из округлых клеток, между которыми много межклетников с воздухом. Этот воздух участвует в газообмене.
Паренхиму пронизывают сосудисто-волокнистые пучки (жилки), это сосуды: ситовидные трубки и трахеиды.
В хлоропластах проходит процесс с участием хлорофилла, энергии света, молекул воды и углерода, который отдаёт углекислый газ. Этот процесс называется фотосинтезом. Лист поглощает энергию солнца, а молекулы хлорофилла действуют на молекулы углекислого газа и воды. Так образуются молекулы глюкозы с выделением кислорода. Когда глюкоза расщепляется, энергия высвобождается с образованием углекислого газа и воды. Процесс фотосинтеза состоит из световой фазы – световая энергия превращается в химическую связь, и темновой, когда образуются органические соединения. За счёт транспирации:
- по сосудам тканей передвигается вода, доставляющая растворённые минеральные соединения;
- растение не перегревается;
- корни постоянно всасывают воду и поднимают по сосудам вверх.
Водяной пар испаряется через щели устьиц – 2 гибких, похожих на подковы, клетки. Смыкаясь, щель не выпускает влагу, при выгибании – открывают выход. Кутикулярная транспирация обеспечивает испарение через поверхности частей.
Дышат растения постоянно, без перерыва. Внутрь поступают порции углекислого газа, для участия в синтезе органических веществ и порции кислорода для дыхания. Наружу листья выделяют молекулы кислорода, высвободившиеся с фотосинтезом, и углекислого газа – продукт дыхания.
У берёзы, тополя, яблони, деревьев средней полосы, листья к зиме опадают. Эти деревья относятся к листопадным, а процесс называют листопадом. Со снижением температуры воздуха и уменьшением светового дня листья стареют и снижают обмен веществ. При разрушении хлоропластов с хлорофиллом приобретается другой цвет. Со сбрасыванием листьев удаляются накопившиеся ненужные вещества, сокращается испарение, а уменьшившееся количество веточек предохраняет растение от тяжести снега, чтобы побеги не ломались.
- Голосемянное растение, Вельвичия мирабилис или Вельвичия удивительная, эндемик пустыни Намиб, обзаводится только 2 листами, длиной по 3-8 м. Ремневидно-кожистые листья, отмирая на концах и постоянно нарастая основанием, живут по 100 с лишним лет. Это уже лохматые, порванные и спутанные «старички». Тонкие листья не запасают воду, как другие пустынные обитатели, а берут влагу из туманов.
- Иголки хвойных растений – видоизменённые листья. Уменьшившаяся площадь листовой пластинки сокращает испарение влаги из организма.
- Зелёная часть папоротника – не листья, а сросшиеся ветви.
- На кипарисе вырастает 50 млн. листьев, другие деревья наращивают гораздо меньше.
- Там, где жара, листовые пластинки к солнцу поворачивают ребро, а в холодных районах – плоскость листа.
- Дерево рафия отращивает листья длиной по 20-25 м, шириной по 3 м и это на стволе высотой 17 м. Народ Конго – бапенде использует гигантские листья для изготовления оригинальных народных костюмов, посмотреть на которые съезжаются туристы. Из вещества черешков и пластин плетут шляпы, корзины, коврики и даже ткань.
- Цветы олеандра, по красоте, затмевают розу, но сок одного листа этого растения настолько ядовит, что способен убить человека. Часто погибают животные, поев сочной зелени олеандра.
Благодаря строению листьев, хлорофиллу и воздушному питанию (фотосинтез) образующими органические вещества, Земля населена живыми организмами. Одни животные поедают растения, другие поедают тех, кто питается зеленью. Так организмы получают нужную для жизни солнечную энергию, которую самостоятельно запасать не могут.
Выполняя воздушное питание, листья выделяют в воздух кислород, вот почему в лесу легко дышать. Ежегодное выделение исчисляется тоннами – 140 млрд. Поражает разнообразие листьев, размеры, а осенняя окраска – чудо природы.
Строение плодового дерева и его частей: биологические и анатомические особенности
Для того чтобы выполнять обрезку плодовых деревьев не вслепую, а со знанием дела, чтобы предугадывать возможную реакцию конкретного растения на те или иные приемы формирования кроны, необходимо знать, как оно устроено и каковы закономерности его роста, развития и плодоношения.
В этой статье рассмотрены биологические и анатомические особенности строения плодового дерева и его надземных частей: ствола, ветвей, почек, листьев и цветов.
Ствол
Надземная часть взрослого плодового дерева представлена большим количеством различающихся по размеру, возрасту и назначению ветвей, составляющих в совокупности крону.
Основную стеблевую часть кроны, ее центральную ось, называют стволом. У одних деревьев ствол четко выделяется на протяжении всей их жизни. У других он на некоторой высоте отклоняется в сторону, разветвляется и теряется среди других ветвей. Место перехода ствола в корень в строении плодового растения называется корневой шейкой. Часть ствола от корневой шейки до первого ответвления — штамб, а выше — центральный проводник, или лидер. Ствол объединяет и удерживает на себе всю массу ветвей, листьев, плодов.
Ветви
На центральном проводнике размещены ветви первого порядка. Следует отметить, что центральный проводник плодоводы относят к нулевому порядку ветвления, а ботаники — к первому. Отсюда несовпадение в счете порядков ветвления. Наиболее крупные ветви считаются основными. Вместе с центральным проводником они составляют остов кроны. В специальной литературе вы можете встретить термины «скелетные ветви» или «сучья» вместо «основные ветви» и «скелет кроны» вместо «остов кроны». Ничего не поделаешь — традиция. Не судите строго. Не вдруг, не сразу формировалась терминология. Поэтому наряду с удачными есть и неудачные термины.
На ветвях первого порядка ветвления размещаются ветви второго порядка, на них — третьего и т. д. Ветвление может продолжаться до 7-8, а иногда и более высоких порядков. Но к основным ветвям относятся ветви первого, второго, редко третьего порядков ветвления. Ветви более высоких порядков ветвления обычно слабые, недолговечные и спустя некоторое время отмирают естественно или при обрезке заменяются на новые.
Все мелкие слаборослые разветвления, не входящие в остов кроны, называют обрастающими ветвями. Они, как правило, несут на себе большую часть урожая, поэтому их часто называют генеративными или плодоносными.
На этом рисунке представлено строение кроны плодового дерева:
Почки
На стеблях растений в пазухах листьев формируются почки. У семечковых они одиночные, у косточковых — и одиночные, и групповые, т. е. собранные в группы по 2-3, из которых бывают 1-2 вегетативные и 1-3 генеративные. У основания почек есть слаборазвитые, почти незаметные простым глазом, но всегда готовые к росту, так называемые запасные почки. Они трогаются в рост при гибели основной почки.
Почки, как правило, прорастают на второй год после их образования. Но есть и такие, которые трогаются в рост, едва успев сформироваться.
Это скороспелые почки. Они характерны для абрикоса, вишни, сливы, персика и некоторых других культур. У семечковых они также бывают, но гораздо реже. Некоторые почки трогаются в рост в исключительных случаях.
Это спящие почки. Они сохраняют способность к росту в течение многих лет и прорастают, например, при подмерзании растения, при его сильных механических повреждениях, при обрезке.
Почка — это побег в миниатюре, закрытый сверху чешуями, предохраняющими его от повреждений. На оси побега размещаются зачаточные листья, а в их пазухах — зачатки новых почек. Так устроены вегетативные почки. Если же, кроме того, в почке имеются зачатки цветков, то почки называют цветочными, или генеративными. Вернее было бы называть их вегетативно-генеративными, так как в них есть начало и вегетативных, и генеративных органов. Но опять-таки, такова терминология у садоводов. У ряда косточковых растений имеются и чисто генеративные почки, содержащие только зачатки цветков. Обычно генеративные почки, будь они простыми или смешанными, более крупные, чем вегетативные, округлые, оттопыренные. Из вегетативных почек образуются побеги.
После листопада побег утрачивает свое наименование и приобретает новое — ветвь. Все ветви по выполняемым ими функциям разделяются на вегетативные и генеративные.
Вегетативные ветви, образующиеся из верхушечных почек, увеличивают общую длину ветвей. Их поэтому называют ветвями продолжения. Из одной-двух почек, ближайших к верхушечной, вырастают побеги почти такой же длины, как и побег продолжения. Это конкуренты. Их назвали так потому, что они конкурируют за доминирующее положение с побегом продолжения. Из нижерасположенных почек вырастают побеги, различающиеся по длине и ориентации относительно основной оси ветви. Ими как бы обрастают все более или менее крупные ветви. Поэтому садоводы и назвали их обрастающими. В зависимости от культуры и сорта продолжительность жизни обрастающих ветвей 3-10 лет, хотя можно встретить и 15-летнюю обрастающую ветвь.
На многолетних частях крупных ветвей из спящих почек иногда появляются сильнорастущие вертикальные побеги. Это волчки, или жировые побеги.
Как видно на фото строения дерева, такие побеги, именуемые волчками, характеризуются удлиненными междоузлиями и крупными листьями:
Иногда побеги появляются у штамба, от корней. Это корнепорослевые побеги. Появление их обусловлено главным образом биологическими особенностями подвоев, но могут появляться вследствие ослабления надземной системы (старение, подмерзание, поломки).
Среди ветвей плодовых растений имеются такие, которые наиболее приспособлены к формированию генеративных почек. По этому признаку такие ветви, независимо от того, есть ли на них генеративные почки в данном году или нет, относят к генеративным. У семечковых пород это плодовый прутик, копьецо, кольчатка; у косточковых — букетные веточки, шпорцы, плодовые и смешанные ветви.
Плодовый прутик — тонкая, часто изогнутая книзу однолетняя ветвь длиной 15-25 см. Копьецо — прямая, заметно суживающаяся к верхушке ветвь длиной 5-15 см. Кольчатка — коротенькая ветвь длиной 0,3-5 см. Общим для всех этих ветвей являются укороченные междоузлия и слабое развитие боковых почек.
Букетная веточка имеет длину 0,5-3 см. На ней размещаются 3-10 почек, из которых 1-2 могут быть вегетативными, а все остальные генеративные. Шпорец— коротенькая копьецевидная ветвь (0,8-10 см). Верхушечная почка вегетативная, боковые — генеративные. Встречаются шпорцы, оканчивающиеся не почкой, а колючкой. Смешанная ветвь— однолетняя ветвь, несущая на себе генеративные и вегетативные боковые почки. Причем верхушечная почка обычно вегетативная. Длина смешанной ветви может быть различной. Плодовая ветвь по морфологическим признакам не отличается от смешанной, но все боковые почки — генеративные.
На плодовых прутиках, копьецах и кольчатках у деревьев многих сортов, главным образом семечковых культур, при пробуждении цветочной почки появляется вздутие — плодовая сумка. На верхушке сумки размещаются цветки (плоды), а нередко и 1-3 побега в виде кольчаток, копьец, плодовых прутиков или побегов вегетативного типа. Их называют побегами замещения.
Генеративные ветви, ежегодно прирастая в длину, превращаются в плодушки, или плодухи, состоящие из плодовых сумок, кольчаток, копьец, плодовых прутиков. Более старые и разветвленные плодухи называют сложными плодовыми, или смешанными обрастающими ветвями.
Лист
Весь круговорот веществ в природе (исключения очень редки) начинается с листа. Лист — это уникальнейшее творение природы. Благодаря листу солнечная энергия связывается в органическом веществе и становится доступной для использования человеком в его жизнедеятельности.
Под верхней кожицей листа находятся удлиненные, напоминающие столбики, тесно подогнанные одна к другой клетки. Это столбчатая, или па лисадная, паренхима. В ее клетках находятся хлоропласты. По форме они напоминают зерна злаковых, но гораздо мельче. В каждой клетке листа яблони, например, может быть до 10-15 хлоропластов.
Хлоропласты благодаря своей структуре и содержащемуся в них хлорофиллу способны поглощать энергию солнечного света и в процессе синтеза передавать ее тому или другому элементу или промежуточному веществу.
С нижней стороны листа расположена губчатая паренхима, или губчатый мезофилл. Она состоит из сравнительно крупных клеток овальной формы. В них также имеются хлоропласты, но их меньше, и они не так активно участвуют в процессе синтеза органического вещества, как хлоропласты столбчатой паренхимы. Да это им и не нужно. Другие функции у них — воздухообмен и транспирация. Между клетками губчатой паренхимы имеются крупные промежутки — резервуары для воздуха и паров воды. На 1 мм2 нижней части листа размещается 200-700 устьиц, через которые и осуществляются эти функции губчатого мезофилла.
За день 1 м2 листьев может синтезировать 10 г и более органического вещества. Чтобы были хорошие урожаи плодов, листья должны быть здоровыми, получать достаточно света, влаги и элементов минерального питания. Тогда и синтез органического вещества будет интенсивным.
Цветки
Цветки у плодовых растений бывают обоеполыми (семечковые, косточковые), когда в одном цветке имеются и тычинки, и пестик. Могут быть и однополыми (клубника, облепиха), когда в одних цветках только мужские, а в других — только женские органы. Цветки бывают одиночными (персик, абрикос, айва) и собранными в соцветия: зонтик (яблоня, черешня, вишня), щиток (груша, арония, рябина), кисть (смородина, крыжовник, малина), дихазий (земляника).
Раскрываются цветки не одновременно, как в пределах растения, так и в пределах ветви и отдельного соцветия. У яблони, например, первыми раскрываются центральные цветки соцветия, у груши — периферийные, у земляники — нижние. Из первых раскрывшихся цветков, как правило, формируется основная часть урожая.
Клеточное строение ветвей
При обрезке, прививке, определении степени подмерзания дерева и в ряде других случаев необходимо знать размещение и назначение основных тканей, из которых состоят ветви растения.
На поперечном срезе ветви в строении дерева можно видеть неоднородность тканей. В центре располагается сердцевина. Она состоит из сравнительно рыхло размещающихся паренхимных клеток. Осенью в них накопляются запасы питательных веществ, за счет которых в период покоя и в первое время после начала вегетации осуществляются все процессы жизнедеятельности растения и обеспечивается начальный рост новых вегетативных и генеративных образований.
Вокруг сердцевины кольцами размещается древесина (ксилема). Количество колец древесины соответствует возрасту ветви, поскольку в каждый год нарастает одно кольцо. Кольца образуются в связи с тем, что весной формируются крупные клетки, а осенью — мелкие. Соответственно, в клеточном строении дерева образуется резкая граница между соседними кольцами древесины. Толщина кольца свидетельствует также об условиях роста ветви в данном году. Древесина состоит из разнообразных сосудов и трахеид, по которым от корневой системы ко всем частям надземной передвигается влага с растворенными в ней элементами минерального питания (восходящий ток). В древесине имеются также толстостенные клетки механического строения, обеспечивающие прочность дерева; клетки древесинной паренхимы, в которых накапливаются запасы, и клетки сердцевинных лучей, по которым перемещаются питательные вещества в поперечном направлении.
Слой древесины окружен кольцом камбия. Простым глазом эту ткань рассмотреть невозможно, так как она состоит всего лишь из одного слоя клеток. Клетки камбия обладают способностью делиться. При делении внутрь откладываются клетки древесины, а наружу — клетки луба. При частичной, а иногда и при полной гибели древесины ветвь может восстановиться, если камбий останется живым. За счет его деятельности создадутся новые слои древесины и луба.
Луб состоит главным образом из ситовидных трубок с сопровождающими клетками. По этим сосудам перемещаются продукты ассимиляции из листьев в другие органы надземной и корневой системы (нисходящий ток). Кроме того, луб содержит паренхимные клетки, в которых размещается часть запасных веществ, и механические ткани — лубяные волокна.
Защищаются все указанные ткани корой, верхний слой которой представлен пробковыми тканями, а более глубокий — паренхимными (запасающими) клетками.
Поскольку в растении наблюдается постоянное перемещение влаги с растворенными в ней элементами минерального питания и продуктов ассимиляции, обеспечение нормального развития транспортных путей является одной из главных задач обрезки. Хорошей проводящей способностью обладают длинные, сильные годичные приросты. Укороченные «тормозят» перемещение веществ по растению. Плохими транспортными способностями характеризуются также подмерзшие ткани, с механическими повреждениями, а также поврежденные вредителями и болезнями.
На этой схеме представлено внутреннее строение дерева в разрезе:
Листья растений
Содержание
Листья – основные фотосинтезирующие органы растений. На развитом побеге они инициируются как листовые примордии в апикальных меристемах. Увеличиваются за счёт деления и растяжения клеток краевых и плоскостных вставочных меристем, а рост их останавливается с наступлением зрелости. Исключение – вельвичия удивительная, листья которой растут всю жизнь за счёт вставочной меристемы, расположенной у оснований. Длительное время нарастают верхушками листья папоротника – вайи.
Как и любой орган, листья имеют определённую структуру. Их расположение, размер, форма и даже внутреннее строение различаются не только у разных видов и родов, но даже у одного растения. Всё это имеет приспособительное значение и зависит от среды обитания и места расположения.
Простые и сложные листья растений
Листья могут быть простыми (состоящими из одной пластинки, которая при листопаде опадает целиком) и сложными. В сложных листьях, например у бузины, грецкого ореха, рябины, пластина разделена на листочки, каждый из которых самостоятельно прикрепляется к обшей оси, а часто даже имеет свой черешок. Листочки сложного листа отрываются отдельно. Общая ось, на которой расположены листочки, называются рахисом. Сложные листья очень разнообразны. В зависимости от расположения на рахисе листочков они бывают:
- тройчатосложными;
- пальчатосложными;
- перистосложными.
Перистосложные листья с нечётной листовой пластинкой на верхушке называют непарноперистыми, а с чётным числом листочков – парноперистыми.
Процесс формирования сложного листа напоминает ветвление, которое может идти до второго-третьего порядка. Тогда образуются листья дважды- и трижды-перисторассечённые, многократно тройчатые и т. д.
Форма листовой пластинки
Форма листовой пластинки – важный систематический признак. Она бывает очень разной. При оценке морфологии листовой пластины смотрят главным образом на её верхушку и основание.
Основание листа может быть почти незаметным или иметь вид небольшого утолщения (подушечки), например у кислицы. Часто основание сильно разрастается в ширину и длину, охватывая узел целиком, и образуя трубку, называемую влагалищем листа. Образование влагалища характерно для многих однодольных, в частности для злаков, из двудольных оно встречается у зонтичных. Влагалища часто защищают почки, стебли, зачаточные побеги и цветы.
Иногда основание листа формирует раструб, который можно расценивать либо как вырост влагалища, либо как результат слияния двух пазушных прилистников. Раструб характерен для всех видов семейства гречишных.
Различают листья и по характеру края листовой пластинки. У берёзы, сирени они простые неразделённые, сплошные — цельнокройные. Но простые листья могут иметь также зубчики, лопасти, доли или сегменты – выемки различных размеров, как у клёна или дуба. Зубчатый край способствуют большей интенсивности фотосинтеза. Если вырезки на листе заходят не глубже, чем на ¼ их ширины, то листовые пластинки называют цельными с изрезанным краем. Изрезанность листовой пластинки – это приспособление для уменьшения воздействия ветра. Если край изрезан глубже, то пластинки называются расчленёнными. В результате получаются перисто- и пальчато-лопастные, перисто- и пальчато-раздельные, перисто- и пальчато-рассечённые листья.
Листорасположение
Листорасположение, или филлотаксис – это порядок размещения листьев на оси побега. Различают несколько основных вариантов листорасположения:
- спиральное, или рассеянное (очерёдное) – на каждом узле расположен один лист и основания листьев одной оси последовательно можно соединить условной спиральной линией: растянутой, если стебель удлинённый, и плоскостной, если он укороченный;
- двурядный вид листорасположения, который можно рассматривать как частный случай спирального. Отражает маятниковую симметрию деятельности апекса. На каждом узле находится один лист, охватывающий основанием всю или почти всю окружность. Средняя линия всех листьев лежит в одной вертикальной плоскости;
- мутовчатое – появляется, если на одном уровне закладывается несколько листовых примордиев, образующих общий узел. Нередко при близком изучении оказывается, что каждый лист мутовки имеет собственный узел, но они сильно сближены;
- супротивное листорасположение – частный случай мутовчатого, когда на одном узле образуется два листа точно напротив друг друга. Часто такое расположение бывает накрест супротивным, т. е. плоскости соседних пар листьев являются взаимно перпендикулярными.
Хотя тип расположения листьев – это наследственный признак, однако он зависит от среды обитания и в процессе роста растения может меняться. Благодаря неравномерности роста стебель может скручиваться вокруг своей оси. Для сохранения симметрии размещения листьев по стеблю их черешки могут изгибаться, поворачивая листья так, что по их расположению уже не удаётся определить исходную формулу филлотаксиса.
Особенно ярко это выражается на листовой мозаике. При этом листья выстраиваются горизонтально, подставляя пластинки свету, так что становятся одной плоскостью. Листовая мозаика способствует максимальному использованию рассеянного света. Её можно наблюдать на горизонтальных ветвях в кроне липы, на побегах плюща, герани, подорожника, табака и т.д.
Листовая мозаика
Внутреннее строение листа
Внутренняя структура листовой пластинки приспособлена для фотосинтеза, газообмена и испарения воды. Вся поверхность листа покрыта прозрачной эпидермой, большинство клеток которой не имеет хлоропластов. Эпидерма верхней стороны листовой пластины содержит восковой кутикулярный слой, препятствующий испарению воды и отражающий солнечные лучи, на нём могут присутствовать железистые волоски и трихомы. Трихомы удерживает влагу и препятствуют её испарению. Эпидерма выполняет несколько функций:
- защита от излишнего испарения;
- регуляция газообмена для дыхания и фотосинтеза;
- выделение воды и некоторых веществ;
- впитывания воды (у некоторых растений, не у всех).
Слой эпидермы на нижней стороне большинства листьев содержит щелевидные отверстия (устьица), с расположенными по бокам замыкающими клетками. При равном освещении обеих сторон листа, устьица образуются на обеих из них. У плавающих в воде листьев устьица есть только на верхней эпидерме. Устьица регулируют газообмен и испарение, они связаны с межклетниками основной ткани листа.
Основная ткань между верхней и нижней эпидермой называется мезофиллом. Мезофилл – важнейшая ткань листа, в её клетках сосредоточены хлоропласты и происходит фотосинтез. Она перемежается жилками различных размеров. Клетки мезофилла покрыты тонкой оболочкой, они не имеют одревесневшей клеточной стенки.
Большинство листьев папоротников и цветковых растений имеет два различных типа мезофилла:
- верхний, столбчатый (палисадный) – состоящий из одного или нескольких (чаще двух) рядов плотноупакованных бочкообразных или цилиндрических вытянутых клеток хлоренхимы (паренхима с хлоропластами). Они расположены прямо под эпидермой вертикально по отношению к ней. Листья, растущие на солнце, содержат до 5 слоёв палисадного мезофилла, в теневых листьях есть только 1 слой. Некоторые растения, например виды Эвкалиптов из-за особого расположения их листьев по отношению к свету (боком) содержат столбовидную хлоренхиму ближе к краям листовой пластинки.
- губчатый – пространство между столбчатой хлоренхимой и нижним эпидермисом заполнено рыхлой паренхимой, между клетками которой имеется множество воздушных пространств. Эти воздушные полости взаимосвязаны с устьицами и участвуют в газообмене и выведении водяного пара из листа. Увеличение межклеточных пространств достигается различными путями: в одних случаях клетки сохраняют округлую форму, в других образуют выросты.
Расположение устьиц преимущественно на нижней стороне листа объясняется не только положением губчатого мезофилла. Потеря воды листом в процессе транспирации идёт медленнее через устьица, расположенной в нижней, а не в верхней эпидерме. Кроме того, главным источником углекислого газа в атмосфере является «почвенное дыхание» — выделение СО2 в результате жизнедеятельности многочисленных живых существ, населяющих почву.
Абсолютная толщина палисадной и губчатой ткани и число слоёв клеток в них различны, в зависимости от освещения и других причин. Даже у одной особи, например на одном кусте сирени, листья, выросшие на свету, имеют более развитый мезофилл, чем теневые листья.
Внутреннее строение листьев может меняться. Если нижняя сторона листьев получает достаточно света, то и на ней образуется столбчатый мезофилл. У многих листьев однодольных растений мезофилл не дифференцируется на столбчатый и губчатый, а состоит из одинаковых клеток. Встречаются уклонения от типичной плоской структуры листа и тогда клеточное строение тоже меняется. У некоторых растений-ксерофитов обе стороны листа имеют одинаковую эпидерму и мезофилл. У многих суккулентов листья цилиндрической формы с радиальной симметрией. У некоторых злаков имеется особенно высокоспециализированный тип мезофилла – корончатый. Здесь клетки мезофилла окружают проводящие пучки, примыкая к ним по радиусу. В промежутках между клетками имеются большие межклетники, против которых с обеих сторон имеются устьица.
Видоизменённые листья растений
Когда растения колонизировали широкий спектр сред от пустынь до тропических лесов, возникали и изменения их органов для приспособления к специфике климата. В связи с приспособлением к выполнению новых функций многие листья изменились и стали непохожими на листья в привычном для нас виде. Краткое описание некоторых их адаптаций следует ниже.
- Хвоя – листья голосеменных хвойных растений. Они стали такими из-за суровости климата в местах произрастания.
- Прицветники, верховые, или цветочные листья. У кизила и пуансеттии есть относительно незаметные, небольшие, зеленовато-жёлтые цветки. Оба растения производят большие, яркие прицветники, у пуансеттии они окрашены в красный или белый цвета, у кизила – в розовый. Эти прицветники окружают настоящие цветы и выполняют функцию лепестков, привлекая насекомых-опылителей.
- Шипики (колючки). Листья или их части многих кактусов и других растений превращаются в шипы. Острые колючки могут сдерживать растительноядных животных и уменьшать степень испарения воды за счёт уменьшения площади собственной поверхности. У барбариса они острые 3-7-раздельные, лишённые мезофилла. У белой акации шипы образовались из прилистников. Листовые колючки не следует путать с колючками, произошедшими из видоизмененных стеблей, как у гледичии трёхколючковой (Gleditsia triacanthos) или с шипами малины, представляющими простые выросты эпидермиса или коры. Отличие: колючки, произошедшие от листа, растут из пазух и имеют рядом пазушную почку. У колючек, появившихся из стебля, почки могут быть только на них самих, а не в пазухах.
- Усики. Превращение части листовой пластинки в усики характерно для растений семейства бобовых. У видов чины, гороха, вики усики образованы из центральной жилки листочка или из жилок нескольких листочков (ветвистые усики). Усики чины прилистниковой появляются из прилистников, а её крупные прилистники выполняют роль листовой пластины. Усики также могут образовываться из черешка листа.
- Чешуйки. У многих растений листья превращаются в чешуйки, например у туи. Толстые, мясистые чешуйки луковицы служат для запасания веществ. Почечные чешуи выполняют защитную функцию, а листья-чешуйки саксаула способствуют уменьшению транспирации.
- Филлодии. У растений засушливых мест обитания, например у австралийских акаций листья редуцированы, а их роль выполняют филлодии – уплощённые листовые черешки.
- Пестик. Это тоже видоизменённые листья.
- Репродуктивные листья. Некоторые растения, например каланхоэ, производят крошечные, но цельные саженцы по краям листьев. Каждый такой саженец, отделяется от листа и вырастает в полноразмерное растение. Американский ходячий папоротник (Asplenium rhizophyllum) производит новые ростки на кончиках листьев.
- Форточки. Некоторые растения, обитающие в пустынях, образуют мясистые листья с прозрачными краями. Сами листья оказываются погребёнными в песок, а прозрачные кончики остаются снаружи. Они снабжены кутинизированным эпидермисом и могут впускать свет внутрь листа. Эта стратегия помогает фотосинтезу происходить под землёй.
- Мешковидные листья. Срастаются так, что образуют структуру, похожую на мешок, способную удерживать воду. Такое приспособление есть у эпифитов и лиан.
- Суккулентные листья – мясистые, служат для запасания воды у растений засушливых мест обитания.
- Насекомоядные листья. Известно почти 200 видов цветковых растений, листья которых приспособлены к ловле насекомых. Растения с насекомоядными листьями чаще растут на кислых болотистых почвах, в которых отсутствуют азот и другие необходимые для жизни элементы или они содержатся в недоступных для растения формах. Поэтому они и восполняют недостаток за счёт насекомых.
Ловчие приспособления разнообразны: пузырьки с клапанами у пузырчатки, липкие желёзки, захлопывающиеся половинки листа и др.
Растения с листьями в форме кувшинов (Саррацения, Дарлингтония или виды Непентеса, или Кувшиночника) накапливают внутри этих резервуаров воду. Внутренняя поверхность листьев очень гладкая, но по их краю спускаются жёсткие волоски. Из такого листа, упавшим насекомым сбежать трудно и они в конце концов тонут. Собственные или бактериальные ферменты разлагают тело насекомого.
Другие растения, такие как росянка (Drosera), имеют желёзки, выделяющие липкую слизь. Она ловит насекомых, которые затем перевариваются ферментами. Листья Венериной мухоловки (Dionaea muscipula) в середине как-будто соединены при помощи шарнира. Когда насекомые задевают крошечные волоски на их листовых пластинках, две половинки листа захлопываются, и пищеварительные ферменты разлагают мягкие части насекомого до такого состояния, в котором они могут быть поглощены через поверхность листа. Интересно, что венерина мухоловка не способна жить в богатой азотом среде. Видимо это стало результатом сложного биохимического процесса, который и развил у растения способность улавливать и переваривать насекомых.
Страница не найдена
Размер:
AAA
Цвет: C C C
Изображения Вкл. Выкл.
Обычная версия сайта
RUENBY
Гомельский государственный
медицинский университет
- Университет
- Университет
- Руководство
- История
- Устав и Символика
- Воспитательная деятельность
- Организация образовательного процесса
- Международное сотрудничество
- Система менеджмента качества
- Советы
- Факультеты
- Кафедры
- Подразделения
- Первичная профсоюзная организация работников
- Издания университета
- Гордость университета
- Выпускник-2021
- Первичная организация «Белорусский союз женщин»
- Одно окно
- ГомГМУ в международных рейтингах
- Структура университета
- Абитуриентам
- Приёмная комиссия
- Целевая подготовка
- Заключение, расторжение “целевого” договора
- Льготы для молодых специалистов
- Архив проходных баллов
- Карта и маршрут проезда
- Порядок приёма на 2022 год
- Специальности
- Контрольные цифры приёма в 2022 году
- Стоимость обучения
- Информация о ходе приёма документов
- Приём документов и время работы приёмной комиссии
- Порядок приёма граждан РФ, Кыргызстана, Таджикистана, Казахстана
- Pепетиционное тестирование
- Горячая линия по вопросам вступительной кампании
- Студентам
- Первокурснику
- Расписание занятий
- Расписание экзаменов
- Информация для студентов
- Студенческий клуб
- Спортивный клуб
- Общежитие
- Нормативные документы
- Практика
- Стоимость обучения
- Безопасность жизнедеятельности
- БРСМ
- Профком студентов
- Учебный центр практической подготовки и симуляционного обучения
- Многофункциональная карточка студента
- Анкетирование студентов
- Выпускникам
- Интернатура и клиническая ординатура
- Докторантура
- Аспирантура
- Магистратура
- Распределение
- Врачам и специалистам
- Профессорский консультативный центр
- Факультет повышения квалификации и переподготовки
- Иностранным гражданам
- Факультет иностранных студентов
- Стоимость обучения
- Регистрация и визы
- Полезная информация
- Правила приёма
- Информация о возможностях и условиях приема в 2022 году
- Официальные представители ГомГМУ по набору студентов
- Страхование иностранных граждан
- Приём на Подготовительное отделение иностранных граждан
- Прием иностранных граждан для обучения на английском языке / Training of foreign students in English
- Повышение квалификации и переподготовка для иностранных граждан
- Научная деятельность
- Направления научной деятельности
- Научно-педагогические школы
- Студенчеcкое научное общество
- Инновационные технологии в ГомГМУ
- Научно-исследовательская часть
- Научно-исследовательская лаборатория
- Конкурсы, гранты, стипендии
- Работа комитета по этике
- Научные мероприятия
- В помощь исследователю
- Диссертационный совет
- «Горизонт Европа»
- Патенты
- Инструкции на метод
- Совет молодых ученых
- Госпрограмма (ЧАЭС)
- Главная
30.
8: Листья — структура и расположение листьев- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 13753
- Безграничный
- Безграничный
Цели обучения
- Эскиз базовой структуры типичного листа
Структура типичного листа
Каждый лист обычно имеет листовую пластинку, называемую листовой пластинкой, которая также является самой широкой частью листа. Некоторые листья прикреплены к стеблю растения черешком. Листья, не имеющие черешка и непосредственно прикрепленные к стеблю растения, называются сидячими. Листья также имеют прилистники, небольшие зеленые придатки, обычно находящиеся у основания черешка. У большинства листьев есть средняя жилка, которая проходит по всей длине листа и разветвляется с каждой стороны, образуя жилки сосудистой ткани. Край листа называется поле .
Внутри каждого листа сосудистая ткань образует жилки. Расположение жилок в листе называется рисунком жилкования . Однодольные и двудольные отличаются рисунком жилкования. Однодольные имеют параллельное жилкование, при котором жилки проходят прямыми линиями по всей длине листа, не сходясь. Однако у двудольных жилки листа имеют сетчатый вид, образуя рисунок, известный как сетчатое жилкование. Ginkgo biloba является примером растения с дихотомическим жилкованием.
Расположение листьев
Расположение листьев на стебле известно как филлотаксия . Количество и расположение листьев растения будет варьироваться в зависимости от вида, при этом каждый вид имеет характерное расположение листьев. Листья классифицируются как очередные, спиральные, супротивные или мутовчатые. Растения, у которых есть только один лист в узле, имеют листья, которые называются очередными или спиральными. Очередные листья чередуются с каждой стороны стебля в плоской плоскости, а спиралевидные листья располагаются по спирали вдоль стебля. При супротивном расположении листьев два листа возникают в одной и той же точке, при этом листья соединяются друг напротив друга вдоль ветви. При наличии трех и более листьев, соединенных в узле, расположение листьев классифицируется как мутовчатое.
Ключевые моменты
- Каждый лист обычно имеет листовую пластинку (пластинку), прилистники, среднюю жилку и край.
- Некоторые листья имеют черешок, которым лист прикрепляется к стеблю; листья, не имеющие черешков, прикрепляются непосредственно к стеблю растения и называются сидячими.
- Расположение жилок в листе называется рисунком жилкования; однодольные имеют параллельное жилкование, а двудольные – сетчатое.
- Расположение листьев на стебле известно как филлотаксия; листья можно разделить на очередные, спиральные, супротивные или мутовчатые.
- Растения с очередным и спиральным расположением листьев имеют только один лист в узле.
- В противоположном расположении листьев два листа соединяются в узле. При мутовчатом расположении три или более листьев соединяются в узле.
Ключевые термины
- черешок : черешок, который простирается от стебля до основания листа
- листовая пластинка : плоская часть листа; лопасть, являющаяся самой широкой частью створки
- прилистник : небольшой зеленый придаток, обычно находящийся у основания черешка
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или Страница
- Автор
- Безграничный
- Количество столбцов печати
- Два
- Печать CSS
- Плотный
- Лицензия
- CC BY-SA
- Версия лицензии
- 4,0
- Показать оглавление
- нет
- Теги
- поля
- филлотаксия
- рисунок жилкования
Анатомия листа – Удаление дерева
Деревья не только красивы своими величественными кронами листьев, но и являются важной частью здоровой окружающей среды. Дерево состоит из нескольких частей, составляющих его целое. Корни под землей помогают закрепить дерево и питаются. Ствол и ветви поддерживают структуру дерева. Цветы в конечном итоге дадут плоды и семена по мере развития вегетационного периода. Листья также являются важной частью анатомии дерева, потому что они поглощают солнечный свет и производят пищу для дерева. Когда листья поглощают солнечный свет, происходит фотосинтез, который обеспечивает дерево энергией. Изучение анатомии деревьев и листьев может помочь вам понять, как функционируют деревья.
Структура листа
Листья имеют разную форму. Простые листья не разделены на отдельные отделы и имеют одну пластинку. Эти листья могут иметь сердцевидную, круглую, перьевую, ладонь, почку или другую форму. Лопастные листья имеют углубления, и они могут иметь форму ладони или пера. Сложные листья имеют отдельные листочки. У одних деревьев четное количество листочков, у других нечетное.
Внутренняя анатомия листьев сложная, многослойная. Наружные слои листа призваны защищать его, служа прочным барьером. Под барьерными слоями у листа есть специальные слои, которые позволяют ему обрабатывать солнечный свет. Листья имеют особую структуру и форму в зависимости от среды, в которой они растут.
- Хлоропласты: Хлоропласты отвечают за производство пищи для растительных клеток. Они поглощают световую энергию и начинают процесс ее преобразования в сахара.
- Кутикула: Кутикула — это покрытие вокруг листа, которое помогает остановить потерю воды. Кутикула гладкая, восковая.
- Эпидермис: Эпидермис — это внешняя оболочка листа, которая помогает защитить ткани под ним.
- Защитные клетки: Эти специальные клетки работают, чтобы открывать и закрывать устьица листа, чтобы поддерживать необходимое количество влаги в листе.
- Листовые волоски: У некоторых листьев есть листовые волоски на эпидермисе, которые помогают защитить лист.
- Столбчатый слой: Столбчатый слой — это слой листа с хлоропластами. Фотосинтез происходит в палисадном слое.
- Губчатый мезофилл: Расположенный под палисадным слоем губчатый мезофилл помогает перемещать кислород и углекислый газ через лист.
- Устьица: Устьица представляют собой небольшие поры в эпидермисе, которые помогают листьям обмениваться газами. Большинство листьев имеют устьица на нижней стороне.
- Сосудистый пучок: Эти жилки листа содержат ткани флоэмы и ксилемы. Ткань флоэмы перемещает большие молекулы по растению. Ткань ксилемы перемещает более мелкие молекулы воды и питательных веществ через растение.
Функция листа и фотосинтез
Эпидермис листа имеет крошечные поры, называемые устьицами. Эти поры представляют собой отверстия, которые принимают и выделяют газы. Многие листья закрывают устьица в ночное время, чтобы листья не теряли слишком много влаги. Поскольку листья выделяют молекулы воды через устьица, они помогают охлаждать воздух. Выделение влаги называется транспирацией, и это одна из причин, почему деревья полезны для окружающей среды.
Фотосинтез — это процесс, при котором листья производят пищу для растения. Углекислый газ и вода попадают в устьица через нижнюю сторону листьев. Когда солнечный свет попадает на верхушки листьев, хлоропласты начинают процесс преобразования солнечного света в энергию для питания растения. Хлоропласты содержат пигмент, называемый хлорофиллом, который является жизненно важным ингредиентом, необходимым для фотосинтеза. Молекулы хлорофилла поглощают энергию света, обеспечивая фотосинтез. Двумя побочными продуктами фотосинтеза являются углеводы и кислород. Растение будет использовать углеводы в качестве пищи, чтобы поддерживать его.
Листья выделяют кислород через устьица в окружающую среду, где он используется другими живыми организмами. Чтобы произвести одну молекулу глюкозы для питания дерева, лист должен поглотить шесть молекул углекислого газа. В свою очередь, лист затем выпускает шесть молекул кислорода обратно в окружающую среду. Одно среднее дерево производит около 260 фунтов кислорода в год.
- Анатомия листьев
- Листовая структура
- Хлоропласты: покажи мне зеленый
- Структура листа и функция
- Анатомия и виды деревьев
- Базовая анатомия растений
- Терминология: характеристики листьев, веток и плодов
- Биология деревьев (PDF)
- Анатомия дерева
- Анатомия и физиология листьев (PDF)
- Понимание анатомии и морфологии листа (PDF)
- Лист: место фотосинтеза и транспирации
- Структура листа и функция
- Структура листа
- Структура листа и приспособления к фотосинтезу
- Анатомия дерева (PDF)
- Древовидная структура и функции (PDF)
- Анатомия дерева (PDF)
- Знай свои деревья (PDF)
- Идентификация деревьев (PDF)
Что такое лист? · Frontiers for Young Minds
Abstract
Когда думаешь о растениях, на ум неизбежно приходит зеленый цвет. Как, пожалуй, самая заметная и легко узнаваемая часть мира природы, листья украшают мир зеленью. Если вы внимательно присмотритесь к растениям в своем дворе или на своей улице, вполне вероятно, что вы не найдете двух совершенно одинаковых. Это во многом благодаря огромному разнообразию форм, текстур и даже цветов листьев! Все листья имеют общие характеристики, которые мы рассмотрим в этой статье, но некоторые из них обладают особыми способностями. Некоторые листья изменены так, чтобы они выглядели как цветы, другие модифицированы, чтобы помочь растению карабкаться, защитить растение от потенциальных угроз или даже действовать как ловушки для насекомых. Мы хотим поделиться с вами необыкновенным миром листьев: продолжайте читать, и вы узнаете множество форм, структур и невероятных функций листьев.
Из чего состоит лист?
Листья состоят из трех основных частей: черешка, основания и листовой пластинки (рис. 1А). Как правило, самая большая часть листа – это лезвие. Основание – это область лезвия, которая прикрепляется к черешку, стеблевидной структуре, которая соединяет лезвие листа со стеблем растения. У некоторых листьев отсутствуют черешки, и они известны как сидячие (неподвижные) листья.
- Рисунок 1 – Внешнее и внутреннее строение створки.
- (A) Верхняя сторона листа. Здесь черешок соединяет лист со стеблем. (B) Поперечное сечение этого листа показывает различные слои клеток; зеленые пятнышки внутри клеток – это хлоропласты. (C) Устьица, структуры, которые открываются и закрываются для обеспечения газообмена с окружающей средой. (D) Сложный пальмовый лист, состоящий из нескольких листочков. (E) Листья супротивные, растущие прямо напротив друг друга на стебле. (Ж) Очередные листья, растущие один за другим на стебле.
Листовая пластинка образована несколькими слоями клеток. Растительные клетки относительно большие и окружены клеточной стенкой (рис. 1В). Клетки листа заполнены хлоропластами, структурами, которые содержат специализированные пигменты, называемые хлорофиллами. Хлорофиллы поглощают свет, позволяя растениям собирать энергию солнца. Листья зеленые, потому что хлорофиллы поглощают все цвета ультрафиолетового света, кроме зеленого. Не поглощая зеленый цвет, они вместо этого отражают его, что затем видят наши глаза. Листья имеют восковой наружный слой, покрытый устьицами, похожими на рты, которые открываются и закрываются для обеспечения газообмена с окружающей средой (рис. 1С). Листья должны соблюдать баланс между открытием устьиц, достаточным для обеспечения газообмена, и сохранением их закрытыми, чтобы предотвратить потерю воды.
Листья имеют жилки, которые проходят через черешок к остальной части растения. Эти вены состоят из сосудистых тканей, которые являются структурными тканями, транспортирующими воду и питательные вещества ко всем частям растения. Если вы забудете полить растение в течение нескольких недель, вы можете заметить, что листья начинают увядать. Однако, если вы добавите немного воды в почву, вода будет перенесена на весь путь к листьям, и они снова станут вертикальными.
Листья могут быть большими или маленькими, симметричными или асимметричными, с зубчатыми или гладкими краями, блестящими или шероховатыми. Листья в целом делятся на два типа. Простые листья состоят из одной пластинки. Сложные листья состоят из нескольких лопастей, которые называются листочками. Расположение листьев на стебле может быть супротивным, например, когда два листа растут прямо напротив друг друга, или чередующимся, когда листья чередуют стороны, один за другим (рис. 1D–F).
Жизненный цикл листа
Многие листья развиваются в защищенных структурах, называемых листовыми почками. Новые листья окружены защитными чешуйками, которые позволяют хрупким листьям отдыхать, пока они не будут готовы расшириться. Эти чешуи представляют собой видоизмененные листья, которые не фотосинтезируют. Некоторые почки лишены этих защитных чешуек и называются голыми почками [1].
На протяжении жизни листья увеличиваются в размерах, а также могут менять цвет. В некоторых частях мира осенний сезон известен кучами оранжевых, желтых, красных и коричневых листьев. Осенью клетки листа накапливают больше светопоглощающих пигментов, называемых каротиноидами. Каротиноиды отражают цвета, такие как желтый и оранжевый. Эти пигменты обычно находятся в более низких концентрациях, чем хлорофилл, поэтому они обычно маскируются зеленым цветом. Деревья могут оценивать температуру и продолжительность светового дня. Поскольку осенью оба вещества уменьшаются, хлорофилл начинает разлагаться, и могут просвечиваться желтые и оранжевые каротиноиды [2].
Когда деревья теряют листья, они не умирают. Прежде чем опадают листья, деревья переносят питательные вещества из листьев в другие части растения, чтобы сохранить эти питательные вещества для последующего использования. Изменения в листьях ослабляют связь между черешком и стеблем, и в конце концов листья естественным образом отделяются от дерева. Возможно, вы слышали термин «вечнозеленое дерево», относящийся к деревьям, которые остаются зелеными зимой. Некоторые вечнозеленые хвойные деревья (например, сосны) сохраняют свои листья как минимум целый год, прежде чем постепенно сбросить старые иголки, чтобы освободить место для новых.
Функция листьев
Поскольку растения не могут двигаться, они не могут пойти на кухню и приготовить бутерброд, когда проголодались. Вместо этого они производят свою собственную еду. Организмы, которые делают это, называются «автотрофами». Листья служат основным местом для производства продуктов питания посредством процесса, называемого фотосинтезом, который происходит в хлоропластах. Во время фотосинтеза свет, вода и углекислый газ из атмосферы поглощаются и превращаются в энергию в виде сахаров. В результате фотосинтеза вода расщепляется на водород и кислород. Растения выделяют дополнительный кислород через устьица, что способствует воздуху, которым мы дышим. Структура листа идеально спроектирована для его роли в фотосинтезе. Лезвие состоит из клеток, заполненных хлоропластами, которые улавливают свет в течение дня. У большинства растений устьиц больше на нижней стороне листа, чтобы уменьшить потерю воды при газообмене.
Растения выделяют много других молекул, которые взаимодействуют с окружающей средой. Некоторые растения издают сигналы, когда они повреждены. Например, сладкий запах свежескошенной травы является сигналом бедствия, а также защитным механизмом от атак травоядных. Подобные сигналы могут либо привлекать хищных видов для травоядных, либо привлекать полезных насекомых (таких как осы-паразиты, отбивающиеся от травоядных), либо предупреждать соседние растения, чтобы они укрепляли свою химическую защиту [3].
Некоторые листья модифицированы для других функций, кроме фотосинтеза. Например, у вьющихся растений, таких как огурцы, некоторые листья скручиваются и образуют усики, которые помогают растению прикрепляться к опоре во время подъема (рис. 2А). Иногда листья обманывают нас, заставляя думать, что они являются частью цветка, превосходно имитируя лепестки (рис. 2E, F). Пример этого можно найти в цветках гортензии. Эти модифицированные листья, называемые прицветниками, простираются под настоящим цветком. Их эффектная окраска привлекает опылителей к цветам, которые они поддерживают (рис. 2E, F). В следующий раз, когда вы увидите гортензию, посмотрите, сможете ли вы отличить прицветники от настоящих цветов. Колючки кактусов также представляют собой листья, которые были сильно уменьшены, чтобы защитить растение своей острой и заостренной формой (рис. 2D). У кактусов фотосинтез происходит в зеленых стеблях. У плотоядных растений скрученные листья действуют как ловушки для насекомых, либо создавая форму длинного кувшина, содержащего ферменты или бактерии, переваривающие насекомых, либо быстро захлопываясь вокруг насекомого. Листья плотоядных растений обычно ярко окрашены для привлечения насекомых (рис. 2В, С).
- Рисунок 2 – Примеры модифицированных листьев.
- Красные стрелки обозначают листья. (A) Усики растений огурца. (B) Свернутый лист растения-кувшина, модифицированный для ловли насекомых. (C) Крупный план верхней части листа растения-кувшина. (D) Колючки кактуса. (E) Розовые прицветники гортензии, простирающиеся под цветками. (F) Прицветник антуриума, красочный для привлечения опылителей. (G) Ювенильный сложный лист из Amorphophallus konjac , который в зрелом возрасте может достигать 4–5 футов в высоту. (H) Raphia regalis , один из самых крупных известных листьев. Размер листа дан относительно роста человека. (I) Ruscus hypoglossum с цветками на листе.
Эволюция листа
Первыми растениями на суше более 400 миллионов лет назад были мохообразные, к которым относятся мхи, роголистники и печеночники [4]. У этих небольших растений полностью отсутствуют сосудистые ткани, у них нет ни листьев, ни корней. Например, если вы очень внимательно посмотрите на мох, вы можете заметить маленькие, зеленые, похожие на листья образования; однако это не настоящие листья, поскольку в них отсутствуют сосудистые ткани. Настоящие листья впервые возникли у сосудистых растений, к которым относятся плауновидные, папоротники и семенные растения. Существует несколько гипотез о том, как появились листья. Возможно, они произошли от ветвей растений, которые стали плоскими и сросшимися, что привело к структуре, которую мы теперь знаем как лист; или листья могли развиться из репродуктивных структур растений [4]. В настоящее время принято считать, что листья развились независимо в нескольких семействах растений. Изучая окаменелости и ДНК, биологи-эволюционисты до сих пор выясняют, сколько раз листья развивались независимо друг от друга.
Отличительные листья
Некоторые растения имеют очень уникальную структуру листьев [5]. Например, у некоторых растений есть только один единственный лист. Amorphophallus konjac ежегодно развивает один сложный лист, черешок которого уходит в землю, так что лист выглядит как небольшое дерево (рис. 2G). Еще один характерный лист относится к Raphia regalis , одному из самых длинных листьев в мире, длина одного листа которого составляет 28 м (91 фут) (рис. 2H) [5]. Представьте себе лист примерно такой же длины, как баскетбольная площадка! Еще один уникальный лист из Ruscus hypoglossum , цветки которого расположены на вершине листовой пластинки (рис. 2I).
Давайте наблюдать вместе!
Теперь вы знаете, что структура и функции листьев очень разнообразны, что способствует поддержанию жизни и здоровья растений: листья бывают разных форм и размеров, и их роль выходит за рамки простого фотосинтеза (рис. 1, 2). Это всегда прекрасное время, чтобы выйти на улицу и собрать листья, даже зимой! Мы создали бинго из листьев, чтобы вы могли использовать его, когда в следующий раз выйдете на улицу (рис. 3). Сколько различных видов листьев вы можете найти? Веселиться!
- Рисунок 3. Листовое бинго, показывающее разнообразие листьев.
- Когда в следующий раз вы проведете какое-то время на улице, найдите листья, соответствующие этим описаниям, и вычеркните их на этой карточке. Когда вы заполнили пять строк подряд, столбец, диагональ или даже всю карточку, вы можете крикнуть «БИНГО!»
Глоссарий
Черешок : ↑ Стебель, соединяющий листовую пластинку со стеблем или меристемой растения.
База : ↑ Область листовой пластинки, которая соединяется с черешком.
Лезвие : ↑ Широкая часть листа, которую также иногда называют листовой пластинкой.
Хлоропласт : ↑ Органелла растительной клетки, являющаяся местом фотосинтеза.
Хлорофилл : ↑ Пигмент, поглощающий и улавливающий энергию солнца.
Устьица : ↑ Поры на поверхности растительных тканей (например, листьев), обеспечивающие газообмен с окружающей средой.
Сосудистые ткани : ↑ Ткани растений, транспортирующие воду вверх и вниз по телу растения.
Каротиноид : ↑ Пигменты, поглощающие свет и отражающие цвет.
Автотроф : ↑ Организм, производящий себе пищу (« авто » = сам, « троф » = пища).
Фотосинтез : ↑ Процесс, при котором энергия солнца, углекислого газа и воды преобразуется в пищу для растений.
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Прежде всего, мы хотели бы поблагодарить молодых рецензентов за их комментарии и полезные отзывы, которые вместе с комментариями редактора помогли нам улучшить окончательную версию этой статьи. Авторы хотели бы поблагодарить учащихся государственных школ Нью-Йорка Обри Хикерсона (10 лет), Анук Роап (11 лет), Майю Палм Сидло (12 лет) и Сантьяго Тобона (9 лет).) за внимательное рассмотрение рукописи и полезные комментарии перед отправкой. Наконец, мы благодарим Джейкоба Суисса (Дендрарий Арнольда Гарвардского университета) за его комментарии и правки в окончательной версии этой рукописи.
Каталожные номера
[1] ↑ Schoonderwoerd, KM, and Friedman, WE 2021. Морфология обнаженных покоящихся почек и их таксономическое и географическое распространение в древесных флорах умеренного пояса. Новый фитолог . 232: 523–36. doi: 10.1111/nph.17506
[2] ↑ Матиле, П. 1992. « Старение хлоропластов », в Фотосинтезе сельскохозяйственных культур: пространственные и временные детерминанты (Амстердам: Elsevier Science). п. 413–40.
[3] ↑ Turlings, CJT, and Erb, M. 2018. Тритрофические взаимодействия, опосредованные травоядными летучими веществами растений: механизмы, экологическая значимость и потенциал применения. год. Преподобный Энтомол . 63:433–52. doi: 10.1146/annurev-ento-020117-043507
[4] ↑ Кенрик П. и Крейн П. Р. 1997. Происхождение и ранняя диверсификация наземных растений . Вашингтон, округ Колумбия: Издательство Смитсоновского института.
[5] ↑ Халле, Ф. 2018. Атлас поэтической ботаники . Кембридж, Массачусетс: MIT Press.
Структура листа и гистохимический анализ семян предоставили структурную информацию о повышении урожайности и качества семян древовидного пиона в условиях легкого затенения
Abstract
В биологии структура является основой функционирования. Для растений изменения их физиологических и экологических функций обычно вызываются структурными изменениями. Чтобы понять, как условия затенения изменяют структуру растения, тем самым предоставляя структурное представление о повышении урожайности и качества, масличный пион был затенен полиэтиленовыми сетками различной плотности с 28 дней после опыления (DAP) до сбора урожая. Толщина листа (LT), жилки (VT), верхнего эпидермиса (UET), нижнего эпидермиса (LET), палисадной ткани (PT), губчатой ткани (ST), а также накопление и распределение крахмала, белка, и жира, наблюдали с 14-дневными интервалами. Результаты показали, что затенение оказало значительное влияние на анатомическую структуру листьев. В период быстрого роста (до 70 DAP) LT, ET и VT при затенении были значительно ниже, чем при отсутствии затенения. В этот период накопление крахмала и белка при затенении было ниже, чем при незатенении. В период созревания (99–112 DAP), LT и PT при затенении были выше, чем при отсутствии затенения, что указывает на то, что легкое затенение замедляет старение листьев и увеличивает фотосинтетическую способность. Затенение задерживает деградацию клеток покровов и продлевает развитие семян и накопление питательных веществ.
Введение
Пион древовидный ( Paeonia , секция Мутань, округ Колумбия) произрастает в Китае, и его дикая зародышевая плазма в основном распространена в горных кустарниках и вторичных широколиственных лесах в провинциях Хэнань, Ганьсу и Шаньси 1 . В процессе пересадки пиона масличного древовидного из горных кустарников в поле их листья обычно желтеют, преждевременно стареют или даже опадают из-за высокой освещенности и температуры, что серьезно снижает урожай и качество семян пиона древовидного 2 . Затенение является предпочтительным методом обеспечения подходящей среды в период роста, поскольку высокая интенсивность света и высокая температура вредны для роста декоративных кустарников 3 . Например, при затенении растений киви значительно оптимизировался микроклимат кроны, снижалась температура и повышалась влажность, что эффективно снижало физиологическое напряжение листьев и плодов и уменьшало количество опавших плодов 4 . В условиях затенения Trollius chinensis Bunge рос относительно лучше и демонстрировал более продолжительный срок наблюдения 5 .
Свет является основным фактором физической среды. Затенение изменяет микроокружение, что, в свою очередь, изменяет физиологические характеристики листа 6 а также конструкция листа 7,8,9 . Затенение увеличивает площадь листьев и уменьшает их толщину 7 , а также уменьшает плотность палисадной ткани и плотность устьиц 8 . Также увеличивается межклеточное пространство между палисадной тканью и губчатой тканью, а также количество сосудистых пучков 5 . Также было обнаружено, что затенение уменьшает толщину палисадной ткани, плотность устьиц и соотношение палисадной ткани к ткани губки 10 . Затенение также влияет на распределение веществ в тканях растений. Распределение крахмальных зерен в основных тканях при 50% затенении было больше, чем при незатенении 11 . Несмотря на эту информацию, влияние затенения на распределение питательных веществ в семенах древовидного пиона остается неясным.
В нашем предыдущем исследовании мы обнаружили, что легкое затенение значительно повышает урожайность и качество семян древовидного пиона 2 . Впоследствии мы исследовали физиологический механизм, посредством которого легкое затенение увеличивает урожай и качество семян посредством анализа процесса развития семян, накопления питательных веществ и физиологических характеристик листьев .6,12 . Структура является основой функции и отвечает за разнообразие физиологии и экологии растений. В этой работе нашими задачами было определить: (i) анатомическое строение листа; (ii) влияние затенения на анатомическую структуру листа; и (iii) гистохимия семян древовидного пиона. Это исследование должно дать теоретическую морфологическую и цитологическую основу для выращивания пиона масличного дерева в условиях легкого затенения.
Материалы и методы
Затенение и экспериментальный план
Экспериментальные схемы затенения были такими же, как описано ранее 2,12 . Полевой эксперимент был проведен на базе посадки масличного пиона на экспериментальной станции Шаньдунского исследовательского центра хлопка, Линьцин (36 °61′ с.ш., 115 °42′ в.д., 35 м над уровнем моря), Шаньдун, Китай, в 2018 г. Два- Для этого исследования были отобраны саженцы Paeonia ostii ‘Feng Dan’, пересаженные в 2015 г. в возрасте 90 лет. Длина ряда × расстояние между рядами составляла 30 см × 50 см. Средняя высота растения до затенения 52,3 см. Участок, на котором последовательно рос пион масличный древовидный, был покрыт полиэтиленовой сеткой трех различных плотностей. Плотность потока фотосинтетических фотонов (PPFD) через три сетки снизилась на 18,4%, 58,6% и 82,1% соответственно при измерении с помощью фотометрической системы Li-1400 (Li-Cor, США), что соответствует трем обработкам легкое, умеренное и сильное затенение 2,6,12 . Размер каждой полиэтиленовой сетки составлял 8 м × 4 м. Обработки были организованы в виде рандомизированных полных блоков с 3 повторениями. Затенение начинали через 28 дней после опыления (DAP) и заканчивали, когда семена были готовы к сбору.
Анатомические анализы
Листья и семена собирали, начиная с 42 DAP с 14-дневными интервалами до сбора семян. Собирали 3-й полностью разросшийся молодой лист на главном стебле от верхушки. Для изучения анатомического строения клеток мезофилла образец листа разрезали на 5-мм сегменты по средней жилке и фиксировали в ФУК (формалин: спирт: ледяная уксусная кислота 90:5:5, В). Затем фиксированный материал обезвоживали с помощью серии этанола, очищали в ксилоле, заливали в парафин, а затем разрезали на срезы толщиной 4–6 мкм с помощью ротационного микротома (RM2016, Leica, Германия). Срезы окрашивали как сафранином, так и быстрым зеленым и фиксировали в нейтральном балате. В дальнейшем их наблюдали и фотографировали с помощью микроскопа (Nikon Eclipse E100, цифровой прицел Nikon DS-U3, Япония).
Семена пиона древовидного фиксировали в FAA и 2,5% глутаральдегиде соответственно. Срезы, фиксированные в FAA, окрашивали PAS (Periodic Acid Schiff) и нафтолсульфоновой кислотой (Servicebio, Ухань, Китай). Гранулы крахмала окрашивались в пурпурно-красный цвет, клеточная стенка также окрашивалась в пурпурно-красный цвет, а белок имел желтый цвет. Семена, фиксированные в 2,5% глутаровом альдегиде, погружали в 1% лимонную кислоту, а затем заливали в парафиновые срезы. Срезы окрашивали нильским красным (Servicebio, Ухань, Китай) в темноте, и под флуоресцентным микроскопом липиды выглядели оранжево-красными или красными.
Сбор и анализ данных
Изображения анализировали с помощью программы Image-Pro Plus 6.0 (Media Cybernetics, США). Отбирали срезы с целыми клетками и измеряли следующие показатели: толщина листа (LT), толщина жилок (VT), толщина верхнего эпидермиса (UET), толщина нижнего эпидермиса (LET), толщина палисадной ткани (PT), толщина губчатой ткани ( ST) и отношение толщины палисадной ткани к толщине губчатой ткани (P/S). Все показатели были собраны по 30 данных и подвергнуты дисперсионному анализу (ANOVA) с использованием программного обеспечения SPSS (SPSS 19)..0, Армонк, штат Нью-Йорк, США).
Результаты
Морфология листьев пиона масличного
Пион масличный обладает двусторонним листом с отчетливой вентральной и абаксиальной поверхностью (рис. 1а). Структура листа снаружи внутрь включает эпидермис, мезофилл и жилки (рис. 1б). Эпидермис является наружным слоем листа и делится на верхний эпидермис и нижний эпидермис, каждый из которых состоит из одной живой клетки. Это основная защитная ткань (рис. 1с). Клетки квадратные и прямоугольные, без хлоропластов, аккуратно расположены, с толстой клеточной стенкой и равномерно покрыты кутикулой. В верхнем эпидермисе нет устьиц или явных дополнительных структур, таких как трихомы или щетинки. Устьица расположены в нижнем эпидермисе.
Рисунок 1 Структура листа пиона масличного. ( a ) Двусторонний лист с вентральной и абаксиальной поверхностью; ( b ) Поперечное сечение листа; ( c ) Структура листа без затенения при 70 DAP; ( d ) Структура листа при легком затенении при 70 DAP; ( e ) Поперечное сечение листа без затенения при 112 DAP; ( f ) Поперечное сечение листа при легком затенении при 112 DAP. ВС: вентральная поверхность; AS: абаксиальная поверхность; UE: верхний эпидермис; PT: палисадная ткань; ST: губчатая ткань; Ер: эпидермис; Я: мезофилл; Ve: вена; Cu: кутикула; LE: нижний эпидермис; МВ: основная вена; ЛЖ: латеральная вена. Шкала баров: ( и ), 1 см; (b) , 100 мкм; ( с, d ), 50 мкм; ( e,f ), 100 мкм.
Изображение полного размера
Мезофилл располагается между верхним и нижним эпидермисом и соединяется с верхним эпидермисом, а длинная ось клетки мезофилла перпендикулярна эпидермису (рис. 1в,г). Палисадная ткань состоит из плотной столбчатой паренхимы, содержащей много хлоропластов. Рыхлые клетки паренхимы между палисадной тканью и нижним эпидермисом представляют собой губчатую ткань. Клетки губчатой ткани имеют неправильный размер и признаки, с короткими рукавообразными выступами и соединены между собой (рис. 1c, d). Хорошо развитое интерстициальное пространство в губчатой ткани и подкамера образуют вентиляционную систему листа. В клетках губчатой ткани меньше хлоропластов.
Ретикулярные вены состоят из главных вен и боковых вен и распределены в ткани мезофилла (рис. 1б). Главная жилка содержит сосудистый пучок, при этом ксилема расположена вверху (вентральная поверхность) и флоэма внизу (абаксиальная поверхность). Между ними находится камбий, при этом его расщепляющая способность была слабой, а время активности коротким. Таким образом, основной сосудистый пучок развит недостаточно. Несколько слоев клеток над сосудистым пучком дифференцируются в ткань колленхимы (рис. 1b), а два самых внешних слоя клеток ниже дифференцируются в ткань склеренхимы, которая усиливает механическую поддержку.
Влияние затенения на клетки эпидермиса
Затенение значительно повлияло на UET (рис. 2a). UET при легком, умеренном и сильном затенении снижалась на 10,0, 16,3 и 19,6% при 42 днях пребывания в день соответственно и на 7,0, 14,2 и 21,8% при 56 днях пребывания в воздухе соответственно. Однако легкое затенение увеличивало UET на 12,0% при 70 DAP, 12,4% при 84 DAP, 13,4% при 98 DAP и 27,2% при 112 DAP. Чтобы исследовать, как различные условия затенения влияют на структуру верхнего эпидермиса, мы измеряли его толщину на разных стадиях роста. UET сначала увеличился, а затем уменьшился, и между видами лечения были значительные различия. UET достигал максимума при 56 DAP при отсутствии затенения, при 84 DAP при легком и умеренном затенении и при 70 DAP при сильном затенении.
Влияние затенения и времени сбора на UET, LET, PT, ST, P/S, VT и LT. ( a ) Влияние затенения и времени на UET; ( b ) Влияние затенения и времени на LET; ( c ) Влияние затенения и времени на PT; ( d ) Влияние затенения и времени на ST; ( e ) Влияние затенения и времени на P/S; ( f ) Влияние затенения и времени на VT; ( г ) Влияние затенения и времени на LT.
Полноразмерное изображение
Затенение оказало значительное влияние на LET (рис. 2b). ЛПЭ при слабом, умеренном и сильном затенении снижалась на 12,5, 19,0 и 20,2% при 56 ДАП и на 10,1, 15,3 и 18,8% при 70 ДАП. Тем не менее, легкое затенение увеличило ЛПЭ на 25,1% при 112 DAP. Подобно UET, изменение LET во времени было параболическим. LET при незатенении достигала максимума при 70 DAP, а затем постепенно снижалась до 112 DAP. Пик ЛПЭ при всех затенениях достигал 84 DAP, в то время как минимум наблюдался при 42 DAP при легком и умеренном затенении и при 112 DAP при сильном затенении.
Влияние затенения на клетки мезофилла
Затенение значительно повлияло на PT (рис. 2c). ПВ при слабом, умеренном и сильном затенении снижалось на 7,3, 14,8 и 17,1% при 42 днях и на 12,0, 14,4 и 17,0% при 56 днях. Напротив, легкое затенение увеличивало PT на 5,1% при 84 DAP и на 7,3% при 98 DAP. PT всех видов лечения сначала увеличивался, а затем уменьшался, при этом наблюдались значительные различия между вариантами лечения. PT при незатенении достигало максимума при 56 DAP, а затем постепенно снижалось до минимума при 112 DAP. При любом затенении PT достигал максимума при 84 DAP и минимума при 42 DAP.
Затенение оказало значительное влияние на ST (рис. 2d). ST при легком, умеренном и сильном затенении снизилась на 5,0, 5,9 и 7,9% при 56 ДПД соответственно и на 4,9, 7,9 и 11,9% при 70 ДПД соответственно. ST при легком затенении был выше, чем при незатенении при 98 и 112 DAP, хотя различия не были достоверными. Тенденция ST была параболической, при этом ST всех видов лечения увеличивалась от минимума в 42 DAP до максимума в 84 DAP, а затем постепенно снижалась.
Затенение значительно повлияло на P/S (рис. 2e). P/S при легком, умеренном и сильном затенении снизилось на 7,3, 9,0 и 9,8% соответственно при 56 ДАП. Светлое затенение увеличивало P/S на 9,0% при 84 DAP, 10,3% при 98 DAP и 10,9% при 112 DAP. P/S всех видов лечения постепенно снижался со временем.
Влияние затенения на толщину жилок и листа
Затенение оказало значительное влияние на VT (рис. 2f). ЖТ при легком, умеренном и сильном затенении снижалась на 5,6, 10,7 и 13,7% при 56 ДАП соответственно и на 5,7, 11,6 и 17,0% при 70 ДАП соответственно. VT при легком затенении был выше, чем при незатенении при 98 и 112 ДАП, но различия не были достоверными. Разница в VT в разное время сбора была значительной. VT при отсутствии затенения достигал максимума при 70 DAP, тогда как при затенении – при 84 DAP.
Затенение значительно повлияло на LT (рис. 1c,f; 2g). LT при световой обработке снизилась на 7,6% при 56 DAP. LT при умеренном затенении снижалась на 10,8% при 56 DAP, 14,2% при 70 DAP и 9,9% при 84 DAP. Сильное затенение снижало LT на 9,0% при 42 DAP, 17,6% при 56 DAP, 18,9% при 70 днях, 13,9% при 84 днях, 10,2% при 98 днях и 9,07% при 112 днях. Между разными временными точками была значительная разница в LT. LT всех вариантов сначала увеличивалась, а затем уменьшалась, тогда как максимум при незатенении и затенении достигался при 70 DAP и 84 DAP соответственно.
Влияние затенения на гистохимию семян
После опыления и осыпания лепестков количество семян, обернутых фолликулами, постепенно увеличивалось (рис. 3а). Семя древовидного пиона состоит из наружного покрова, внутреннего покрова, эндосперма и зародыша снаружи внутрь (рис. 3b,c). И эндосперм, и зародыш проходят стадии свободного ядра и целлюляризации. Эндосперм является основной тканью для хранения питательных веществ и появляется в жидком состоянии на стадии свободного ядра. Объем эндосперма увеличивался по мере увеличения количества свободных ядер. Процесс целлюляризации эндосперма продвигался снаружи внутрь, и самый внутренний слой клеток оставался безъядерным до тех пор, пока эндосперм не был полностью клеточным (рис. 3d). Внутренний покров постепенно деградировал и поглощался клетками эндосперма, наружный покров также деградировал изнутри наружу. Накопление питательных веществ в эндосперме происходило снаружи внутрь, при этом сначала накапливался крахмал, а затем постепенно увеличивалось количество белков и жиров.
Гистохимия семян пиона. ( a ) Семена, завернутые в фолликулы после опадения лепестков; ( b ) Вертикальный срез семян без затенения при 70 DAP; ( c ) Поперечное сечение семян без затенения при 84 DAP; ( d ) Процесс целлюляризации эндосперма продвигался снаружи внутрь; ( e ) Срез семян без затенения при 70 DAP; ( f ) Семена поперечного среза с сильным затенением при 70 DAP. Fo: фолликулы; П: лепестки; En: эндосперм; Эм: эмбрион; OI: наружные покровы; II: внутренние покровы; Ка: полость. Шкала баров: ( а–б ), 1 см; (d ), 200 мкм; ( c,e,f ), 1 мм.
Изображение в натуральную величину
Наблюдалась очевидная разница в реакции окрашивания семян между незатенением и сильным затенением при 70 DAP. Поперечный срез эндосперма без затенения был окрашен в темно-красный цвет с помощью PAS и в желтый цвет с помощью нафтолсульфоновой кислоты (рис. 3e). Напротив, поперечное сечение при сильном затенении было окрашено в розовый и светло-желтый цвета (рис. 3f). Это указывало на то, что эндосперм при незатенении содержал больше крахмала и белка по сравнению с эндоспермом при сильном затенении. Красные сферические капли липидов, окрашенные Нильским красным, на срезе эндосперма при отсутствии затенения были более многочисленными, чем при сильном затенении (рис. 4а, б), что указывает на то, что накопление жира при отсутствии затенения было больше, чем при сильном затенении. . В период созревания красных сферических липидных капель при легком затенении было больше, чем при незатенении (рис. 4c, d). Это указывало на то, что при легком затенении в эндосперме было больше жира, чем при отсутствии затенения. Покровы при отсутствии затенения были значительно истончены и дегенерированы (рис. 4е), а при легком затенении оставались толстыми (рис. 4д) и, таким образом, могли продолжать накапливать питательные вещества.
Липидные капли и покровы семян пиона. ( a ) Капли липидов в эндосперме при незатенении при 70 DAP; ( b ) Капли липидов в эндосперме при сильном затенении при 70 DAP; ( c ) Капли липидов в эндосперме при отсутствии затенения при 112 DAP; ( d ) Капли липидов в эндосперме при легком затенении при 112 DAP; ( e ) Более толстый покров при легком затенении на 112 DAP; ( f ) Более тонкий покров при незатенении при 112 DAP. Ne: ядро; ЛД: липидные капли; CW: клеточная стенка; En: эндосперм; Я: покровы. Шкала баров: ( а–д ), 100 мкм; (e,f ), 50 мкм.
Изображение в полный размер
Обсуждение
PT при легком затенении было выше, чем при незатенении, что может улучшить фотосинтез листьев в период созревания
Анатомия растений связывает анатомические структуры с их функциями и их взаимодействием с окружающей средой, и позволяет прояснить изменения в физиологии и функциях растений с микрокосмической точки зрения 13,14,15,16 . Лист является основным органом ассимиляции и транспирации, а также наиболее чувствительным к стрессу органом. Таким образом, большое значение имеет изучение стрессоустойчивости растений путем изучения анатомического строения листа 9 .0489 17,18 . Исследования анатомической структуры листьев в основном сосредоточены на эпидермисе, мезофилле и жилках 19,20 . Некоторые исследования показали, что структура листа меняется в зависимости от окружающей среды, при этом интенсивность света играет важную роль в росте и морфологической структуре растений 21 .
Свет оказывает важное влияние на фотосинтез, но также действует как сигнал окружающей среды для растений, регулирующий рост, развитие, физиологию, морфологическую структуру и анатомическую структуру для адаптации к различным условиям окружающей среды 22,23,24 . Например, листья Chrysanthemum morifolium были тоньше при освещении 25% и 15% и имели рыхлую палисадную ткань и неравномерно расположенные губчатые клетки мезофилла 25 . Регулировка морфологии и анатомической структуры листа позволяет полностью поглощать свет и улучшать скорость фотосинтеза, тем самым способствуя росту и развитию и, в конечном итоге, влияя на размножение.
В нашей предыдущей работе развитие семян было разделено на три периода следующим образом: быстрый рост (до 70 ДАП), медленный рост (71–98 DAP) и созревание (99–112 DAP) 12 . В этом исследовании LT, VT, PT и ST снижались с уменьшением освещенности на стадии быстрого роста, что согласуется с результатами Camellia 3 и Lycoris radiata var. радиата 23 . Тонкий лист улучшил способность улавливания и использования света, а также эффективность фотосинтеза 26 . LT и PT при затенении были выше, чем при незатенении на стадии созревания (рис. 1д, е). На этой стадии растение явно стареет при отсутствии затенения, демонстрируя пожелтение и высыхание листьев, потерю воды клетками мезофилла и уменьшение объема клеток, в то время как лист при легком затенении оставался зеленым и в хорошем состоянии. Аналогичные результаты были получены в Brassica napus L. 27 . Кроме того, клетки палисадной ткани, как основная площадка для фотосинтеза, имеют прямое отношение к фотосинтетической способности 28 . Тот факт, что PT при легком затенении был выше, чем при незатенении, обеспечивает анатомическую основу для улучшения фотосинтетической способности при легком затенении, чем при отсутствии затенения.
Светлое затенение замедляет деградацию клеток наружного покрова и продлевает развитие семян и накопление питательных веществ
В гистохимии используются специальные химические реагенты на растительных материалах для наблюдения и определения состава или распределения вещества под микроскопом. Он объединяет клеточную морфологию, химический состав и функцию 29 . Эндосперм семян древовидного пиона является основной тканью для хранения питательных веществ и производства масла. Развитие эндосперма начинается с оплодотворенного первичного ядра эндосперма. «Фэн Дан» имеет ядерный эндосперм и проходит стадию свободного ядра и целлюляризации, что согласуется с наблюдением Донг 9.0489 30 . Процесс целлюляризации эндосперма шел снаружи внутрь, следовательно, крахмал, белок и жир также накапливались снаружи внутрь.
Динамические изменения накопления продуктов хранения семян, таких как растворимый сахар, крахмал, растворимый белок, сырое масло и ненасыщенные жирные кислоты, наблюдались в нашем предыдущем исследовании 12 . Мы определили содержание крахмала, белка и сырого жира в тех же образцах, которые использовались для гистохимического анализа, и результаты соответствовали нашим предыдущим наблюдениям. В нашем предыдущем исследовании мы обнаружили, что содержание питательных веществ при затенении было меньше, чем при незатенении на стадии быстрого роста, тогда как содержание жира при легком затенении было выше, чем при незатенении на стадии созревания 12 . Тот же результат мы наблюдали на окрашенном срезе эндосперма.
На стадии свободного ядра покровы были примерно в два раза меньше размера семени, после чего внутренние покровы деградировали по мере увеличения числа свободных ядер и эндосперма. На стадии целлюляризации покровы составляли около трети размера семени, а внутренний и внешний покровы подвергались дальнейшей деградации, пока внешний покров не затвердел, а внутренний покров не сжался в тонкий слой плотных клеток. Когда семя созрело, основная деятельность эндосперма заключалась в преобразовании питательных веществ, а не в накоплении питательных веществ. Дегенерация покровов в затенении задерживалась, а развитие семян удлинялось, что продлевало накопление питательных веществ в эндосперме.
Выводы
Наше исследование показывает, что затенение оказывает значительное влияние на анатомическую структуру листа пиона масличного дерева. На стадии быстрого роста (до 70 DAP) LT при затенении была ниже, чем при отсутствии затенения, что уменьшало потери света и увеличивало скорость фотосинтеза. На стадии созревания (99–112 DAP) LT и PT при легком затенении были выше, чем при отсутствии затенения, что свидетельствовало о замедлении старения листьев и повышении фотосинтетической способности. На стадии быстрого роста накопление крахмала и белка при затенении было меньше, чем при незатенении, а накопление жира при легком затенении было больше, чем при незатенении на стадии созревания. Затенение задерживает дегенерацию покровов и продлевает развитие семян и накопление питательных веществ. Таким образом, был сделан вывод, что структура листа и гистохимический анализ семян позволили получить структурное представление о повышении урожайности и качества семян древовидного пиона в условиях легкого затенения.
Ссылки
Хонг, Д.Ю. и Пан, К.Ю. Таксономическая история и ревизия секты Paeonia. Мутан (Paeoniaceae). Acta Phytotaxono. Грех . 37 , 351–368 (на китайском языке) (1999).
Фини А., Феррини Ф., Франги П., Аморозо Г. и Джордано К. Рост, газообмен листьев и строение листьев трех декоративных кустарников, выращенных при разной интенсивности освещения. евро. Дж. Хортик. науч. 75 , 111–117 (2010).
КАС Google ученый
Хе, К. Дж., Ван, З. Ю. и Ван, Р. К. Влияние затенения летом на экологические факторы и фотосинтез фруктового сада куви. J. Fruit Sci . 24 , 616–619 (на китайском языке) (2007 г.).
Lv, JH, и др. . Влияние затенения на рост, развитие и физиологические характеристики Trollius chinensis Bunge.
Науч. Агр. Грех . 46 , 1772–1780 (на китайском языке) (2013 г.).Han, CJ et al Влияние затенения на старение листьев и физиологические характеристики масличного пиона. J. Nucl. Агр. Наука . 33 , 2492–2498 (на китайском языке, 2019 г.).
Сантос, С.А. и др. . Влияние затенения на морфоанатомию листа и толерантность к глифосату у Commelina benghalensis L. и Cyperus rotundus L. Aust. J. Crop Sci. 9 , 135–142 (2015).
Google ученый
Wijeratne, T.L., Mohotti, AJ & Nissanka, S.P. Влияние длительного затенения на физиологические, анатомические и биохимические изменения чая (Camellia sinensis (L.) O. Kuntz). троп. Агр. Рез. 20 , 376–387 (2008).
Google ученый
Пирес М.В., Алмейда А.
Ф., Абреу П.П. и Сильва Д.К. Объясняет ли затенение вариации морфофизиологических признаков тропических эпифитных орхидей, выращенных в искусственных условиях. Acta Physiol. Завод. 34 , 2155–2164 (2012).Артикул Google ученый
Го, Ю. Х., Цай, З. К., Цао, К. Ф. и Ван, В. Л. Фотосинтетическая и анатомическая акклиматизация листьев четырех видов деревьев тропических лесов к различным условиям роста освещения. Дж. Ухань Бот. Рез . 22 , 240–244 (на китайском языке) (2004 г.).
Sun, Z. R. и др. . Влияние интенсивности света на морфологическую структуру и содержание полисахаридов в стебле Dendrobium loddigesii Rolfe. Китайская ассоциация китайской медицины, 9-я конференция по оценке китайской медицины, Чжэцзян, Цзянде, Китай (на китайском языке) (2008 г.).
Хан, С. Дж., Ван, К., Чжан, Х.
Б. и Донг, Х. З. Развитие семян и накопление питательных веществ под влиянием легкого затенения у масличного пиона (Paeonia ostii ‘Feng Dan’). Науч. Хортик. 251 , 25–31 (2019).КАС Статья Google ученый
Миральес, Дж., Мартинес-Санчес, Дж. Дж., Франко, Дж. А. и Баньон, С. Рост Rhamnus alaternus в четырех смоделированных условиях тени: морфологические, анатомические и физиологические реакции. Науч. Хортик. 127 , 562–570 (2011).
КАС Статья Google ученый
Толен Д., Бум С. и Чжу С. Г. Мнение: перспективы улучшения фотосинтеза путем изменения анатомии листа. Растениевод. 197 , 92–101 (2012).
КАС Статья Google ученый
Бакли, Т. Н., Джон, Г. П., Скоффони, К. и Сак, Л.
Как анатомия листа влияет на перенос воды за пределы ксилемы? Завод физиол. 168 , 1616–1635 (2015).КАС Статья Google ученый
Бинкс, О. и др. . Ограниченная акклиматизация анатомии листьев к экспериментальной засухе деревьев тропических лесов. Физиол дерева. 36 , 1550–1561 (2016).
ОБЪЯВЛЕНИЯ Статья Google ученый
Цзян, С. Д., Ван, X., Гао, Х. Ю., Ши, Л. и Чоу, В. С. Системная регуляция анатомической структуры листа, фотосинтетических характеристик и устойчивости к яркому свету у сорго. Завод физиол. 155 , 1416–1424 (2011).
КАС Статья Google ученый
Лаутербах, М. и др. . Эволюция анатомии листа в засушливых условиях – тематическое исследование южноафриканских Tetraena и Roepera (Zygophyllaceae).
Мол. Филогенет. Эвол. 97 , 129–144 (2016).Артикул Google ученый
Evans, J. R. Анатомия листа обеспечивает более равный доступ света и CO2 между хлоропластами. Новый фитол. 143 , 93–104 (1999).
Артикул Google ученый
ДосАнжос, Л. и др. . Основные признаки листьев, свидетельствующие о пластичности фотосинтеза у тропических видов деревьев. Деревья. 29 , 247–258 (2015).
КАС Статья Google ученый
Гонсалвес Б. и др. . Структура листьев и функция черешни (Prunus avium L.) сортов с открытой и густой кроной. Науч. Хортик. 116 , 381–387 (2008).
Артикул Google ученый
Чжоу, С. Б., Лю, К., Чжан, Д., Ли, К. Ф. и Чжу, Г. П. Фотосинтетические характеристики Lycoris radiata var. radiata для затенения процедур. Photosynthetica 48 , 241–248 (2010).
Артикул Google ученый
Иванова Л. А., Пьянков В. И. Структурная адаптация мезофилла листа к затенению. Рус. J. Физиол растений. 49 , 419–431 (2002).
КАС Статья Google ученый
Хан, С. и др. . Фотосинтетические реакции Chrysanthemum morifolium на ростовое облучение: морфология, анатомия и ультраструктура хлоропластов.
Фотосинтетика. 55 , 184–192 (2017).КАС Статья Google ученый
Чжан, З. Ю., Дуань, П. Н. и Чен, X. Влияние затенения на рост и характеристики фотосинтеза сеянцев Sorbus folgneri. Дж. Ганьсу Агрик. Университет. 6 , 138–143 (2014).
Google ученый
Брюнель-Мюге, С. и др. . Ограничение освещения задерживает старение листьев озимого рапса (Brassica napus L.). J. Регулятор роста растений. 32 , 506–518 (2013).
КАС Статья Google ученый
Гото, Э. и др. . Палисадная форма клеток влияет на индуцированные светом движения хлоропластов и фотосинтез листьев. Науч. 8 , 1472 (2018).
ОБЪЯВЛЕНИЯ Статья Google ученый
Донг З.Л. Исследования репродуктивной биологии Paeonia ostii ‘Feng dan’, магистерская диссертация. Пекинский университет лесного хозяйства (на китайском языке) (2010 г.).
Хан, Си Джей и др. . Легкое затенение улучшает урожайность и качество семян пиона масличного (Paeonia ostii Feng Dan). Дж. Интегр. Агр. 17 , 1631–1640 (2018).
Артикул Google ученый
Mediavilla, S., Escudero, A. & Heilmeier, H. Внутренняя анатомия листьев и эффективность использования фотосинтетических ресурсов: межвидовые и внутривидовые сравнения. Физиол. дерева. 21 , 251–259 (2001).
КАС Статья Google ученый
Wang, XY. Исследование экстракции и локализации алкалоида галантамина в Aurea. Кандидатская диссертация, Нанкинский университет лесного хозяйства (на китайском языке) (2010 г.).
Загрузить ссылки
Благодарности
Эта работа была поддержана Проектом сельскохозяйственных научных и технологических инноваций Шаньдунской академии сельскохозяйственных наук (CXGC2018E06), Шаньдунским провинциальным фондом естественных наук, Китай (ZR2017BC034), и Шаньдунским провинциальным ключевым исследовательским и Программа развития (2018GNC110032), Taishan Scholars, правительство провинции Шаньдун, Китай (tspd20150213).
Информация об авторе
Авторы и организации
Исследовательский центр хлопка Шаньдунской академии сельскохозяйственных наук, Цзинань, 250100, Шаньдун, КНР
Chenjing Han, Qi Wang & Hezhong Dong 90 Департамент пищевой науки
22 Машиностроение, Шаньдунский сельскохозяйственный и инженерный университет, Цзинань, 250100, Шаньдун, КНРШаньдунский центр лесных ресурсов зародышевой плазмы, Цзинань, 250102, Шаньдун, КНР
Lei Wang, Haiping Yang & Yongjun Zhao
Мэй Ю
Авторы
- Chenjing Han
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Mei Yu
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Qi Wang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Lei Wang
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Haiping Yang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Yongjun Zhao
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Hezhong Dong
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Вклады
Чэньцзин Хан отвечает за реализацию всего эксперимента и написание рукописи, Мэй Ю и Лэй Ван отвечают за сбор образцов, Ци Ван и Хайпин Ян отвечают за управление масличными пионами, профессор Чжао и Дун отвечает за дизайн и стоимость теста.
Авторы, переписывающиеся
Соответствие Юнцзюнь Чжао или Хечжун Донг.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете соответствующую ссылку на оригинальный автор(ы) и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Морфология листьев 40 вечнозеленых и листопадных широколиственных субтропических пород деревьев и связь с функциональными экофизиологическими признаками
. 2015 март; 17(2):373-83.
doi: 10.1111/plb.12250.
Epub 2014 1 декабря.
Вт Кребер 1 , H Heklau, H Bruelheide
принадлежность
- 1 Институт биологии/геоботаники и ботанический сад Университета Мартина Лютера Галле-Виттенберг, Галле, Германия.
- PMID: 25441614
- DOI: 10.1111/плб.12250
W Kröber et al. Растение Биол (Штутт). 2015 март
. 2015 март; 17(2):373-83.
doi: 10.1111/plb.12250.
Epub 2014 1 декабря.
Авторы
В Кребер 1 , Х. Хеклау, Х. Брюльхайде
принадлежность
- 1 Институт биологии/геоботаники и ботанический сад Университета Мартина Лютера Галле-Виттенберг, Галле, Германия.
- PMID: 25441614
- DOI: 10.1111/плб.12250
Абстрактный
Мы исследовали потенциал морфологических и анатомических признаков листьев для прогнозирования ключевых экофизиологических функций субтропических деревьев. Мы спросили, можно ли предсказать экофизиологические параметры устьичной проводимости и уязвимости ксилемы к кавитации на основе микроскопических признаков листьев. Мы исследовали 21 вид лиственных и 19 вечнозеленых субтропических деревьев, используя особей одного возраста и из одной и той же среды в эксперименте «Биоразнообразие-экосистемное функционирование» в Цзянси (BEF-Китай). Выбор теоретико-информационной линейной модели использовался для определения наилучшей комбинации морфологических и анатомических предикторов экофизиологических функций. Анатомия и морфология листа сильно зависят от формы листа. Вечнозеленые виды, как правило, имеют более толстые листья, более толстый губчатый и палисадный мезофилл, больше палисадных слоев мезофилла и более толстый субэпидермис. Более 50% всех вечнозеленых видов имели листья с многослойной палисадной паренхимой, в то время как только у одного листопадного вида (Koelreuteria bipinnata) она была. Взаимодействие с формой листа также было включено в лучшие мультипредикторные модели устьичной проводимости (gs) и уязвимости ксилемы к кавитации. Кроме того, максимальная gs была положительно связана с логарифмическим отношением частокола к толщине губчатого мезофилла. Дефицит давления пара (vpd) для максимальной gs увеличивался с логарифмическим отношением толщины палисада к толщине губчатого мезофилла у видов, имеющих листья с сосочками. Напротив, максимальная удельная гидравлическая проводимость и давление ксилемы, при которых происходит 50%-ная потеря максимальной удельной гидравлической проводимости ксилемы (Ψ50), лучше всего прогнозировались по форме листа и плотности губчатой паренхимы. Вечнозеленые виды имели более низкие значения Ψ50 и более низкую максимальную гидравлическую проводимость ксилемы. Поскольку гидравлические характеристики листа и древесины отражались в структурных признаках листа, существует большой потенциал для выявления дополнительных связей между морфологическими и анатомическими признаками листа и экофизиологическими реакциями.
Ключевые слова: БЭФ-Китай; экофизиологические признаки; анатомические признаки листа; функциональные признаки листа; субтропики; деревья.
© 2014 Немецкое ботаническое общество и Королевское ботаническое общество Нидерландов.
Похожие статьи
Связывание гидравлической проводимости ксилемы и уязвимости со спектром экономики листа — межвидовое исследование 39вечнозеленые и листопадные широколиственные субтропические породы деревьев.
Кребер В., Чжан С., Эмиг М., Брюлхайде Х. Кребер В. и др. ПЛОС Один. 2014 25 ноября; 9(11):e109211. doi: 10.1371/journal.pone.0109211. Электронная коллекция 2014. ПЛОС Один. 2014. PMID: 25423316 Бесплатная статья ЧВК.
Полуденная устьичная проводимость больше связана с водным статусом ствола, чем листьев субтропических листопадных и вечнозеленых широколиственных деревьев.
Zhang YJ, Meinzer FC, Qi JH, Goldstein G, Cao KF.
Чжан Ю.Дж. и др.
Окружающая среда растительной клетки. 2013 янв; 36(1):149-58. doi: 10.1111/j.1365-3040.2012.02563.x. Epub 2012 13 июля.
Окружающая среда растительной клетки. 2013.
PMID: 22715809Анатомия листа обеспечивает координацию гидравлической проводимости листа и проводимости мезофилла с CO 2 у Oryza.
Сюн Д., Флексас Дж., Ю Т., Пэн С., Хуан Дж. Сюн Д. и соавт. Новый Фитол. 2017 Январь; 213 (2): 572-583. дои: 10.1111/nph.14186. Epub 2016 22 сентября. Новый Фитол. 2017. PMID: 27653809
Проводимость мезофилла листа и гидравлическая проводимость листа: введение в их измерение и координацию.
Флексас Дж., Скоффони С., Гаго Дж., Сак Л. Флексас Дж. и соавт. J Опытный бот.
2013 г., октябрь; 64 (13): 3965-81. дои: 10.1093/jxb/ert319.
J Опытный бот. 2013.
PMID: 24123453
Обзор.Кавитация, устьичная проводимость и отмирание листьев у сеянцев двух сосуществующих средиземноморских кустарников во время сильной засухи.
Вилагроса А., Беллот Дж., Вальехо В.Р., Гил-Пелегрин Э. Вилагроса А. и др. J Опытный бот. 2003 сен; 54 (390): 2015-24. дои: 10.1093/jxb/erg221. Epub 2003 28 июля. J Опытный бот. 2003. PMID: 12885857 Обзор.
Посмотреть все похожие статьи
Цитируется
Связи между анатомией листьев и массой листьев в расчете на площадь травянистых видов по склону на субальпийском лугу в восточной части Тибета.
Li X, Zhao X, Tsujii Y, Ma Y, Zhang R, Qian C, Wang Z, Geng F, Jin S.
Ли Х и др.
Эколь Эвол. 2022 2 июня; 12(6):e8973. doi: 10.1002/ece3.8973. электронная коллекция 2022 июль.
Эколь Эвол. 2022.
PMID: 35784019
Бесплатная статья ЧВК.Сравнительная анатомия черешков листьев деревьев и кустарников умеренного пояса: роль размера растений, окружающей среды и филогении.
Филартига А.Л., Климеш А., Альтман Дж., Нобис М.П., Кривелларо А., Швайнгрубер Ф., Долежал Дж. Филартига А.Л. и др. Энн Бот. 2022 13 апр; 129(5): 567-582. doi: 10.1093/aob/mcac014. Энн Бот. 2022. PMID: 35136925
Влияние функционального разнообразия на увеличение продуктивности с возрастом в эксперименте по лесному биоразнообразию.
Бонгерс Ф.Дж., Шмид Б., Брюлхайде Х., Бонгерс Ф., Ли С., фон Охаймб Г.
, Ли И., Ченг А., Ма К., Лю Х.
Бонгерс Ф.Дж. и соавт.
Нат Экол Эвол. 2021 Дек;5(12):1594-1603. doi: 10.1038/s41559-021-01564-3. Epub 2021 4 ноября.
Нат Экол Эвол. 2021.
PMID: 34737435Направленная потеря видов снижает продуктивность сообщества в эксперименте по биоразнообразию субтропических лесов.
Чен Ю, Хуан Ю, Никлаус П.А., Кастро-Исагирре Н., Кларк А.Т., Брюлхайде Х., Ма К., Шмид Б. Чен Ю и др. Нат Экол Эвол. 2020 апр;4(4):550-559. doi: 10.1038/s41559-020-1127-4. Epub 2020 2 марта. Нат Экол Эвол. 2020. PMID: 32123320
Накопленная реакция листопадных Liquidambar formosana Hance и Evergreen Cyclobalanopsis glauca Thunb. Проростки для имитации азотных добавок.
Zhang Z, Zhao Y, Zhang X, Tao S, Fang X, Lin X, Chi Y, Zhou L, Wu C.
Чжан Цзи и др.
Фронт завод науч. 2019 20 декабря; 10:1596. doi: 10.3389/fpls.2019.01596. Электронная коллекция 2019.
Фронт завод науч. 2019.
PMID: 31921245
Бесплатная статья ЧВК.
Просмотреть все статьи “Цитируется по”
Типы публикаций
термины MeSH
Как большие деревья, такие как секвойи, пропускают воду от корней к листьям?
На прошлой неделе мы представили общий план того, как деревья поднимают воду. Дональд Дж. Мерхаут из Monrovia Nursery Company со штаб-квартирой в Азусе, Калифорния, предоставил более подробный ответ:
«Вода часто является наиболее ограничивающим фактором для роста растений. Поэтому растения разработали эффективную систему для поглощения, перемещения, хранения и использования воды. Чтобы понять транспорт воды в растениях, нужно сначала понять устройство растений. Растения содержат обширную сеть каналов, которая состоит из тканей ксилемы и флоэмы.Этот путь транспорта воды и питательных веществ можно сравнить с сосудистой системой, транспортирующей кровь по всему телу человека.Как и сосудистая система у людей, ткани ксилемы и флоэмы распространяются по всему растению.Эти проводящие ткани начинаются в корнях и пересекают стволы деревьев, разветвляясь на ветви, а затем разветвляясь еще дальше на каждый лист.
«Ткань флоэмы состоит из живых удлиненных клеток, соединенных друг с другом. Ткань флоэмы отвечает за транспортировку питательных веществ и сахаров (углеводов), вырабатываемых листьями, в области растения, которые метаболически активны (требующие сахара для энергии и роста). Ксилема также состоит из удлиненных клеток. После образования клеток они умирают. Но клеточные стенки остаются неповрежденными и служат отличным трубопроводом для транспортировки воды от корней к листьям. одно дерево будет иметь много тканей или элементов ксилемы, проходящих через дерево. Каждый типичный сосуд ксилемы может иметь диаметр всего несколько микрон.
“Физиология поглощения и транспорта воды также не столь сложна. Основной движущей силой поглощения и транспорта воды в растение является транспирация воды из листьев. Транспирация – это процесс испарения воды через специальные отверстия в листьях, называемые устьицах. Испарение создает отрицательное давление водяного пара в окружающих клетках листа. Как только это происходит, вода втягивается в лист из сосудистой ткани, ксилемы, чтобы заменить воду, которая испарилась из листа. воды, или напряжение, возникающее в ксилеме листа, будет распространяться вниз через остальную часть ксилемного столба дерева и в ксилему корней благодаря когезионным силам, удерживающим вместе молекулы воды вдоль сторон трубок ксилемы (помните, что ксилема представляет собой непрерывный столб воды, который простирается от листа до корней). увеличение поглощения воды из почвы.
“Теперь, если транспирация из листа уменьшится, как это обычно происходит ночью или в пасмурную погоду, падение давления воды в листе будет не таким большим, и поэтому потребность в воде будет меньше (меньше напряжение) помещенного на ксилему. Потеря воды листом (отрицательное давление воды или вакуум) сравнима с помещением всасывания на конец соломинки. Если созданное таким образом вакуум или всасывание достаточно велико, вода будет подниматься вверх через соломинку .Если бы у вас была соломинка очень большого диаметра, вам потребовалось бы большее всасывание, чтобы поднять воду. Точно так же, если бы у вас была очень узкая соломинка, потребовалось бы меньшее всасывание. Такая корреляция возникает в результате когезионной природы воды вдоль стороны соломинки (стороны ксилемы).Из-за узкого диаметра трубки ксилемы степень натяжения воды (вакуум), необходимая для проталкивания воды через ксилему, может быть легко достигнута за счет нормальной скорости транспирации, которая часто имеет место. в листьях».
Алан Дикман — руководитель учебной программы факультета биологии Орегонского университета в Юджине. Он предлагает следующий ответ на этот часто задаваемый вопрос:
«Оказавшись внутри клеток корня, вода попадает в систему взаимосвязанных клеток, составляющих древесину дерева и простирающихся от корней через стебель и ветви в листья. Научное название древесной ткани — ксилема; состоит из нескольких различных типов клеток. Клетки, которые проводят воду (вместе с растворенными минеральными питательными веществами), длинные и узкие, и перестают быть живыми, когда они выполняют функцию переноса воды. Некоторые из них имеют открытые отверстия наверху и внизу и сложены более или менее подобно бетонным канализационным трубам. Другие клетки сужаются на концах и не имеют полных отверстий. Однако у всех есть ямки в стенках ячеек, через которые может проходить вода. Вода перемещается из одной ячейки в другую, когда есть давление разница между ними.
«Поскольку эти клетки мертвы, они не могут активно участвовать в перекачивании воды. Может показаться возможным, что живые клетки в корнях могут создавать высокое давление в клетках корней, и в ограниченной степени этот процесс действительно происходит. Но общий опыт говорит нам что вода в древесине находится не под положительным давлением, а под отрицательным или всасывающим. Чтобы убедиться в этом, представьте себе, что происходит, когда рубят дерево или когда в стволе просверливают отверстие. если бы давление в штоке было положительным, можно было бы ожидать выхода струи воды, что случается редко.
«На самом деле всасывание, существующее внутри водопроводящих клеток, возникает в результате испарения молекул воды с листьев. Каждая молекула воды имеет как положительно, так и отрицательно электрически заряженные части. В результате молекулы воды имеют тенденцию прилипать друг к другу; благодаря этой адгезии вода образует округлые капли на гладкой поверхности, а не растекается в совершенно плоскую пленку.Когда одна молекула воды испаряется через пору листа, она оказывает небольшое притяжение на соседние молекулы воды, уменьшая давление в листе. проводящие воду клетки листа и выводящие воду из соседних клеток.Эта цепочка молекул воды простирается от листьев до корней и даже уходит от корней в почву.Таким образом, простой ответ на вопрос о том, что продвигает воду от корней к листьям в том, что это делает солнечная энергия: солнечное тепло заставляет воду испаряться, приводя в движение водную цепь».
Обновлено 8 февраля 1999 г.
Хэм Кейлор-Фолкнер — профессор лесного хозяйства Колледжа сэра Сэндфорда Флеминга в Линдси, Онтарио. Вот его объяснение:
Изображение: CHERYL MATTHEWS, Humboldt Redwoods Interpretive Association REDWOOD TREES. Старовозрастные секвойи, такие как эти гиганты из Рокфеллеровского леса в государственном парке Гумбольдт-Редвудс в Калифорнии, достигают высоты 100 метров и более. |
Чтобы превратиться в высокие, самоподдерживающиеся наземные растения, деревья должны были развить способность транспортировать воду из запаса в почве к кроне — расстояние по вертикали, которое в некоторых случаях составляет 100 метров и более ( высота 30-этажного дома). Чтобы понять это эволюционное достижение, необходимо знать структуру древесины, некоторые биологические процессы, происходящие внутри деревьев, и физические свойства воды.
Вода и другие материалы, необходимые для биологической активности деревьев, переносятся по стволу и ветвям в тонких полых трубках в ксилеме или древесной ткани. Эти трубки называются сосудистыми элементами в лиственных или лиственных деревьях (те, которые теряют листья осенью), и трахеидами в хвойных или хвойных деревьях (те, которые сохраняют большую часть своей последней листвы в течение зимы). Элементы сосудов соединяются встык через перфорационные пластины, образуя трубки (называемые сосудами), длина которых варьируется от нескольких сантиметров до многих метров в зависимости от вида. Их диаметр колеблется от 20 до 800 микрон. Вдоль стенок этих сосудов есть очень маленькие отверстия, называемые ямками, которые позволяют перемещать материалы между соседними сосудами.
Трахеиды у хвойных деревьев намного меньше, редко превышая пять миллиметров в длину и 30 микрон в диаметре. У них нет перфорированных концов, поэтому они не соединяются встык с другими трахеидами. В результате ямки у хвойных, также встречающиеся по длине трахеид, приобретают более важную роль. Это единственный способ, с помощью которого вода может перемещаться от одной трахеиды к другой, когда она движется вверх по дереву.
Для движения воды через эти элементы от корней к кроне должен образоваться непрерывный столб. Считается, что эта колонка инициируется, когда дерево является недавно проросшим рассадами, и поддерживается на протяжении всей жизни дерева двумя силами-одна толкает воду из корней, а другая тянет воду к короне. Толчок осуществляется двумя действиями, а именно капиллярным действием (тенденция воды подниматься в тонкой трубе, потому что он обычно течет вдоль стен трубки) и давление корня. Капиллярное действие является второстепенным компонентом толчка. Давление корней обеспечивает большую часть силы, толкающей воду, по крайней мере, на небольшой путь вверх по дереву. Корневое давление создается водой, перемещающейся из его резервуара в почве в корневую ткань путем осмоса (диффузия вдоль градиента концентрации). Этого действия достаточно, чтобы преодолеть гидростатическую силу толщи воды-и осмотический градиент в тех случаях, когда уровень воды в почве низкий.
Капиллярное действие и корневое давление могут поддерживать столб воды высотой от двух до трех метров, но более высокие деревья — фактически все деревья в зрелом возрасте — очевидно, требуют большей силы. У некоторых более старых экземпляров, включая некоторые виды, такие как Sequoia , Pseudotsuga menziesii и многие виды из влажных тропических лесов, крона достигает 100 и более метров над землей! В этом случае дополнительной силой, которая тянет столб воды вверх по сосудам или трахеидам, является эвапотранспирация, потеря воды листьями через отверстия, называемые устьицами, и последующее испарение этой воды. Поскольку вода теряется из клеток листа в результате транспирации, устанавливается градиент, при котором движение воды из клетки увеличивает ее осмотическую концентрацию и, следовательно, давление всасывания. Это давление позволяет этим клеткам высасывать воду из соседних клеток, которые, в свою очередь, берут воду из соседних клеток и так далее — от листьев к веткам, к ветвям, к стеблям и вниз к корням — поддерживая постоянное притяжение.
Изображение: ГЭРИ АНДЕРСОН, Университет Южного Миссисипи ТИПЫ КСИЛЕМ. Некоторые элементы сосудов имеют полную перфорацию ( 1 ), а другие не имеют торцевых стенок ( 2 ). Трахеиды ( 3 ) имеют перекрывающиеся стенки и ямки. |
Для поддержания непрерывного столба молекулы воды также должны иметь сильное сродство друг к другу. Эта идея называется теорией когезии. Действительно, вода обладает огромной когезионной силой. Теоретически это сцепление оценивается в 15 000 атмосфер (атм). Однако экспериментально оказалось, что оно намного меньше и составляет всего 25–30 атм. При атмосферном давлении на уровне земли девяти атм более чем достаточно, чтобы «повесить» водяной столб в узкой трубке (трахеидах или сосудах) с верхушки стометрового дерева. Но требуется большая сила, чтобы преодолеть сопротивление течению и сопротивление поглощению корнями. Тем не менее, многие исследователи продемонстрировали, что силы сцепления воды более чем достаточно для этого, особенно когда этому способствует капиллярное действие внутри трахеид и сосудов.
В заключение, деревья вошли в круговорот, в котором вода циркулирует из почвы в облака и обратно. Они способны удерживать воду в жидкой фазе на всю свою высоту, поддерживая столб воды в небольших полых трубках, используя корневое давление, капиллярное действие и силу сцепления воды.
Марк Витош, ассистент программы в расширенном лесном хозяйстве в Университете штата Айова, добавляет следующую информацию:
Изображение: ТИХООКЕАНСКИЙ ЛЮТЕРАНСКИЙ УНИВЕРСИТЕТ КСИЛЕМ. Вода проходит от корней дерева к его кроне посредством этой проводящей ткани. |
В деревьях происходит множество различных процессов, которые позволяют им расти. Одним из них является движение воды и питательных веществ от корней к листьям в кроне или верхним ветвям. Вода является строительным материалом живых клеток; это питательный и очищающий агент, а также транспортная среда, которая позволяет распределять питательные вещества и соединения углерода (пищу) по всему дереву. Прибрежное красное дерево, или Sequoia sempervirens может достигать высоты более 300 футов (или примерно 91 метр), что является большим расстоянием для перемещения воды, питательных веществ и соединений углерода. Чтобы понять, как вода движется по дереву, мы должны сначала описать ее путь.
Вода и минеральные питательные вещества — так называемый сокодвижение — проходят от корней к верхушке дерева внутри слоя древесины, находящегося под корой. Эта заболонь состоит из проводящей ткани, называемой ксилемой (состоящей из маленьких трубчатых клеток). Существуют большие различия между лиственными (дуб, ясень, клен) и хвойными (секвойя, сосна, ель, пихта) породами в строении ксилемы. В лиственных породах вода движется по дереву в клетках ксилемы, называемых сосудами, которые выстроены в линию конец к концу и имеют большие отверстия на концах. Напротив, ксилема хвойных состоит из закрытых клеток, называемых трахеидами. Эти клетки также выстроены встык, но часть примыкающих к ним стенок имеет отверстия, играющие роль сита. По этой причине вода движется быстрее по более крупным сосудам лиственных пород, чем по более мелким трахеидам хвойных.
Сосудистые и трахеидные клетки позволяют воде и питательным веществам перемещаться вверх по дереву, в то время как специализированные лучевые клетки пропускают воду и пищу горизонтально через ксилему. Все клетки ксилемы, несущие воду, мертвы, поэтому действуют как труба. Ткань ксилемы находится во всех годичных кольцах (древесине) дерева. Не все виды деревьев имеют одинаковое количество годичных колец, участвующих в движении воды и минеральных веществ. Например, хвойные деревья и некоторые лиственные породы могут иметь несколько годичных колец, которые являются активными проводниками, тогда как у других видов, таких как дубы, функционирует только годичный годичный годичный год.
Эта уникальная ситуация возникает из-за того, что ткань ксилемы дуба имеет очень большие сосуды; они могут быстро переносить много воды, но также могут быть легко разрушены из-за замерзания и воздушных карманов. Удивительно, как 200-летний живой дуб может выжить и расти, используя только поддержку очень тонкого слоя ткани под корой. Остальные 199 колец роста в основном неактивны. Однако у прибрежного красного дерева ксилема в основном состоит из трахеид, которые медленно перемещают воду к вершине дерева.
Изображение: УНИВЕРСИТЕТ PURDUE СТОМАТЫ. Эти поры в листьях позволяют воде выходить и испаряться — процесс, который помогает вытягивать больше воды через дерево от его корней. |
Теперь, когда мы описали путь, по которому вода проходит через ксилему, мы можем поговорить о задействованном механизме. Вода обладает двумя характеристиками, которые делают ее уникальной жидкостью. Во-первых, вода прилипает ко многим поверхностям, с которыми соприкасается. Во-вторых, молекулы воды также могут слипаться или держаться друг за друга. Эти две особенности позволяют воде, как резиновой ленте, тянуться вверх по маленьким капиллярным трубкам, таким как клетки ксилемы.
У воды есть энергия для выполнения работы: она переносит химические вещества в растворе, прилипает к поверхностям и делает живые клетки набухшими, заполняя их. Эта энергия называется потенциальной энергией. В состоянии покоя чистая вода обладает 100 % своей потенциальной энергии, которая по соглашению равна нулю. Когда вода начинает двигаться, ее потенциальная энергия для дополнительной работы уменьшается и становится отрицательной. Вода перемещается из областей с наименьшей отрицательной потенциальной энергией в области, где потенциальная энергия более отрицательна. Например, самый отрицательный водный потенциал в дереве обычно находится на границе лист-атмосфера; наименьший отрицательный водный потенциал обнаруживается в почве, где вода поступает в корни дерева. По мере того как вы продвигаетесь вверх по дереву, водный потенциал становится более отрицательным, и эти различия создают притяжение или напряжение, которое поднимает воду вверх по дереву.
Ключевым фактором, который помогает создать притяжение воды вверх по дереву, является потеря воды из листьев в результате процесса, называемого транспирацией.