Схемы по клеточкам сложные: Рисунки по клеточкам «Сложные» ☆ 1000 рисунков
Простые и сложные маленькие картинки по клеточкам. Схемы и фото
Рисование по клеткам — это способ творческого самовыражения и раскрытия талантов.
Научиться рисовать, просто взяв листок, карандаш, фломастер или ручку, не получится — придется потрудиться.
А рисование по клеточкам поможет научиться рисовать быстро и легко
Как научиться рисовать по клеточкам
Чтобы нарисовать по клеточкам, нужно знать правила и принципы рисунка по клеткам.
Это не сложно, и уже после нескольких пробных рисунков вы научитесь рисовать.
Главное — терпение.
Не каждому удалось окончить художественную школу, чтобы научиться технике рисования. Если хотите сделать креативную открытку своими руками или заполнить дневник оригинальными рисунками, освойте рисование по клеточкам. Маленькие картинки по клеточкам смогут сделать даже новички. Главное, купить тетрадку для математики со светлой бумагой.
Как рисовать по клеточкам
Многие любят разгадывать японские кроссворды, в основу которых положено рисование по клеточкам. Если у вас есть готовые разгаданные кроссворды или ответы к ним, то сможете просто перерисовать в свою тетрадку большие фигуры.
Если у вас есть готовые разгаданные кроссворды или ответы к ним, то сможете просто перерисовать в свою тетрадку большие фигуры.
Самый хороший способ использовать готовые схемы, которые были специально разработаны для тех, кто не умеет рисовать. Вы можете закрашивать по схеме клеточки в собственной тетради, а потом удивлять красивыми изображениями близких и родных.
Среди шаблонов вы найдете схемы человеческих лиц
Оригинально смотрятся фрукты по клеточкам. Если хорошо закрасить рисунок яркими фломастерами, то потом можно его вырезать и использовать для декора интерьера или украшения аппликации.
Хотите сделать открытку или описать в своем дневнике романтическую историю, тогда нарисуйте сердечко по клеткам.
Конфетки, букетики, цветочки – все это можно нарисовать по клеточкам.
Если вы освоите принцип, то потом сможете изображать все, что угодно в своей собственной тетради.
Хотите придумать свой собственный рисунок? Тогда сделайте легкую зарисовку, а потом начинайте превращать ее в рисунок по клеточкам. Начинать лучше всего с контура. Потом можете выделять мелкие детали. Не забудьте отметить, каким цветом, какая деталь должна быть выделена, чтобы рисунок получится ярким и красивым.
Начинать лучше всего с контура. Потом можете выделять мелкие детали. Не забудьте отметить, каким цветом, какая деталь должна быть выделена, чтобы рисунок получится ярким и красивым.
3D-рисунки по клеточкам – это хороший способ провести интересно досуг и реализовать свои творческие способности.
Вы еще ни разу не рисовали по клеточкам? Тогда обязательно попробуйте. Это занятие придется по душе как маленьким детям, так и взрослым. Специалисты отметили, что это хобби развивает творческое мышление, координацию движений при письме, концентрацию внимания и логику. Проводите досуг с пользой, выдумывая новые 3Д схемы простые и сложные для рисования по клеточкам.
Сложный рисунок по клеточкам
Предлагаем фото нескольких популярных схем для начинающих
Мороженное по клеточкам
Посмотрите сложные схемы изо по клеточкам и попробуйте нарисовать такие же.
Из мультика Энгри Бердс
Миньоны по клеточкам
Не обязательно с точностью копировать рисунки и их цветовую гамму, фантазируйте, тогда ваша тетрадка в клетку будет заполнена маленькими необычными картинками. Если вырезать картинку и поместить ее в самодельную рамочку для фотографии, то получится креативный подарок.
Если вырезать картинку и поместить ее в самодельную рамочку для фотографии, то получится креативный подарок.
Рубрика: Рисуют дети
Детей бывает сложно удивить, но это не означает, что сделать это невозможно. И после целого дня беготни, прыганья, танцев, игр, каждый должен немного успокоиться и заняться чем-то творческим и развивающим. На помощь и приходят маленькие рисунки по клеточкам. Когда нужно занять малышей – вытяните большой лист бумаги в клеточку, чтобы дети могли рисовать вместе.
Маленькие рисунки по клеточкам, хорошая или плохая идея?
Конечно, маленькие рисунки по клеточкам в блокноте – также хорошая идея, особенно, когда вы находитесь в пути с ребенком и занять его нечем. Маленькие и милые они помогут вашему чаду хорошо провести время, они получат от таких занятий максимум пользы. Маленькие рисунки по клеточкам в тетради — простая художественная деятельность, в которой сочетаются искусство и математика.
Леденцы по клеточкам фото
Картошка фри по клеточкам
Котенок по клеткам фото
Инструменты для рисования маленьких картинок по клеткам
Не говорите детям много, сделайте сюрприз, возьмите бумагу разного типа, маркеры или цветные карандаши и ручки и позвольте детям приступить к рисованию. Рисунки могут быть произвольными, иногда полезно дать возможность ребенку развить фантазию посредствам рисования. Но можно выбирать и конкретные рисунки, легкие для 5 лет.
Рисунки могут быть произвольными, иногда полезно дать возможность ребенку развить фантазию посредствам рисования. Но можно выбирать и конкретные рисунки, легкие для 5 лет.
Если у вас есть домашний принтер – тогда вообще здорово. Вы можете настроить и создать собственную графическую бумагу в специальном приложении. У них есть много вариантов для графической бумаги — обычный квадрат, треугольник, и многое другое. Но на этот шаг решайтесь после того, как дети освоят рисование по клеткам. В приложении все же легко выбрать размер формы, которая вам нужна, толщину, цвет линий и многое другое. Тогда макет просто сохраняется их в формате pdf и вы можете распечатать его сразу же.
Используя обычную бумагу в клеточку, можно сделать простые повторяющиеся рисунки, рисунки шахматной доски. Можно объединить квадраты, чтобы делать большие фигуры и разделять квадраты на треугольники и меньшие квадраты и даже на восьмиугольники, чтобы делать всевозможные интересные изображения.
Треугольники и шестиугольники также хорошо подходят для узоров и картин. Для тех, кто уже хорошо справляется с разными фигурами и отлично ориентируется в основах геометрических форм, можно взять за шаблон смайлики из вк. Позвольте ребенку выбрать любимые смайлики и перерисовать их в тетради. Хорошей идеей являются и животные.
Для тех, кто уже хорошо справляется с разными фигурами и отлично ориентируется в основах геометрических форм, можно взять за шаблон смайлики из вк. Позвольте ребенку выбрать любимые смайлики и перерисовать их в тетради. Хорошей идеей являются и животные.
Рисовать их первый раз может быть не так просто, если использовать клеточки, но на самом деле, дети быстро подхватят эту идею и уже спустя какое-то время смогут воплощать на листе в клеточку самые смелые идеи.
Несмотря на то, что это простая идея, она дает много пространства для творчества, что с большим количеством случайных математических понятий дает большой бонусный плюс для развития ребенка.
Арбуз по клеткам фото
Миньоны по клеткам фото
Супергерои по клеткам
Котик аниме по клеткам
Графический диктант
Стоит отметить, что задания с графической бумагой популярны в детских садиках. Один из распространенных приемов – создание рисунка без образца. Это своеобразный графический диктант. Такое задание легко воспроизвести дома со своим ребенком. Для этого упражнения мы будем использовать листы бумаги формата 4×4. Начиная с левого верхнего угла, мы будем начинать закрашивать квадратики с помощью простых инструкций. Эти инструкции включают:
Для этого упражнения мы будем использовать листы бумаги формата 4×4. Начиная с левого верхнего угла, мы будем начинать закрашивать квадратики с помощью простых инструкций. Эти инструкции включают:
- переместить один квадрат вправо;
- переместить один квадрат влево;
- переместить один квадрат вверх;
- переместить один квадрат вниз. Вот как мы будем писать алгоритм, чтобы проинструктировать ребенка (который будет закрашивать клеточки).
Выберите простой рисунок, такой как шахматная доска, который будет использоваться в качестве примера. Это хороший способ ввести все символы в ключ. Чтобы начать, заполните график для ребенка — квадрат к квадрату — затем попросите его помочь описать, что вы только что сделали. Во-первых, вы можете говорить алгоритм вслух, тогда вы можете превратить свои словесные инструкции в программу. Пример алгоритма: «Переместить вправо, заполнить квадрат, двигаться вправо, сдвигаемся вниз. Заполнить квадрат, переместиться влево, переместиться влево, заполнить квадрат».
Если ребенок хорошо справляется с этим упражнением, то это повод придумать альтернативное задает с похожей сутью, но сложнее. Если есть еще непонимание, сохраните это задание и попробуйте повторить это на следующий день, а пока поработайте с другим примером.
Если ребенок понимает алгоритм и может определить правильные символы для каждого шага, он готов двигаться дальше. В зависимости от вашего ребенка, его возраста и развития вы можете либо попытаться сделать сложную сетку вместе, либо перейти к тому, чтобы ребенок работал в паре с другом. Им понравится играть вместе, давая друг другу такие задания. Это отличный способ заставить ребенка работать творчески, придумывая собственные веселые картинки и разбивая их на алгоритмы передвижения по клеткам и их заполнения.
Маленькие рисунки по клеточкам на фото:
Прошу тебя, проголосуй!
(315 оценок, среднее: 4,18 из 5)
Загрузка…
>
Рисунки по клеткам сложные схемы
Все мы художники в душе. И всем нам хочется свой мир разукрасить. А потому рисунки по клеточкам в тетради могут нам в этом помочь. С ними легко можно выполнить сложные и простые рисунки. Понять, как нарисовать сердце по клеточкам, или же, еду, цветы, игривую маму-кошку и ее забияку котенка. А хотите, у вас могут получиться и портреты? Например, есть такие рисунки по клеточкам, фото которых напоминают и изображения людей: мальчика и девочку, все эти разные рисунки несложно освоить.
И всем нам хочется свой мир разукрасить. А потому рисунки по клеточкам в тетради могут нам в этом помочь. С ними легко можно выполнить сложные и простые рисунки. Понять, как нарисовать сердце по клеточкам, или же, еду, цветы, игривую маму-кошку и ее забияку котенка. А хотите, у вас могут получиться и портреты? Например, есть такие рисунки по клеточкам, фото которых напоминают и изображения людей: мальчика и девочку, все эти разные рисунки несложно освоить.
Чтобы понять, как рисовать по клеточкам цветные красивые картинки, стоит познакомиться с техникой нанесения узора по номерам. Увидеть, что есть разные схемы и все они очень легкие, доступные даже новичкам. Ими можно быстро овладеть. Ведь для каждого из нас по небольшим частям воспроизвести нарисованных зверушек, смайлы и сердечки будет не сложно.
И все же, какие есть маленькие и большие, цветные и черно-белые рисунки, выполненные так, чтобы их легко было повторить; и какие перспективы овладеть этой техникой:
- Какие существенные преимущества имеют рисунки по клеточкам для начинающих?
- Тематические рисунки карандашом по клеточкам;
- Область применения таких оригинальных рисунков;
- Какие возможности дают красивые рисунки по небольшим частям.

Самое важное в знакомстве – увидеть, что это подготовленная на нашем сайте для вас коллекция очень красива. И здесь собраны интересные и легкие рисунки. Среди них есть те, которые высоко оценены нашими гостями и давно им знакомы, а есть и новые, любопытные рисунки по клеточкам для личного дневника.
Каждый может быть художником! Это заявление абсолютно точно гарантирует, что все наши гости, как только узнают, как научиться рисовать по клеточкам, и смогут скачать на сайте пару-тройку вариантов, красиво все повторят и разукрасят. Для каких бы целей ни служили наши подсказки, например, если это – картинки по клеточкам для девочек 12 лет или рисунки с аппетитной едой, все их можно использовать, чтобы отточить свои художественные способности.
Не только образцы готовых открыток у нас есть, но и рисунки по клеточкам: схемы. Такая подсказка, как готовая инструкция поможет двигаться четко по плану, а может быть и в своей, привычной, любимой манере выполнить работу любой сложности. Например, сделать рисунок мороженого по клеточкам, или животных, того же самого котика, или целые композиционные иллюстрации для личного дневника.
Например, сделать рисунок мороженого по клеточкам, или животных, того же самого котика, или целые композиционные иллюстрации для личного дневника.
Не только для давних друзей нашего развлекательного ресурса предоставляется такая возможность, но и новые гости тоже получат шанс обучиться этому искусству, они имеют возможность взять своеобразный мастер класс, урок по изображению всевозможных картинок, на любой вкус и разной сложности.
Картинки на разнообразные темы
Самое привлекательное, что на сайте есть иллюстрации, интересные, как для девочек, так и для мальчиков. А есть нейтральные темы, к примеру, рисунки по клеточкам еда, а так же, иллюстрации по клеточкам животные: домашние любимцы или лесные зверушки, есть и сказочные, такие, как единорог.
Специально, для всех деток, кто любит мультфильм про милых пони и их дружбу, мы подготовили сюрприз! У нас есть картинки по клеточкам пони. Яркие, красочные, они очень привлекательные для деток. А потому мы предлагаем схему, как нарисовать пони по клеточкам. Эта и подобные «инструкции» достаточно понятные и лёгкие даже для ребенка. А главное, они интересные для малышей.
Эта и подобные «инструкции» достаточно понятные и лёгкие даже для ребенка. А главное, они интересные для малышей.
Отдельная категория – это рисунки по клеточкам смайлики. Они всегда интересны и всегда актуальны. Они передают настроение и их просто повторить. Для взрослых и детей такая тема именно то, что может подарить радость от плодотворного труда.
Удивительно, как часто подобные картинки для выручают нас. Благодаря им можно прекрасно провести время с ребеночком, сколько бы ему не было лет, 5,7 или только год. Мы можем в блокноте делать наброски на скучных совещаниях или в дороге занять себя. А картинки по клеточкам для личного дневника – это вообще незаменимая вещь. А потому, везде и при любых случаях скачивайте или сами нарисуете милые иллюстрации.
(18 оценок, среднее: 4,33 из 5)
Дорогие пользователи, а так же гости нашего сайта, сегодня мы с вами рассмотрим технологию рисования рисунки по клеточкам .
Наверное, каждый из нас закрашивал клеточки на полях школьных тетрадей. У кого-то из этого всего получались интересные орнаменты, кто-то писал таким образом тексты, но далеко не всем известна технология рисования
У кого-то из этого всего получались интересные орнаменты, кто-то писал таким образом тексты, но далеко не всем известна технология рисования
Если хотите усовершенствовать свой навык рисования обязательно прочитайте статью рисунки карандашом. Нужен ли особый талант?
Что такое рисунки по клеточкам?
Рисунки по клеточкам это вид изобразительного искусства, в котором используется пиксельная (точечная) графика. В зависимости от сложности такого изображения увеличивается его площадь и количество пикселей (в нашем случае – клеток), которые закрашиваются. Чем больше будет площадь изображения, тем выше будет реалистичность изображения при осмотре с дальнего расстояния.
Давайте рассмотрим один из примеров таких работ:
Как вы сами можете заметить, если смотреть на картинку издали – мы видим чёткое изображение, но если приблизиться – наблюдаем отдельные закрашенные квадратики.
А сейчас давайте немного окунёмся в историю.
Косички по клеточкам (видео)
Какой след рисунки п
о тетрадным клеткам оставили в истории?Безусловно, каждый из нас, чьё детство прошло в 80-е или 90-е, даст ответ на этот вопрос. И ответ на него простой – видеоигры!
Все мы помним легендарные игры из нашего детства: Марио, «танчики», Pacman, Donkey Kong и многие другие. Об этих играх знают и наши дети, но в курсе ли они, что Марио не всегда был трёхмерным?
В наше детство игры были 8-битными, и даже самые красочные пейзажи составлялись по технологии пиксельной графики. Используя эту же технологию, рисуются рисунки по тетрадным клеткам . И кто знает, может быть, легендарный Марио или Donkey Kong тоже когда-то были просто рисунками на полях школьной тетради?
Давайте и мы с вами попробуем нарисовать наш первый рисунок по тетрадным клеткам, и кто знает, может быть, он вдохновит вас на что-то такое, что перевернёт наш мир, как когда-то его перевернуло появление видеоигр.
Что необходимо для рисования простых рисунков по тетрадным клеткам?
Для рисования простых рисунков по клеткам нам понадобятся:
- Чёрная гелиевая ручка
- Фломастеры
Как нарисовать простой рисунок по тетрадным клеткам?
В рисовании простых рисунков по тетрадным клеткам нет ничего сложного. Всё что вам нужно – посчитать клеточки, начертить контур и закрасить рисунок в соответствии с оригиналом. Давайте рассмотрим это подробнее на примере сердечка.
- Возьмите тетрадный лист и чёрную гелиевую ручку, поставьте три крестика так, как это показано на рисунке. Крестики будут означать то, что эти квадратики мы будем закрашивать чёрным цветом.
- Далее нарисуйте линии, которые обозначат границы нашего рисунка в этой области.
- Поставим ещё 6 крестиков сверху, по три крестика с каждой стороны. Обратите внимание на отступы, считайте клеточки, которые нужно оставить пустыми.
- Проведём ещё 2 линии, чтобы обозначить границы рисунка.

5. Поставим ещё по крестику слева и справа, а так же проведём горизонтальную линию под верхними крестиками, обозначая границы в этом месте. Сделайте это так, как показано на рисунке.
6. Проставим 8 крестиков по вертикали, по 4 крестика с каждой стороны, так как это изображено на следующем рисунке.
7. Проведём вертикальную линию слева, а так же линии сверху, так как это сделано на рисунке. Этим мы полностью обозначим верхнюю границу нашего сердечка.
10.Теперь нам осталось обозначить границы сердечка по всему его периметру, так как это сделано на рисунках ниже. Наш рисунок уже напоминает сердечко, однако, это ещё не всё. Теперь мы должны закрасить наше сердечко, чтобы оно приобрело готовый вид.
11. Закрасим внутреннюю часть сердечка красным фломастером, но оставим три клеточки белыми в левом верхнем углу, дабы обозначить световой блик. Сделайте это так, как это показано на рисунке.
12. Последнее, что нам осталось сделать – это закрасить чёрным фломастером те части, которые мы помечали крестиками.
И вот, наш рисунок приобрел свой готовый вид. Теперь вы умеете рисовать простые рисунки по тетрадным клеточкам и можете попробовать свои силы в рисовании других картинок, которые можно без труда найти в интернете по ключевым словам «8bit art ».
Если вы не хотите ограничивать свои умения рисованием простых рисунков, давайте рассмотрим с вами, как рисуются сложные рисунки по клеточкам . Изначально процесс может показаться вам очень сложным, но не отчаивайтесь раньше времени, стоит всего один раз попробовать и вы поймёте, что рисовать подобные рисунки не только просто, но и очень увлекательно!
Что необходимо для рисования сложных
рисунков по тетрадным клеткам ?Для рисования сложных рисунков нам понадобятся:
- Чёрная гелиевая ручка
- Фломастеры или карандаши
- Тетрадь (или тетрадный лист) в клетку
- Компьютер
- Фотография
- Редактор фотографий Adobe Photoshop
В рисовании сложных рисунков , вам тоже придётся просчитывать клеточки, которые нужно закрашивать. Сложность в данном случае заключается только в том, чтобы не ошибиться в просчёте, так как клеточек у нас будет больше, нежели на предыдущем рисунке. А так же наша задача – правильно подобрать оттенки фломастеров или карандашей, чтобы наш рисунок соответствовал фотографии, с которой мы будем его рисовать.
Сложность в данном случае заключается только в том, чтобы не ошибиться в просчёте, так как клеточек у нас будет больше, нежели на предыдущем рисунке. А так же наша задача – правильно подобрать оттенки фломастеров или карандашей, чтобы наш рисунок соответствовал фотографии, с которой мы будем его рисовать.
И так, давайте приступим!
- Для начала давайте подберём фотографию. Я выбрал фотографию милого щенка, которую нашёл в интернете. Вот она:
- Давайте откроем редактор фотографий Adobe Photoshop и загрузим нашу фотографию:
Теперь нам нужно применить фильтр, чтобы обозначить клеточки на фотографии, по которым мы впоследствии будет ориентироваться. Для этого выбираем сверху вкладку «Фильтр» и жмём на параметр «Галерея фильтров».
4. В открывшимся окне выбираем вкладку «Текстура» и один раз кликаем на фильтр «Цветная плитка».
5.Ползунки параметров справа нужно установить следующим образом:
Размер квадратов – 10
Рельеф – 0
Затем нажимаем ОК.
6. Теперь наша фотография разбита на клеточки. Давайте сохраним её на нашем компьютере, чтобы впоследствии её можно было открыть на весь экран, либо распечатать.
- Теперь остаётся только открыть или распечатать нашу фотографию, подобрать карандаши или фломастеры по оттенкам и закрасить клеточки в соответствии с оттенками.
Вот и всё!
Теперь вы умеете рисовать простые и сложные рисунки по клеточкам !
Благодарим вас за ваше внимание!
Следите за нашими новостями и учитесь рисовать вместе с нами!
Рисуем по клеточкам (видео)
Красиво рисовать – могут единицы! А тем, у кого нет особенных способностей – о рисовании остается только мечтать! Ну и любоваться чужими рисунками, конечно же! Еще совсем недавно – так и было! Но теперь – все изменилось, потому что с помощью клеточек любой из нас сможет нарисовать красивую картину! Да-да! Рисунки по клеточкам сложные и большие – ничем не уступают по красоте настоящим картинам!
В детстве многие мечтают стать настоящим художником! Это же так здорово – рисовать красивые рисунки, дарить их своим друзьям и близким! Увы, не всем даны способности и таланты, поэтому чаще всего, в будущем приходится выбирать совсем другие профессии! А на красивые картины – любоваться на выставках! Но сегодня – все изменилось. И нарисовать их сможет каждый! Ведь теперь есть картинки по клеточкам!
И нарисовать их сможет каждый! Ведь теперь есть картинки по клеточкам!
Отсчитав нужное количество клеточек и закрасив их в определенный цвет, вы сможете нарисовать красивый портрет, пейзаж, любимого персонажа или целый сюжет! Вам потребуется немало терпения и внимательности, но результат того стоит! Для больших рисунков лучше всего подойдет миллиметровая бумага, но можно использовать и обычные листы в клетку, склеив их в один большой лист! Хотите попробовать нарисовать настоящую большую картину?
С помощью клеточек можно нарисовать все, что угодно. В тетради или блокноте – небольшие рисунки цветов, животных или любимых персонажей, на большом тетрадном листе – красивую композицию, а на листе миллиметровой бумаги – даже огромный натюрморт или портрет! Все зависит только от сложности выбранного вами образца для перерисовки. Конечно, начинать сразу с огромных картин – не стоит, но если постараться, можно очень быстро перейти от самых простых картинок к гораздо более сложным!
Более сложные рисунки подойдут тем кто уже натренировался на и рисунках по клеточкам, и желает попробовать нарисовать что-то более сложное. В нашей галерее представлены как портреты так и и просто классные рисунки по клеточкам для срисовки в тетради.
В нашей галерее представлены как портреты так и и просто классные рисунки по клеточкам для срисовки в тетради.
Для более сложных рисунков лучше подойдёт миллиметровая бумага.
В Живую это выглядит примерно вот так:
Не каждому удалось окончить художественную школу, чтобы научиться технике рисования. Если хотите сделать креативную открытку или заполнить дневник оригинальными рисунками, освойте рисование по клеточкам. Маленькие картинки по клеточкам смогут сделать даже новички. Главное, купить тетрадку для математики со светлой бумагой.
Как рисовать по клеточкам
Многие любят разгадывать японские кроссворды, в основу которых положено рисование по клеточкам. Если у вас есть готовые разгаданные кроссворды или ответы к ним, то сможете просто перерисовать в свою тетрадку большие фигуры.
Самый хороший способ использовать готовые схемы, которые были специально разработаны для тех, кто не умеет рисовать. Вы можете закрашивать по схеме клеточки в собственной тетради, а потом удивлять красивыми изображениями близких и родных.
Среди шаблонов вы найдете
Оригинально смотрятся фрукты по клеточкам . Если хорошо закрасить рисунок яркими фломастерами, то потом можно его вырезать и использовать для декора интерьера или украшения аппликации.
Хотите сделать открытку или описать в своем дневнике романтическую историю, тогда нарисуйте сердечко по клеткам.
Конфетки, букетики, цветочки – все это можно нарисовать по клеточкам.
Если вы освоите принцип, то потом сможете изображать все, что угодно в своей собственной тетради.
Хотите придумать свой собственный рисунок? Тогда сделайте легкую зарисовку, а потом начинайте превращать ее в рисунок по клеточкам. Начинать лучше всего с контура. Потом можете выделять мелкие детали. Не забудьте отметить, каким цветом, какая деталь должна быть выделена, чтобы рисунок получится ярким и красивым.
3D-рисунки по клеточкам – это хороший способ провести интересно досуг и реализовать свои творческие способности.
Вы еще ни разу не рисовали по клеточкам? Тогда обязательно попробуйте. Это занятие придется по душе как маленьким детям, так и взрослым. Специалисты отметили, что это хобби развивает творческое мышление, координацию движений при письме, концентрацию внимания и логику. Проводите досуг с пользой, выдумывая новые 3Д схемы простые и сложные для рисования по клеточкам.
Это занятие придется по душе как маленьким детям, так и взрослым. Специалисты отметили, что это хобби развивает творческое мышление, координацию движений при письме, концентрацию внимания и логику. Проводите досуг с пользой, выдумывая новые 3Д схемы простые и сложные для рисования по клеточкам.
Сложный рисунок по клеточкам
Предлагаем фото нескольких популярных схем для начинающих
Как нарисовать по клеточкам разные красивые рисунки.
В последнее время набирает популярности способ создания рисунков по клеточкам. Не только детям нравиться рисовать»пиксельные картинки». Взрослые с таким же интересом берутся постигать этот стиль рисования.
Из статьи вы узнаете, как научиться рисовать по клеточкам, какие материалы и навыки необходимы, и подберете схемы рисунков, которые вам больше по душе.
Как научиться рисовать по клеткам для начинающих и детей?
- Не обязательно обладать талантом художника, чтобы переносить на бумагу понравившиеся изображения и формы.
 Рисование по клеточкам — легкий и интересный способ разнообразить свой досуг, заполнить страницы скетчбука или обычного ежедневника.
Рисование по клеточкам — легкий и интересный способ разнообразить свой досуг, заполнить страницы скетчбука или обычного ежедневника. - Для работы используются фломастеры или цветные карандаши ярких цветов. Самые разнообразные рисунки получаются путем закрашивания клетки за клеткой. Используя этот способ рисования можно перенести на бумагу пейзаж, нарисовать человека или зверушку, сказочного персонажа или просто создать красивый и необычный орнамент.
- Если вы решили научиться рисовать по клеточкам, то попробуйте срисовать один из представленных в статье рисунков. Для начала остановитесь на наиболее простом варианте. После того, как рисунок будет готов, вы сможете попробовать перенести на лист бумаги более сложную схему из картинок галереи.
- Используя данный способ рисования, вы точно не будете скучать, ведь попробовав рисовать по клеточкам, вам обязательно захочется продолжить это интересное занятие.
Видео: Как нарисовать по клеточкам Angry Birds
Чем полезно рисование по клеточкам:
- В нашей фотоподборке собраны не просто схемы картинок.
 Каждое изображение — это вариант графического диктанта. Такие картинки стали очень модными сейчас.
Каждое изображение — это вариант графического диктанта. Такие картинки стали очень модными сейчас. - Вероятно, растущий интерес к ним связан с простотой исполнения и тем, что данное занятие еще и очень полезно.
- Рисование по клеточкам способствует развитию усидчивости, обретению навыков письма (если рисует ребенок), развивает логическое и абстрактное мышление, расслабляет.
- Благодаря такому способу рисования можно откорректировать правильность движений при письме, улучшить координацию.
- Забавные картинки словно сами по себе появляются на листе бумаги. За таким занятием не жаль провести свободное время.
Рисунок создается двумя способами:
- первый способ — построчный: заполняются разными цветами строчка за строчкой
- второй способ — клетки закрашиваются поочередно: сначала используется один цвет, потом — другой и так далее
Что понадобится для рисунка:
- цветные карандаши или маркеры (можно использовать фломастеры, простой карандаш, обычную ручку)
- тетрадь в клеточку со светлыми листами или миллиметровая бумага (для создания рисунков большого формата)
- понадобится еще хорошее настроение, немного свободного времени, а еще — множество схем из нашей галереи
Почувствуйте себя настоящим художником! Ваш будущий шедевр может выглядеть очень просто или состоять из нескольких сложных схем.
Схемы рисунков по клеточкам

Как рисовать по клеточкам в тетради маленькие, лёгкие и простые рисунки поэтапно и красиво: схемы
- Если у вас на полочке за плечами нет обучения в художественной школе, но появилось желание научиться технике рисования, то попробуйте освоить метод рисования по клеточкам.
- Оригинальные рисунки, созданные в такой технике, отлично подойдут для создания креативной открытки, для заполнения личного дневника. С маленькой картинкой справиться даже новичок.
- В качестве схем подойдут представленные в нашей статье картинки или разгаданные японские кроссворды, ведь в их основе — рисование по клеточкам.
- Если вы не умеете заполнять клеточки японских кроссвордов, то воспользуйтесь ответами к ним и перерисуйте в тетрадь фигуры большего формата.
- Еще одним вариантом рисования является использование готовых схем, разработанных специально для тех, кто впервые рисует по клеточкам и не имеет навыков рисования.

Ниже представлена фотоподборка рисунков по клеточкам:
Видео: Рисуем по клеточкам — ЧЕЛОВЕК ПАУК
Как нарисовать по клеточкам разные красивые рисунки для личного дневника, в тетради?
- Красиво нарисованную картинку можно использовать в качестве декора для интерьера. Для этого картинка обрезается по контуру и клеится на плотную бумагу. Потом ярко разукрашенный рисунок можно поместить в рамочку.
- Поместив в самодельную рамочку рисунок в клеточку, можно превратить его в креативный подарок хенд-мейд.
- Рисунок по клеточкам может стать элементом аппликации. Вы можете сделать модные открытки, украсив их рисунками в клеточку или «проиллюстрировать» записанную в дневнике романтическую историю. Сердечки, нарисованные по клеткам, лица девушек или парней, герои мультфильмов, пирожные, конфеты, цветочки — любой образ можно создать, используя данный способ рисования.

- Такой способ рисования станет прекрасным тренажером для отработки мелкой моторики. Потому это занятие полезно не только для детей, но и для взрослых. Насладиться творчеством можно после того, как одна из предложенных в нашей подборке схем будет полностью перенесена в вашу тетрадь.
- Можно использовать и часть схемы. Например, если вы хотите изобразить какое-то животное не полностью, а ограничиться рисованием лишь отдельно взятого элемента для заполнения страницы дневника картинкой.
Освоив принцип создания рисунков по клеточкам, вы сможете сами придумывать схемы и рисовать любые понравившиеся объекты в тетради.
Как рисовать собственный рисунок?
- обдумываем, что мы хотим изобразить
- делаем легкую зарисовку
- превращаем первоначальные линии в рисунок по клеточкам
- в первую очередь обрисовываем контуры
- переходим к выделению мелких деталей
- отмечаем, какая деталь каким цветом должна быть закрашена (это необходимо для яркого и красивого рисунка, однако вы можете создавать и черно-белые картинки)
- пополняйте коллекцию собственных 3D схем простыми или сложными картинками по клеточкам
Не стоит копировать увиденный где-то рисунок с точностью, повторять цветовую гамму.
- Чтобы заполнить тетрадь оригинальными картинками, вносите изменения в схемы, меняйте цвета. Пусть эти маленькие картинки станут отражением вашего внутреннего мира.
Как научить рисовать по клеточкам ребенка?
- Рисование по клеточкам поможет ребенку поверить в то, что он может самостоятельно создавать красивые рисунки. А ведь именно от вдохновения в раннем возрасте зависит то, будет ли ребенок обращаться к каким-либо творческим занятиям в будущем.
- Чтобы было удобнее рисовать по клеточкам с ребенком, лучше заранее распечатать понравившийся шаблон.
- Когда у малыша будет готов набор для рисования по клеточкам, включающий тетрадный лист, фломастеры и распечатанный шаблон, можно будет немедленно приступать к рисованию любимых мультяшных героев или зверушек.
- Прежде, чем начинать зарисовывать клеточки в тетради, с ребенком 4-5 лет можно обсудить будущий рисунок. Пусть юное дарование расскажет, какие цвета он будет использовать для рисунка и какие элементы начнет рисовать в первую очередь.

- После обсуждения отберите в малышом фломастеры, которые будете использовать во время рисования.
- Расскажите ребенку о принципах рисования картинок по клеточкам.
- Предложите малышу выбрать клеточку на шаблоне, из которой он начнет «надстраивать» остальные элементы. Спросите, почему именно эта клеточка стала началом рисунка. Найдите вместе с юным художником эту клетку в тетради.
Видео: Рисунок по клеткам # 40 Оленёнок
- Поскольку у ребенка 4-5 лет не достаточно усидчивости, то длительность занятия не должна превышать 15-20 минут. Вернуться к рисунку можно еще раз в течение дня.
- Если вам нужно заинтересовать ребенка, то попробуйте такой способ: перенесите сами схему картинки в клеточку на лист бумаги, упустив один или несколько элементов. Потом попросите ваше юное дарование дорисовать то, чего не хватает на картинке. Для срисовывания недостающей детали малыш может использовать готовую схему.
- При желании, клеточки в схеме рисунка можно заполнять не только разукрашенными квадратиками, но и использовать для заполнения части рисунка разнообразные знаки.
 Такой способ поможет вам создать по-настоящему уникальный рисунок.
Такой способ поможет вам создать по-настоящему уникальный рисунок. - Начинаем переносить схему с правильного расположения рисунка на листе. Картинку можно начинать рисовать с верхней части, а можно с нижней. Все зависит от того, какая у вас схема. Если больше элементов расположено вверху, то и начинать рисунок нужно с этой части, «надстраивая» остальные клеточки.
- Способ рисования по клеточкам можно использовать и для переноса изображения на лист бумаги. Таким образом можно перерисовать все: от выкройки до картины. Рисунок по клеточкам использовался еще до появления кальки или других способов копирования изображения. Можно нарисовать даже лицо знакомого человека или родственника и презентовать необычный автопортрет на день рождения.
Маленькие схемы для вышивки крестом по клеткам
Содержание
- Маленькие схемы для вышивки крестом по клеткам и советы по выбору образа
- Распространенные образы
- Вышивка крестиком: картинки со схемами по клеточкам и практические советы
- Творческая вышивка крестиком для детей: схемы по клеточкам
- Простые схемы вышивки 50 на 50 клеток крестиком
- Популярные записи
Маленькие схемы для вышивки крестом по клеткам и советы по выбору образа
Новичкам рекомендуется выбрать простые схемы, позволяющие в сжатые сроки создать разнообразное изображение. При этом не стоит тревожиться, что упрощенная модель не позволит реализовать сложный проект. Натюрморты, пейзажи, фигурки сказочных персонажей – все это можно вышить крестом. Перед началом работы следует внимательно подойти к вопросу выбора эскиза.
При этом не стоит тревожиться, что упрощенная модель не позволит реализовать сложный проект. Натюрморты, пейзажи, фигурки сказочных персонажей – все это можно вышить крестом. Перед началом работы следует внимательно подойти к вопросу выбора эскиза.
Каждый цвет и крестик на рисунке получит необходимое отражение на этапе вышивки. Благодаря этому, можно провести первоначальную оценку сложности.
Распространенные образы
- Цветочные элементы – полевые цветы, букеты или отдельно стоящие представители флоры украсят гостиную, спальню или прихожую. Молодоженам рекомендуется выбрать розы – символ верности, любви и гармонии. Больше всего времени уйдет на формирование многочисленных изгибов грациозного цветка;
- Маки – вопреки кажущейся простоте, представители мира флоры станут хорошим украшением для помещения, оформленного в классическом стиле. В большинстве случаев вышивка дополняется образами полевых цветов: василек, колоски;
- Пионы – подходят для украшения спальни, являясь признанными символом радости, благоденствия и любви.

Подборку схем для вышивки картин с пионами вы найдёте по ссылке: https://homeli.ru/dekor/vyshivka/krestikom/vyshivka-krestom-piony-skhemy.
На втором месте по частоте встречаемости находятся образы животных. Если необходимо подчеркнуть динамическую составляющую, то выбор делается в пользу бегущих коней. На протяжении веком парнокопытные были символом безудержной энергии и тяги к свободе. Если речь идет о спальне, то здесь уместны лебеди – грациозные птицы не имеют сложных элементов. Для начинающих будет несложно перенести их на материал. Дизайн с акцентом на минимализм подчеркнут грациозные бабочки. Цвет и форма выбирается с учетом преобладающей в помещении цвета обоев и мебели.
Варианты схем по вышивке необычных картин с бабочками представлены в нашем материале: https://homeli.ru/dekor/vyshivka/krestikom/skhema-vyshivki-krestom-babochki.
Вышивка крестиком: картинки со схемами по клеточкам и практические советы
Новичкам каждый шаг необходимо сверять с выбранной схемой. В этом случае не придется переделывать значительный кусок работы. Начинать процесс вышивания следует с заранее отмеченной середины. В этом случае намного проще будет перенести на ткань рисунок. Если необходимо сделать «крест» или «полукрест», то для этого используется 2 сложения. После того как сделан шов, игла возвращается на исходную позицию.
В этом случае не придется переделывать значительный кусок работы. Начинать процесс вышивания следует с заранее отмеченной середины. В этом случае намного проще будет перенести на ткань рисунок. Если необходимо сделать «крест» или «полукрест», то для этого используется 2 сложения. После того как сделан шов, игла возвращается на исходную позицию.
Необходимо убедиться, что каждый «крест» покрывает собой 2 нити на материале. Речь идет о покрытии по ширине и высоте.
Вне зависимости от сложности выбранного эскиза, для работы понадобится следующее:
- Основа для вышки – подбирается с учетом особенностей и количества элементов на выбранном эскизе. Учитывается не только плотность их размещения, но и тип выбранных ниток. Для работы подойдет канва, габардин или лен. Если в доме есть люди, страдающие аллергией, то предпочтение отдается натуральному материалу;
- Разноцветные нитки – мастер не ограничен в вопросе их выбора. Подойдут мулине, «кировские» или DMC. Цветовая гамма выбирается с учетом особенностей выбранного эскиза;
- Остроконечная игла с большим «ушком»;
- Содержащий выбранный рисунок набор;
- Станок для вышивания;
- Пяльца;
- Маркеры для разметки канвы – применяются они, если вышивание крестиком предусматривает создание массивных изображений.
 Перед началом работы необходимо убедиться, что следы от маркера легко смыть водой. В противном случае, работа окажется испорчена.
Перед началом работы необходимо убедиться, что следы от маркера легко смыть водой. В противном случае, работа окажется испорчена.
В процессе создания картины необходимо помнить о правильной геометрии стежков. Они всегда располагаются в одном заранее выбранном направлении. От использования больших протяжек рекомендуется отказаться, иначе картина не будет иметь завершенный вид.
Творческая вышивка крестиком для детей: схемы по клеточкам
Творческая реализация необходима не только взрослым, но и детям. В связи с этим вышивка по клеточкам несколько особых разновидностей. Для работы понадобятся нитки мулине в четыре сложения, игла 3 см с широким «ушком», ножницы, 18 см пяльцы, вафельное полотно с плотной структурой, игольница, несколько полосок картона с заранее нанесенной разметкой.
Взрослые должны помнить, что дети всегда очень активны, поэтому необходимо рассказать несовершеннолетним о правилах техники безопасности.
Дальнейший порядок действий выглядит следующим образом:
- Рассмотреть вместе с ребенком будущую схему для вышивания по клеткам.
 При желании, ребята могут самостоятельно дополнить представленный эскиз;
При желании, ребята могут самостоятельно дополнить представленный эскиз; - Воспользовавшись картонными горизонтальными полосками, ребята вместе с педагогом дорисовывают необходимое количество дополнительных элементов.
- Создание картинки начинается снизу вверх. Перед каждым действием необходимо взглянуть на схему для правильного подсчета крестиков. После того как на схеме найдена середина, необходимо при помощи простого карандаша и линейки несколько линий;
- Если все сделано верно, то картинка окажется заключенной в квадрат. После этого ткань складывается пополам. Найдя нижний горизонтальный ряд, необходимо отсчитать определенное количество клеток. Именно в этой точки и делается первый стежок;
- В процессе создания картинки необходимо оставить по краям ткани до 2 см для создания декоративной рамки.
Детям подойдёт схемы попроще. В нашей следующей статье представлена подборка идей для вышивания: https://homeli.ru/dekor/vyshivka/krestikom/skhema-vyshivki-krestom-dlya-detej.
Простые схемы вышивки 50 на 50 клеток крестиком
Украшение просторной комнаты при помощи вышивки требует определенных навыков. В первую очередь подбираются легкие по наполнению картины. Речь может идти о цветах или абстрактных формах. Цветовая гамма подбирается с учетом помещения и степени его освещенности. Если окна выходят на солнечную сторону, то слишком яркая вышивка в буквальном смысле утонет в световом потоке. Для комнаты, где солнце только несколько часов в течение суток, подойдут теплые пастельные тона. Еще одна рекомендация связана с геометрической формой вышивки.
Квадратные и маленькие – вот основа правильного наполнения интерьера декоративными элементами.
Для реализации проекта понадобится выполнить следующие рекомендации:
- Выбрать схему, созданную в формате 50 на 50 клеток;
- Ткань для переноса изображения выбирается с учетом плотности и насыщенности рисунка небольшими элементами;
- Во время проведения линий желательно использовать измерительные приборы.
 Отклонение даже на 0,5 см приведет к необходимости все переделать;
Отклонение даже на 0,5 см приведет к необходимости все переделать; - Каждый стежок покрывает с обеих сторон проведенную на ткани линию;
- Основной успешной реализации проекта является осторожность. Если необходимого опыта у человека пока нет, каждый стежок проводится после согласования предполагаемого действия со схемой.
маленькие для детей, легкие 50 на 50 для начинающих
Компания “Фокус” – строительство домов из теплоблоков, экоблоков > Статьи
Благодаря простой схеме вышивки крестом возможно создать самую разнообразную картину со своей историей Даже в современном искусстве есть место вечному – вышивке крестом по схемам в клеточку. Благодаря технически правильно разработанным схемам, справиться, даже со сложной композицией, под силу и новичку.
Содержание:
- Схемы вышивки крестиком по клеткам: учимся понимать
- Вышивка крестиком: схемы по клеточкам для начинающих
- Маленькие схемы для вышивки крестом по клеткам
- Как вышивать крестиком: осваиваем простой счетный крест
- Современная вышивка крестиком: схемы по клеточкам, картинки (видео)
Схемы вышивки крестиком по клеткам: учимся понимать
Ни одно вышивание крестиком не обходится без схемы – разлинованного на маленькие квадратики рисунка. Каждая клеточка на схеме обозначает вышитый крестик на ткани.
Каждая клеточка на схеме обозначает вышитый крестик на ткани.
Процесс обучения вышивания крестом довольно прост. От рукодельницы потребуется только усидчивость, немного времени и терпение.
Очень часто, для удобства и простого понимания, квадраты схемы помечают различными цветами и значками.
Перед началом работы внимательно изучите соответствие номеров ниток и их значков, чтобы не совершить ошибку при вышивании
Итак, разберемся со всем детально:
- Схема разделена на крупные квадраты 10 на 10 клеток, что помогает в процессе подготовки ткани-основы к вышивке, а также при сравнении вышивки на канве с рисунком на бумаге;
- У всех моточков мулине есть свой номер, а если на схеме есть квадраты, которые никак не обозначены, значит, на канве в этих местах ничего не нужно вышивать;
- Для того чтобы ваши картинки смотрелись гармонично и были расположены правильно, начинать вышивку нужно строго с середины канвы.

Для удобства и правильности расчетов можно карандашом или специальным фломастером обозначить центр полотна стрелочками.
Вышивка крестиком: схемы по клеточкам для начинающих
За счет несложных схем вышивки крестом по клетке получается множество роскошных картин. Довольно часто крестиком вышивают цветы, например букеты в вазе или те, которые растут на лугу.
Чаще всего в вышивке крестом встречаются такие цветы:
- Розы – это легкие красивые цветы, которые символизируют верность, взаимную любовь и дарят полное взаимопонимание мужчины и женщины. Поэтому их изображения так нравятся многим рукодельницам. Вышитое изображение розы приносит гармонию в семейных отношениях.
- Маки. Внешне они довольно просты, но такие цветы смотрятся очень оригинально и интересно. Считается, что вышивка этих цветов оберегает от бед и ненастий. Часто такие картины дополняют полевыми цветами: васильками, колосками или ромашками.

- Пионы. Цветы эти в вышивке крестом выглядят превосходно. Они символизируют благополучие, любовь и радость.
Вам будет намного легче, если вы начнете с любимой и простой схемы для вышивания
Красочный натюрморт способен украсить любую кухню, цветы подойдут для гостиной или спальной комнаты, а картину с ангелочками можно подарить на рождение ребенка.
При выборе эскиза для вышивки крестиком стоит учитывать все элементы на рисунке, это поможет в получении хорошего результата.
Маленькие схемы для вышивки крестом по клеткам
Для начинающих рукодельниц хорошо подойдут маленькие простые схемы, которые помогут освоить искусство воссоздания рисунков по клеточкам. Не стоит сразу браться за сложные работы. Стоит набраться опыта. И тогда в будущем такие картины можно будет преподнести в качестве оригинального подарка своим родным и близким на любой праздник.
Маленькие схемы для вышивки крестом хороши тем, что можно быстрее завершить процесс рукоделия и начать создавать новые шедевры
Специфика технологии маленьких вышивок:
- Мелкий крестик на ткани для вышивания поможет сделать рисунок красивее и эффектней, поэтому лучше, чтобы клетка на ткани была маленькой.
- Есть схемы с очень большой палитрой цветов. Некоторые рукодельницы считают, что работа с такими схемами более кропотлива и непроста, так как иногда нужно вышить лишь несколько крестиков одинаковым цветом, нужно постоянно менять нитку. Есть и такие схемы, в которых диапазон цветов меньше, правда с ними изделия могут выглядеть несколько примитивно. Что лучше выбирать вам.
- При выполнении маленьких вышивок важным технологическим моментом является обводка. С ней вышивка приобретет форму и четкость.
Обводка – это обведение контура фигур вышивки тонкой линией нити.
Обводка делается следующим образом: сначала обводится контур в одну сторону через стежок, после чего в обратном направлении заполняются пропущенные стежки, а иголка опускается в противоположную дырочку на ткани. Обводкой при необходимости оформляются также линии внутри фигуры.
Обводкой при необходимости оформляются также линии внутри фигуры.
Как вышивать крестиком: осваиваем простой счетный крест
На сегодняшний день существует уже более 30 самых разнообразных видов вышивки крестиком. Однако именно обычная вышивка крестом до сих пор остается самой популярной среди рукодельниц. Она проста и универсальна, поэтому вышивать крестиком учат даже детей.
Один из вариантов выполнения простого счетного креста – каждый крест вышивается целиком, а затем вышивается следующий
Обычный крестик сделан из двух наклонных стежков. Следует просто поднять иголку через отверстие снизу в канве, положить диагональный стежок на ткань и пропустить иголку через противоположную дырочку в канве.
Важно! Для того чтобы работа выглядела аккуратно, крестики должны быть вышиты с уклоном на одну сторону.
Современная вышивка крестиком: схемы по клеточкам, картинки (видео)
Рукодельницы, которые садятся за вышивание крестиком первый раз, могут сделать несколько тренировочных рядов крестиком на отдельном кусочке канвы. Это необходимо для того, чтобы «набить» руку и набраться уверенности в себе. После можно будет переходить к «чистовому» варианту канвы и смело творить.
Похожие статьи
- Схемы для вышивки крестом для начинающих: маленькие бесплатно, видео правильное для детей, как шить, скачать фото
Вышивание – это интересное хобби, которое на сегодняшний день является популярным среди многих рукодельниц Сегодня вышивка становится все более…
- Наборы больших вышивок крестом: большой размер и бесплатные схемы, скачать без регистрации, формат крупный
Украсить интерьер помещения можно при помощи красивой и уникальной вышитой картины Вышивки крестом ручной работы всегда отличаются своей красотой и…
- Вышивка крестом павлина: схему бесплатно скачать, набор белый без регистрации, монохром гладью и китайские пальмы
С самых древних времен в истории человечества павлины символизировали царственное величие, аристократизм и благородство, достаток и красоту, и поэтому.
 ..
..
Вышивка по клеточкам схемы – Дневник строителя Pilonstroy.ru
Рейтинг статьиЗагрузка…
Вышивка крестиком: схемы по клеточкам и 30 картинок
Человеку не требуются глубокие знания в области рукоделия и значительный опыт. Опираясь на изображенные на бумаге символы и цвета, человек стежок за стежком создает рисунок из нитей. Каждая клеточка на поле соотносится только с одним стежком или крестиком.
Маленькие схемы для вышивки крестом по клеткам и советы по выбору образа
Новичкам рекомендуется выбрать простые схемы, позволяющие в сжатые сроки создать разнообразное изображение. При этом не стоит тревожиться, что упрощенная модель не позволит реализовать сложный проект. Натюрморты, пейзажи, фигурки сказочных персонажей – все это можно вышить крестом. Перед началом работы следует внимательно подойти к вопросу выбора эскиза.
Каждый цвет и крестик на рисунке получит необходимое отражение на этапе вышивки.
Благодаря этому, можно провести первоначальную оценку сложности.
Распространенные образы
- Цветочные элементы – полевые цветы, букеты или отдельно стоящие представители флоры украсят гостиную, спальню или прихожую. Молодоженам рекомендуется выбрать розы – символ верности, любви и гармонии. Больше всего времени уйдет на формирование многочисленных изгибов грациозного цветка;
- Маки – вопреки кажущейся простоте, представители мира флоры станут хорошим украшением для помещения, оформленного в классическом стиле. В большинстве случаев вышивка дополняется образами полевых цветов: василек, колоски;
- Пионы – подходят для украшения спальни, являясь признанными символом радости, благоденствия и любви.
На втором месте по частоте встречаемости находятся образы животных. Если необходимо подчеркнуть динамическую составляющую, то выбор делается в пользу бегущих коней. На протяжении веком парнокопытные были символом безудержной энергии и тяги к свободе. Если речь идет о спальне, то здесь уместны лебеди – грациозные птицы не имеют сложных элементов. Для начинающих будет несложно перенести их на материал. Дизайн с акцентом на минимализм подчеркнут грациозные бабочки. Цвет и форма выбирается с учетом преобладающей в помещении цвета обоев и мебели.
Если речь идет о спальне, то здесь уместны лебеди – грациозные птицы не имеют сложных элементов. Для начинающих будет несложно перенести их на материал. Дизайн с акцентом на минимализм подчеркнут грациозные бабочки. Цвет и форма выбирается с учетом преобладающей в помещении цвета обоев и мебели.
Вышивка крестиком: картинки со схемами по клеточкам и практические советы
Новичкам каждый шаг необходимо сверять с выбранной схемой. В этом случае не придется переделывать значительный кусок работы. Начинать процесс вышивания следует с заранее отмеченной середины. В этом случае намного проще будет перенести на ткань рисунок. Если необходимо сделать «крест» или «полукрест», то для этого используется 2 сложения. После того как сделан шов, игла возвращается на исходную позицию.
Необходимо убедиться, что каждый «крест» покрывает собой 2 нити на материале. Речь идет о покрытии по ширине и высоте.
Вне зависимости от сложности выбранного эскиза, для работы понадобится следующее:
- Основа для вышки – подбирается с учетом особенностей и количества элементов на выбранном эскизе.
 Учитывается не только плотность их размещения, но и тип выбранных ниток. Для работы подойдет канва, габардин или лен. Если в доме есть люди, страдающие аллергией, то предпочтение отдается натуральному материалу;
Учитывается не только плотность их размещения, но и тип выбранных ниток. Для работы подойдет канва, габардин или лен. Если в доме есть люди, страдающие аллергией, то предпочтение отдается натуральному материалу; - Разноцветные нитки – мастер не ограничен в вопросе их выбора. Подойдут мулине, «кировские» или DMC. Цветовая гамма выбирается с учетом особенностей выбранного эскиза;
- Остроконечная игла с большим «ушком»;
- Содержащий выбранный рисунок набор;
- Станок для вышивания;
- Пяльца;
- Маркеры для разметки канвы – применяются они, если вышивание крестиком предусматривает создание массивных изображений. Перед началом работы необходимо убедиться, что следы от маркера легко смыть водой. В противном случае, работа окажется испорчена.
В процессе создания картины необходимо помнить о правильной геометрии стежков. Они всегда располагаются в одном заранее выбранном направлении. От использования больших протяжек рекомендуется отказаться, иначе картина не будет иметь завершенный вид.
Творческая вышивка крестиком для детей: схемы по клеточкам
Творческая реализация необходима не только взрослым, но и детям. В связи с этим вышивка по клеточкам несколько особых разновидностей. Для работы понадобятся нитки мулине в четыре сложения, игла 3 см с широким «ушком», ножницы, 18 см пяльцы, вафельное полотно с плотной структурой, игольница, несколько полосок картона с заранее нанесенной разметкой.
Взрослые должны помнить, что дети всегда очень активны, поэтому необходимо рассказать несовершеннолетним о правилах техники безопасности.
Дальнейший порядок действий выглядит следующим образом:
- Рассмотреть вместе с ребенком будущую схему для вышивания по клеткам. При желании, ребята могут самостоятельно дополнить представленный эскиз;
- Воспользовавшись картонными горизонтальными полосками, ребята вместе с педагогом дорисовывают необходимое количество дополнительных элементов.
- Создание картинки начинается снизу вверх.
 Перед каждым действием необходимо взглянуть на схему для правильного подсчета крестиков. После того как на схеме найдена середина, необходимо при помощи простого карандаша и линейки несколько линий;
Перед каждым действием необходимо взглянуть на схему для правильного подсчета крестиков. После того как на схеме найдена середина, необходимо при помощи простого карандаша и линейки несколько линий; - Если все сделано верно, то картинка окажется заключенной в квадрат. После этого ткань складывается пополам. Найдя нижний горизонтальный ряд, необходимо отсчитать определенное количество клеток. Именно в этой точки и делается первый стежок;
- В процессе создания картинки необходимо оставить по краям ткани до 2 см для создания декоративной рамки.
Простые схемы вышивки 50 на 50 клеток крестиком
Украшение просторной комнаты при помощи вышивки требует определенных навыков. В первую очередь подбираются легкие по наполнению картины. Речь может идти о цветах или абстрактных формах. Цветовая гамма подбирается с учетом помещения и степени его освещенности. Если окна выходят на солнечную сторону, то слишком яркая вышивка в буквальном смысле утонет в световом потоке. Для комнаты, где солнце только несколько часов в течение суток, подойдут теплые пастельные тона. Еще одна рекомендация связана с геометрической формой вышивки.
Еще одна рекомендация связана с геометрической формой вышивки.
Квадратные и маленькие – вот основа правильного наполнения интерьера декоративными элементами.
Для реализации проекта понадобится выполнить следующие рекомендации:
- Выбрать схему, созданную в формате 50 на 50 клеток;
- Ткань для переноса изображения выбирается с учетом плотности и насыщенности рисунка небольшими элементами;
- Во время проведения линий желательно использовать измерительные приборы. Отклонение даже на 0,5 см приведет к необходимости все переделать;
- Каждый стежок покрывает с обеих сторон проведенную на ткани линию;
- Основной успешной реализации проекта является осторожность. Если необходимого опыта у человека пока нет, каждый стежок проводится после согласования предполагаемого действия со схемой.
Как начать вышивать по схеме (видео)
Декоративная вышивка крестиком на ткани – увлекательное и доступное представителям различных возрастов занятие. Не нужно иметь углубленные знания в области рукоделья. Достаточно подключить воображение для того чтобы перенести желаемое изображение на ткань. Во время работы необходимо постоянно сверяться с выбранной схемой. В этом случае не придется часть переделывать неверный стежок.
Не нужно иметь углубленные знания в области рукоделья. Достаточно подключить воображение для того чтобы перенести желаемое изображение на ткань. Во время работы необходимо постоянно сверяться с выбранной схемой. В этом случае не придется часть переделывать неверный стежок.
голоса
Рейтинг статьи
Оценка статьи:
Загрузка…
Adblock
detector
Вышивка по клеточкам схемы – Строй журнал artikagroup.ru
Рейтинг статьиЗагрузка…
Вышивка крестиком: схемы по клеточкам и 30 картинок
Человеку не требуются глубокие знания в области рукоделия и значительный опыт. Опираясь на изображенные на бумаге символы и цвета, человек стежок за стежком создает рисунок из нитей. Каждая клеточка на поле соотносится только с одним стежком или крестиком.
Маленькие схемы для вышивки крестом по клеткам и советы по выбору образа
Новичкам рекомендуется выбрать простые схемы, позволяющие в сжатые сроки создать разнообразное изображение. При этом не стоит тревожиться, что упрощенная модель не позволит реализовать сложный проект. Натюрморты, пейзажи, фигурки сказочных персонажей – все это можно вышить крестом. Перед началом работы следует внимательно подойти к вопросу выбора эскиза.
При этом не стоит тревожиться, что упрощенная модель не позволит реализовать сложный проект. Натюрморты, пейзажи, фигурки сказочных персонажей – все это можно вышить крестом. Перед началом работы следует внимательно подойти к вопросу выбора эскиза.
Каждый цвет и крестик на рисунке получит необходимое отражение на этапе вышивки. Благодаря этому, можно провести первоначальную оценку сложности.
Распространенные образы
- Цветочные элементы – полевые цветы, букеты или отдельно стоящие представители флоры украсят гостиную, спальню или прихожую. Молодоженам рекомендуется выбрать розы – символ верности, любви и гармонии. Больше всего времени уйдет на формирование многочисленных изгибов грациозного цветка;
- Маки – вопреки кажущейся простоте, представители мира флоры станут хорошим украшением для помещения, оформленного в классическом стиле. В большинстве случаев вышивка дополняется образами полевых цветов: василек, колоски;
- Пионы – подходят для украшения спальни, являясь признанными символом радости, благоденствия и любви.

На втором месте по частоте встречаемости находятся образы животных. Если необходимо подчеркнуть динамическую составляющую, то выбор делается в пользу бегущих коней. На протяжении веком парнокопытные были символом безудержной энергии и тяги к свободе. Если речь идет о спальне, то здесь уместны лебеди – грациозные птицы не имеют сложных элементов. Для начинающих будет несложно перенести их на материал. Дизайн с акцентом на минимализм подчеркнут грациозные бабочки. Цвет и форма выбирается с учетом преобладающей в помещении цвета обоев и мебели.
Вышивка крестиком: картинки со схемами по клеточкам и практические советы
Новичкам каждый шаг необходимо сверять с выбранной схемой. В этом случае не придется переделывать значительный кусок работы. Начинать процесс вышивания следует с заранее отмеченной середины. В этом случае намного проще будет перенести на ткань рисунок. Если необходимо сделать «крест» или «полукрест», то для этого используется 2 сложения. После того как сделан шов, игла возвращается на исходную позицию.
Необходимо убедиться, что каждый «крест» покрывает собой 2 нити на материале. Речь идет о покрытии по ширине и высоте.
Вне зависимости от сложности выбранного эскиза, для работы понадобится следующее:
- Основа для вышки – подбирается с учетом особенностей и количества элементов на выбранном эскизе. Учитывается не только плотность их размещения, но и тип выбранных ниток. Для работы подойдет канва, габардин или лен. Если в доме есть люди, страдающие аллергией, то предпочтение отдается натуральному материалу;
- Разноцветные нитки – мастер не ограничен в вопросе их выбора. Подойдут мулине, «кировские» или DMC. Цветовая гамма выбирается с учетом особенностей выбранного эскиза;
- Остроконечная игла с большим «ушком»;
- Содержащий выбранный рисунок набор;
- Станок для вышивания;
- Пяльца;
- Маркеры для разметки канвы – применяются они, если вышивание крестиком предусматривает создание массивных изображений.
 Перед началом работы необходимо убедиться, что следы от маркера легко смыть водой. В противном случае, работа окажется испорчена.
Перед началом работы необходимо убедиться, что следы от маркера легко смыть водой. В противном случае, работа окажется испорчена.
В процессе создания картины необходимо помнить о правильной геометрии стежков. Они всегда располагаются в одном заранее выбранном направлении. От использования больших протяжек рекомендуется отказаться, иначе картина не будет иметь завершенный вид.
Творческая вышивка крестиком для детей: схемы по клеточкам
Творческая реализация необходима не только взрослым, но и детям. В связи с этим вышивка по клеточкам несколько особых разновидностей. Для работы понадобятся нитки мулине в четыре сложения, игла 3 см с широким «ушком», ножницы, 18 см пяльцы, вафельное полотно с плотной структурой, игольница, несколько полосок картона с заранее нанесенной разметкой.
Взрослые должны помнить, что дети всегда очень активны, поэтому необходимо рассказать несовершеннолетним о правилах техники безопасности.
Дальнейший порядок действий выглядит следующим образом:
- Рассмотреть вместе с ребенком будущую схему для вышивания по клеткам.
 При желании, ребята могут самостоятельно дополнить представленный эскиз;
При желании, ребята могут самостоятельно дополнить представленный эскиз; - Воспользовавшись картонными горизонтальными полосками, ребята вместе с педагогом дорисовывают необходимое количество дополнительных элементов.
- Создание картинки начинается снизу вверх. Перед каждым действием необходимо взглянуть на схему для правильного подсчета крестиков. После того как на схеме найдена середина, необходимо при помощи простого карандаша и линейки несколько линий;
- Если все сделано верно, то картинка окажется заключенной в квадрат. После этого ткань складывается пополам. Найдя нижний горизонтальный ряд, необходимо отсчитать определенное количество клеток. Именно в этой точки и делается первый стежок;
- В процессе создания картинки необходимо оставить по краям ткани до 2 см для создания декоративной рамки.
Простые схемы вышивки 50 на 50 клеток крестиком
Украшение просторной комнаты при помощи вышивки требует определенных навыков. В первую очередь подбираются легкие по наполнению картины. Речь может идти о цветах или абстрактных формах. Цветовая гамма подбирается с учетом помещения и степени его освещенности. Если окна выходят на солнечную сторону, то слишком яркая вышивка в буквальном смысле утонет в световом потоке. Для комнаты, где солнце только несколько часов в течение суток, подойдут теплые пастельные тона. Еще одна рекомендация связана с геометрической формой вышивки.
Речь может идти о цветах или абстрактных формах. Цветовая гамма подбирается с учетом помещения и степени его освещенности. Если окна выходят на солнечную сторону, то слишком яркая вышивка в буквальном смысле утонет в световом потоке. Для комнаты, где солнце только несколько часов в течение суток, подойдут теплые пастельные тона. Еще одна рекомендация связана с геометрической формой вышивки.
Квадратные и маленькие – вот основа правильного наполнения интерьера декоративными элементами.
Для реализации проекта понадобится выполнить следующие рекомендации:
- Выбрать схему, созданную в формате 50 на 50 клеток;
- Ткань для переноса изображения выбирается с учетом плотности и насыщенности рисунка небольшими элементами;
- Во время проведения линий желательно использовать измерительные приборы. Отклонение даже на 0,5 см приведет к необходимости все переделать;
- Каждый стежок покрывает с обеих сторон проведенную на ткани линию;
- Основной успешной реализации проекта является осторожность.
 Если необходимого опыта у человека пока нет, каждый стежок проводится после согласования предполагаемого действия со схемой.
Если необходимого опыта у человека пока нет, каждый стежок проводится после согласования предполагаемого действия со схемой.
Как начать вышивать по схеме (видео)
Декоративная вышивка крестиком на ткани – увлекательное и доступное представителям различных возрастов занятие. Не нужно иметь углубленные знания в области рукоделья. Достаточно подключить воображение для того чтобы перенести желаемое изображение на ткань. Во время работы необходимо постоянно сверяться с выбранной схемой. В этом случае не придется часть переделывать неверный стежок.
Вышивка крестиком схемы по клеточкам картинки: маленькие для детей, легкие 50 на 50 для начинающих
Благодаря простой схеме вышивки крестом возможно создать самую разнообразную картину со своей историей
Даже в современном искусстве есть место вечному – вышивке крестом по схемам в клеточку. Благодаря технически правильно разработанным схемам, справиться, даже со сложной композицией, под силу и новичку.
Схемы вышивки крестиком по клеткам: учимся понимать
Ни одно вышивание крестиком не обходится без схемы – разлинованного на маленькие квадратики рисунка. Каждая клеточка на схеме обозначает вышитый крестик на ткани.
Процесс обучения вышивания крестом довольно прост. От рукодельницы потребуется только усидчивость, немного времени и терпение.
Очень часто, для удобства и простого понимания, квадраты схемы помечают различными цветами и значками.
Перед началом работы внимательно изучите соответствие номеров ниток и их значков, чтобы не совершить ошибку при вышивании
Итак, разберемся со всем детально:
- Схема разделена на крупные квадраты 10 на 10 клеток, что помогает в процессе подготовки ткани-основы к вышивке, а также при сравнении вышивки на канве с рисунком на бумаге;
- У всех моточков мулине есть свой номер, а если на схеме есть квадраты, которые никак не обозначены, значит, на канве в этих местах ничего не нужно вышивать;
- Для того чтобы ваши картинки смотрелись гармонично и были расположены правильно, начинать вышивку нужно строго с середины канвы.

Для удобства и правильности расчетов можно карандашом или специальным фломастером обозначить центр полотна стрелочками.
Вышивка крестиком: схемы по клеточкам для начинающих
За счет несложных схем вышивки крестом по клетке получается множество роскошных картин. Довольно часто крестиком вышивают цветы, например букеты в вазе или те, которые растут на лугу.
Чаще всего в вышивке крестом встречаются такие цветы:
- Розы – это легкие красивые цветы, которые символизируют верность, взаимную любовь и дарят полное взаимопонимание мужчины и женщины. Поэтому их изображения так нравятся многим рукодельницам. Вышитое изображение розы приносит гармонию в семейных отношениях.
- Маки. Внешне они довольно просты, но такие цветы смотрятся очень оригинально и интересно. Считается, что вышивка этих цветов оберегает от бед и ненастий. Часто такие картины дополняют полевыми цветами: васильками, колосками или ромашками.

- Пионы. Цветы эти в вышивке крестом выглядят превосходно. Они символизируют благополучие, любовь и радость.
Вам будет намного легче, если вы начнете с любимой и простой схемы для вышивания
Красочный натюрморт способен украсить любую кухню, цветы подойдут для гостиной или спальной комнаты, а картину с ангелочками можно подарить на рождение ребенка.
При выборе эскиза для вышивки крестиком стоит учитывать все элементы на рисунке, это поможет в получении хорошего результата.
Маленькие схемы для вышивки крестом по клеткам
Для начинающих рукодельниц хорошо подойдут маленькие простые схемы, которые помогут освоить искусство воссоздания рисунков по клеточкам. Не стоит сразу браться за сложные работы. Стоит набраться опыта. И тогда в будущем такие картины можно будет преподнести в качестве оригинального подарка своим родным и близким на любой праздник.
Маленькие схемы для вышивки крестом хороши тем, что можно быстрее завершить процесс рукоделия и начать создавать новые шедевры
Специфика технологии маленьких вышивок:
- Мелкий крестик на ткани для вышивания поможет сделать рисунок красивее и эффектней, поэтому лучше, чтобы клетка на ткани была маленькой.

- Есть схемы с очень большой палитрой цветов. Некоторые рукодельницы считают, что работа с такими схемами более кропотлива и непроста, так как иногда нужно вышить лишь несколько крестиков одинаковым цветом, нужно постоянно менять нитку. Есть и такие схемы, в которых диапазон цветов меньше, правда с ними изделия могут выглядеть несколько примитивно. Что лучше выбирать вам.
- При выполнении маленьких вышивок важным технологическим моментом является обводка. С ней вышивка приобретет форму и четкость.
Обводка – это обведение контура фигур вышивки тонкой линией нити.
Обводка делается следующим образом: сначала обводится контур в одну сторону через стежок, после чего в обратном направлении заполняются пропущенные стежки, а иголка опускается в противоположную дырочку на ткани. Обводкой при необходимости оформляются также линии внутри фигуры.
Как вышивать крестиком: осваиваем простой счетный крест
На сегодняшний день существует уже более 30 самых разнообразных видов вышивки крестиком. Однако именно обычная вышивка крестом до сих пор остается самой популярной среди рукодельниц. Она проста и универсальна, поэтому вышивать крестиком учат даже детей.
Однако именно обычная вышивка крестом до сих пор остается самой популярной среди рукодельниц. Она проста и универсальна, поэтому вышивать крестиком учат даже детей.
Один из вариантов выполнения простого счетного креста – каждый крест вышивается целиком, а затем вышивается следующий
Обычный крестик сделан из двух наклонных стежков. Следует просто поднять иголку через отверстие снизу в канве, положить диагональный стежок на ткань и пропустить иголку через противоположную дырочку в канве.
Важно! Для того чтобы работа выглядела аккуратно, крестики должны быть вышиты с уклоном на одну сторону.
Современная вышивка крестиком: схемы по клеточкам, картинки (видео)
Рукодельницы, которые садятся за вышивание крестиком первый раз, могут сделать несколько тренировочных рядов крестиком на отдельном кусочке канвы. Это необходимо для того, чтобы «набить» руку и набраться уверенности в себе. После можно будет переходить к «чистовому» варианту канвы и смело творить.
После можно будет переходить к «чистовому» варианту канвы и смело творить.
Обзор маленьких схем для вышивания крестиком для детей
Чтобы заниматься вышивкой, нужно выбрать подходящие ткань, нитки, иголки и другие аксессуары. Легче всего изготовить миниатюрную вышивку крестиком. В процессе работы можно определить свои предпочтения и сделать изящную миниатюру.
Вышивка крестом: мини-вышивки – что это
Этот вид вышивки появился много веков тому назад. Миниатюрной вышивкой занимались представители многих именитых фамилий. Среди них можно назвать Екатерину Медичи, Марию Стюарт, Марию-Антуанетту.
Схема вышивки птиц
Маленькие схемы вышивки крестом дают возможность почти сразу увидеть результат работы. Крохотные изображения можно делать различным образом в зависимости от замыслов и творческих предпочтений мастерицы.
К сведению! Небольшие вышивки можно делать не только крестиком, но и гладью.
Если к делу приступает начинающий, для него это удобная возможность определиться с предпочтительной манерой своей работы. Вышивая миниатюрные рисунки, он может определиться с техническими деталями работы и творческими предпочтениями. В качестве сюжетов могут использоваться, растения, бабочки, животные, узоры и другие.
Вышивая миниатюрные рисунки, он может определиться с техническими деталями работы и творческими предпочтениями. В качестве сюжетов могут использоваться, растения, бабочки, животные, узоры и другие.
Иногда этот вид творчества позволяет красиво замаскировать огрехи в одежде. Если имеется небольшая потертость или повреждения, то их можно прикрыть при помощи подходящей необычной мини-вышивки. Можно применять такие изделия не только для украшения одежды, но и в качестве предметов декора. Такие изображения можно применять для сумочек, косметичек, игольниц или других вещей. Можно изготавливать вышитые открытки.
Головки булавок и медальон
Иногда миниатюрные произведения искусства создаются при помощи очень маленьких стежков. В этом случае картина может содержать, несмотря на небольшие размеры, сложные сочетания цветов. Такие схемы для вышивания крестиком маленькие, но могут быть очень сложными.
Обратите внимание! Еще одной сферой применения является создание оберегов.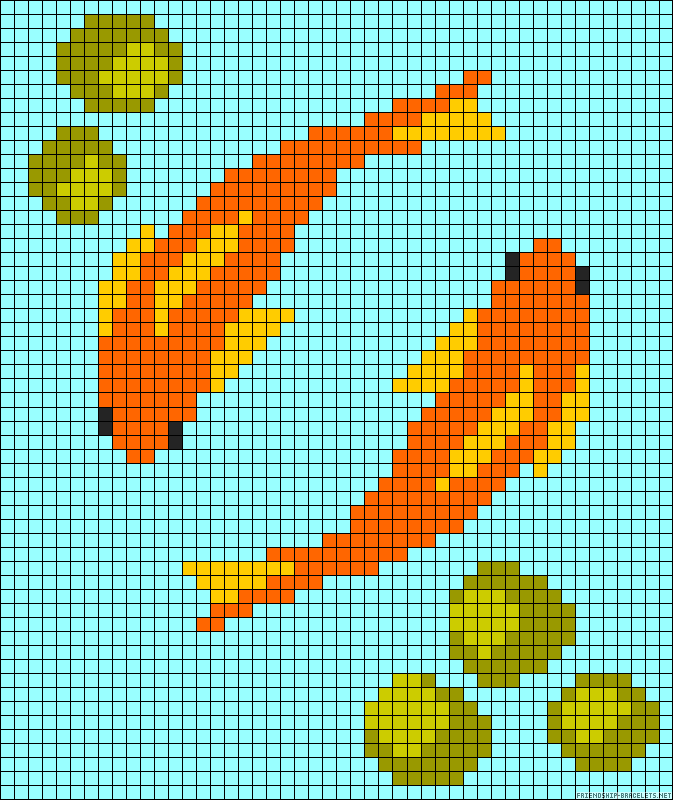 Некоторые считают, что если человек вышивает, думая в это время о хорошем, то он заряжает сделанное им произведение положительной энергией.
Некоторые считают, что если человек вышивает, думая в это время о хорошем, то он заряжает сделанное им произведение положительной энергией.
Такой рисунок может в дальнейшем использоваться в качестве оберега, защищающего от неприятностей.
Необходимые материалы для вышивки крестиком
Чтобы начать маленькие схемы для вышивки крестом, необходимо подготовить ткань и аксессуары. Если планируется сделать рисунок на одежде, скатерти или белье, канву заправляют в пяльцы и начинают с цветочка или другого рисунка. Когда планируют вышивать для декора, то для начинающих наилучшим образом подойдет канва виниловая или Аида. Если нет возможности ими воспользоваться, следует взять любую простую ткань с крупным плетением. Для вышивки крестиком подойдут, например, мешковина, хлопок, лен или холст. На этих тканях имеются хорошо различимые мелкие квадратики, что оказывает существенную помощь при вышивании крестиком.
Перед началом работы над маленькими вышивками края ткани необходимо обметать или закрепить клеем. Некоторые мастерицы для этой цели используют ленты, которые нашивают на края.
Некоторые мастерицы для этой цели используют ленты, которые нашивают на края.
Могут быть использованы такие нитки:
- мулине;
- металлизированные;
- гобеленовая шерсть;
- меланж;
- шелк;
- тонкая шерсть.
Букеты цветов
Обратите внимание! Ткань складывают пополам, чтобы определить центральную точку будущего рисунка. Это поможет правильно расположить вышивку на канве.
Основные инструменты для работы с миниатюрой
При подготовке к выполнению мини-вышивок рекомендуется приобрести несколько разновидностей игл. Это важно для тех, кто только осваивает этот вид искусства. Если попробовать разные варианты, то можно будет найти наиболее подходящий размер для игл.
Использование игл с круглым концом позволит не протыкать ткань, а разделять волокна. При использовании ниток мулине или шерсти стоит выбрать такой формат, где используется большое ушко. Для работы потребуется удобная игольница, в которой будут храниться булавки и иголки.
Вышитый медальон
Важным приспособлением для вышивки маленьких схем крестиком являются пяльцы. Они могут быть деревянными, пластмассовыми или металлическими. Пластмассовые часто соскальзывают, а металлические могут оставить след на ткани. Размер пялец должен подбираться с учетом вкусов мастера и особенностей вышивки
Красивая вышивка крестиком: маленькие схемы
При создании таких вышивок для заполнения клеток применяются несколько видов стежков:
- полукрестик;
- гобеленовый стежок;
- крестик и другие.
Применяется две схемы: обычная и датская. В первом случае, сначала вышивают крестик в одном квадратике и затем переходят к следующему. Во втором – делают весь рисунок диагональными стежками одного направления, затем нужно вышить узор перпендикулярными стежками.
Перед тем, как начать работу, нужно продумать следующие вопросы:
- Выбрать ткань, на которой будет выполняться вышивка.
- Нужно продумать сюжет. Он должен соответствовать тому, где будет использоваться изделие.
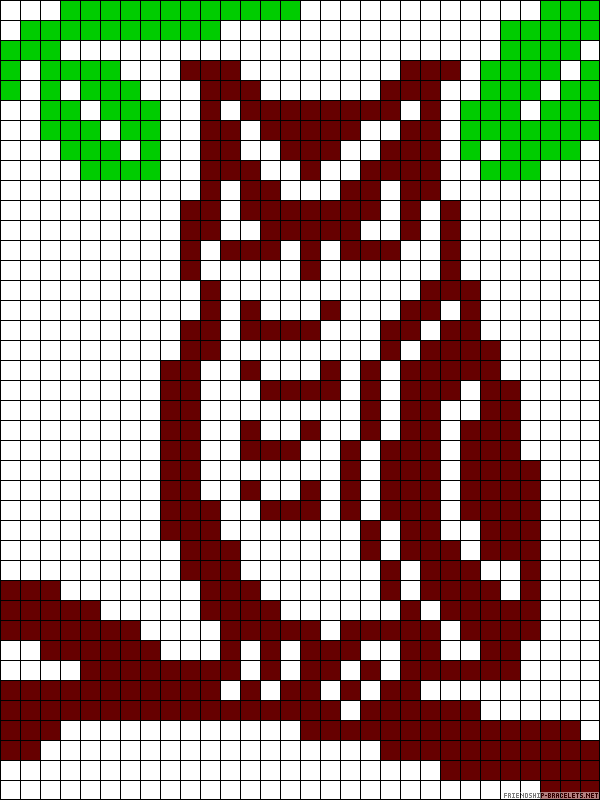
Когда создаются маленькие вышивки крестиком, схемы могут быть сделаны в черно-белом или цветном виде.
Изображение бабочки
Перед началом работы готовят все необходимое, включая инструменты, материалы и рисунок будущей вышивки.
Белый медведь из дикой природы
Этот вариант хорошо подойдет для детских сюжетов. Кроме этого нужно учитывать, что вышитый белый медведь является символом, который имеет эзотерическое значение. Он одновременно обозначает силу и несокрушимую мощь, и защищает от негативной энергии.
Мини-вышивка белого медведя
Можно сделать декоративный предмет с такой вышивкой и разместить в офисе. Такой талисман будет благоприятствовать карьерному росту. Этот сюжет будет красиво выглядеть, если его использовать для домашнего декора. Его можно использовать в качестве новогоднего мотива.
Мини-сюжеты садовые
Садовые сюжеты с незакрашенным полем выглядят легкими, наполненными воздухом. Рисунки растений, улиток, птиц и цветов создают атмосферу гармонии и беззаботности. Они подойдут в качестве украшений, применяющихся в домашнем декоре. Это могут быть изображения растений или абстрактные узоры.
Они подойдут в качестве украшений, применяющихся в домашнем декоре. Это могут быть изображения растений или абстрактные узоры.
Садовые сюжеты
Эти сюжеты будут хорошо смотреться в детской комнате. Они подойдут для украшения одежды или создания сюжетных брошей. Микровышивки позволят внести новый акцент в уже привычную одежду. Изображение простых садовых изображений в качественном исполнении может выражать особый смысл и создавать интересные смысловые оттенки.
Неразлучники — свадебная кукла
Изображения кукол-неразлучников представляют собой древний славянский оберег.
К сведению! Считается, что неразлучников людям дала богиня Макошь, покровительница семейного очага.
Такие вышивки отлично подойдут для самых разных видов свадебной символики.
Неразлучники
Мини-вышивка неразлучников является подходящим символом для украшения дома. Она обладает силой поддерживать отношения, отводить недобрые взгляды.
Цветочная открытка
Изображения цветов являются символом красоты в любой стране. Такие мини-вышивки имеют универсальное значение. Они подходят и для украшения одежды, и в качестве домашнего декора. Такими вышивками уместно украшать детскую комнату, располагая их на мебели, стенах или создавая элементы декора. Можно помещать их на детскую одежду или аксессуары.
Такие мини-вышивки имеют универсальное значение. Они подходят и для украшения одежды, и в качестве домашнего декора. Такими вышивками уместно украшать детскую комнату, располагая их на мебели, стенах или создавая элементы декора. Можно помещать их на детскую одежду или аксессуары.
Цветочные открытки
Хорошо смотрятся цветы, вышитые на детской одежде или сделанные на женских аксессуарах (например, сумочках или косметичках) или на предметах, которыми пользуется ребенок. Выбирая подходящие цветы, мастерица может выразить различные художественные идеи.
Тепло ладоней — зайчик
Этот сюжет символизирует душевное тепло и доброту. Он является символом, который будет напоминать ребенку о любви мамы и папы, тепле домашнего уюта. Эта мини вышивка подойдет для оформления детской комнаты и будет хорошо выглядеть в качестве одного из символов благополучия семьи.
Тепло ладоней – зайчик
Изображение небольшого зайчика в ладонях создает ощущение человеческого тепла, символизирует крепость семьи и несет положительную энергетику.
Садовые открытки
Использование открыток с вышивкой является не только красивым предметом декора, но и произведением искусства, выражающим творческий замысел мастера. Садовая тема сочетает скромность и простоту с универсальностью символов природы.
Садовые открытки
Применение мини-вышивок с мотивом в виде изображений растений и плодов хорошо подойдет для домашнего декора.
Простейшие схемы для вышивания
Они сочетают в себе красоту природы и символы благополучия.
Наборы для вышивания: основные преимущества
Такие наборы очень востребованы среди тех, кто занимается этим видом искусства.
Обратите внимание! Производители стремятся обеспечить широкий выбор таких товаров, для того чтобы каждая швея могла найти подходящий для себя вариант.
Обычно такие наборы содержат сразу после покупки все необходимое для того, чтобы сразу приступить к работе. Наборы, как правило, включают в себя:
- Набор схем вышивок. Они могут быть выполнены в монохроме или цветном виде.
 Может быть предложен один вариант или комплект узоров или картинок.
Может быть предложен один вариант или комплект узоров или картинок. - В состав набора входит канва Аида, считающаяся одним из наиболее предпочтительных вариантов для вышивки, которая делается по канве.
- Для вышивки обычно предлагается использование ниток мулине. Их разнообразие и количество таковы, что для выполнения работы их должно хватить с запасом.
- Для работы в комплект включены одна или несколько игл, которые наиболее подходят для выполнения данного вида вышиивки.
Вышивка розы
В комплект могут также быть добавлены дополнительные материалы или аксессуары: бисер, ленты, пайетки или другие элементы. Здесь учитывается, что набор может быть приобретен начинающей мастерицей. Обязательно присутствует инструкция о том, как делать вышивку. Иногда на канву наносится контур будущего рисунка.
Вышитые игольницы
Миниатюрные вышивки могут помочь начинающему мастеру овладеть профессией. Однако им присуща самостоятельная художественная ценность. Создавая такие вещи, можно украсить свою жизнь, внести в нее больше красоты и творческого света.
Создавая такие вещи, можно украсить свою жизнь, внести в нее больше красоты и творческого света.
Вышивка крестиком: схемы по клеточкам, картинки для начинающих
Схемы вышивки крестиком по клеткам: учимся понимать
Ни одно вышивание крестиком не обходится без схемы – разлинованного на маленькие квадратики рисунка. Каждая клеточка на схеме обозначает вышитый крестик на ткани.
Процесс обучения вышивания крестом довольно прост. От рукодельницы потребуется только усидчивость, немного времени и терпение.
Очень часто, для удобства и простого понимания, квадраты схемы помечают различными цветами и значками.
Итак, разберемся со всем детально:
- Схема разделена на крупные квадраты 10 на 10 клеток, что помогает в процессе подготовки ткани-основы к вышивке, а также при сравнении вышивки на канве с рисунком на бумаге;
- У всех моточков мулине есть свой номер, а если на схеме есть квадраты, которые никак не обозначены, значит, на канве в этих местах ничего не нужно вышивать;
- Для того чтобы ваши картинки смотрелись гармонично и были расположены правильно, начинать вышивку нужно строго с середины канвы.

Для удобства и правильности расчетов можно карандашом или специальным фломастером обозначить центр полотна стрелочками.
Вышивка крестиком: схемы по клеточкам для начинающих
За счет несложных схем вышивки крестом по клетке получается множество роскошных картин. Довольно часто крестиком вышивают цветы, например букеты в вазе или те, которые растут на лугу.
Чаще всего в вышивке крестом встречаются такие цветы:
- Розы – это легкие красивые цветы, которые символизируют верность, взаимную любовь и дарят полное взаимопонимание мужчины и женщины. Поэтому их изображения так нравятся многим рукодельницам. Вышитое изображение розы приносит гармонию в семейных отношениях.
- Маки. Внешне они довольно просты, но такие цветы смотрятся очень оригинально и интересно. Считается, что вышивка этих цветов оберегает от бед и ненастий. Часто такие картины дополняют полевыми цветами: васильками, колосками или ромашками.

- Пионы. Цветы эти в вышивке крестом выглядят превосходно. Они символизируют благополучие, любовь и радость.
Красочный натюрморт способен украсить любую кухню, цветы подойдут для гостиной или спальной комнаты, а картину с ангелочками можно подарить на рождение ребенка.
При выборе эскиза для вышивки крестиком стоит учитывать все элементы на рисунке, это поможет в получении хорошего результата.
Маленькие схемы для вышивки крестом по клеткам
Для начинающих рукодельниц хорошо подойдут маленькие простые схемы, которые помогут освоить искусство воссоздания рисунков по клеточкам. Не стоит сразу браться за сложные работы. Стоит набраться опыта. И тогда в будущем такие картины можно будет преподнести в качестве оригинального подарка своим родным и близким на любой праздник.
Специфика технологии маленьких вышивок:
- Мелкий крестик на ткани для вышивания поможет сделать рисунок красивее и эффектней, поэтому лучше, чтобы клетка на ткани была маленькой.

- Есть схемы с очень большой палитрой цветов. Некоторые рукодельницы считают, что работа с такими схемами более кропотлива и непроста, так как иногда нужно вышить лишь несколько крестиков одинаковым цветом, нужно постоянно менять нитку. Есть и такие схемы, в которых диапазон цветов меньше, правда с ними изделия могут выглядеть несколько примитивно. Что лучше выбирать вам.
- При выполнении маленьких вышивок важным технологическим моментом является обводка. С ней вышивка приобретет форму и четкость.
Обводка – это обведение контура фигур вышивки тонкой линией нити.
Обводка делается следующим образом: сначала обводится контур в одну сторону через стежок, после чего в обратном направлении заполняются пропущенные стежки, а иголка опускается в противоположную дырочку на ткани. Обводкой при необходимости оформляются также линии внутри фигуры.
Как вышивать крестиком: осваиваем простой счетный крест
На сегодняшний день существует уже более 30 самых разнообразных видов вышивки крестиком. Однако именно обычная вышивка крестом до сих пор остается самой популярной среди рукодельниц. Она проста и универсальна, поэтому вышивать крестиком учат даже детей.
Однако именно обычная вышивка крестом до сих пор остается самой популярной среди рукодельниц. Она проста и универсальна, поэтому вышивать крестиком учат даже детей.
Обычный крестик сделан из двух наклонных стежков. Следует просто поднять иголку через отверстие снизу в канве, положить диагональный стежок на ткань и пропустить иголку через противоположную дырочку в канве.
Важно! Для того чтобы работа выглядела аккуратно, крестики должны быть вышиты с уклоном на одну сторону.
Современная вышивка крестиком: схемы по клеточкам, картинки (видео)
Рукодельницы, которые садятся за вышивание крестиком первый раз, могут сделать несколько тренировочных рядов крестиком на отдельном кусочке канвы. Это необходимо для того, чтобы «набить» руку и набраться уверенности в себе. После можно будет переходить к «чистовому» варианту канвы и смело творить.
голоса
Рейтинг статьи
Оценка статьи:
Загрузка. ..
..
Adblock
detector
Восходящие проекции простых и сложных клеток в слое 6 полосатой коры кошек
1. Ахмед Б., Андерсон Дж. К., Дуглас Р. Дж., Мартин К. М., Нельсон Дж. К. Полинейрональная иннервация шипиковых звездчатых нейронов зрительной коры кошек. J Комп Нейрол. 1994; 341:39–49. [PubMed] [Google Scholar]
2. Bolz J, Gilbert CD. Генерация концевого торможения в зрительной коре через межламинарные связи. Природа. 1986; 320: 362–365. [PubMed] [Google Scholar]
3. Bourassa J, Deschenes M. Кортико-таламические проекции первичной зрительной коры у крыс: исследование отдельных волокон с использованием биоцитина в качестве антероградного индикатора. Неврология. 1995;66:253–263. [PubMed] [Google Scholar]
4. Бояпати Дж., Генри Г. Кортикофугальные аксоны в латеральном коленчатом теле кошки. Мозг Res. 1984; 53: 335–40. [PubMed] [Google Scholar]
5. Bullier J, Henry GH. Ламинарное распределение нейронов первого порядка и афферентных окончаний в стриарной коре кошек. J Нейрофизиол. 1979; 42: 1271–1281. [PubMed] [Google Scholar]
J Нейрофизиол. 1979; 42: 1271–1281. [PubMed] [Google Scholar]
6. Bullier J, Kennedy H, Salinger W. Бифуркация подкорковых афферентов к зрительным областям 17, 18 и 19в коре кошек. J Комп Нейрол. 1984; 228: 309–328. [PubMed] [Google Scholar]
7. Burkhalter A. Внутренние связи первичной зрительной коры крысы: ламинарная организация проекций аксонов. J Комп Нейрол. 1989; 279: 171–186. [PubMed] [Google Scholar]
8. Callaway E. Локальные схемы в первичной зрительной коре макаки. Annu Rev Neurosci. 1998; 21:47–74. [PubMed] [Google Scholar]
9. Касагранде В.А. Третий параллельный зрительный путь к области приматов V1. Тренды Нейроси. 1994;17:305–310. [PubMed] [Google Scholar]
10. Конли М., Рачковски Д. Субламинарная организация в слое VI полосатой коры у галаго. J Комп Нейрол. 1990; 302: 425–436. [PubMed] [Google Scholar]
11. Эдвардс Ф.А., Коннерт Т. Пэтч-фиксация клеток в препаратах срезов тканей. Методы Энзимол. 1992; 207: 208–222. [PubMed] [Google Scholar]
12.![]() Ферстер Д. Пространственное противопоставление возбуждения и торможения в простых клетках зрительной коры кошки. Дж. Нейроски. 1988; 8: 1172–1180. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ферстер Д. Пространственное противопоставление возбуждения и торможения в простых клетках зрительной коры кошки. Дж. Нейроски. 1988; 8: 1172–1180. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Ferster D. X- и Y-опосредованные синаптические потенциалы в нейронах 17 и 18 областей зрительной коры кошки. Vis Neurosci. 1990; 4: 115–133. [PubMed] [Google Scholar]
14. Ferster D, Lindström S. Внутриклеточный анализ геникуло-кортикальной связи в области 17 кошки. J Physiol (Лондон) 1983; 342: 181–215. [PMC free article] [PubMed] [Google Scholar]
15. Ferster D, Lindström S. Синаптическое возбуждение нейронов в области 17 кошки внутрикортикальными коллатералями аксонов кортико-геникулярных клеток. J Physiol (Лонд) 1985;367:233–252. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Фитцпатрик Д. Функциональная организация локальных цепей в зрительной коре: результаты исследования полосатой коры землеройки. Кора головного мозга. 1996; 6: 329–341. [PubMed] [Google Scholar]
17. Физтпатрик Д., Лунд Дж. С., Бласдел Г. Г. Внутренние связи в полосатой коре макака: афферентные и эфферентные связи пластинки 4C. Дж. Нейроски. 1985; 5: 3329–3349. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Физтпатрик Д., Лунд Дж. С., Бласдел Г. Г. Внутренние связи в полосатой коре макака: афферентные и эфферентные связи пластинки 4C. Дж. Нейроски. 1985; 5: 3329–3349. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Фитцпатрик Д., Усри В.М., Шофилд Б.Р., Эйнштейн Г. Субламинарная организация кортикогенных нейронов в слое 6 полосатой коры макака. Vis Neurosci. 1994;11:307–315. [PubMed] [Google Scholar]
19. Гилберт CD. Ламинарные различия в свойствах рецептивного поля клеток первичной зрительной коры кошек. J Physiol (Лондон) 1977; 268: 391–421. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Gilbert CD, Kelly JP. Проекции клеток разных слоев зрительной коры. J Комп Нейрол. 1975; 163: 81–106. [PubMed] [Google Scholar]
21. Gilbert CD, Wiesel TN. Морфология и интракортикальные проекции функционально идентифицированных нейронов зрительной коры кошек. Природа. 1979;280:120–125. [PubMed] [Google Scholar]
22. Gilbert CD, Wiesel TN. Ламинарная специализация и внутрикорковые связи в первичной зрительной коре кошек. В: Шмитт Ф.О., Уорден Ф.Г., Адельман Г., Деннис С.Г., редакторы. Организация коры головного мозга. Массачусетский технологический институт; Кембридж: 1981. стр. 164–190. [Google Scholar]
В: Шмитт Ф.О., Уорден Ф.Г., Адельман Г., Деннис С.Г., редакторы. Организация коры головного мозга. Массачусетский технологический институт; Кембридж: 1981. стр. 164–190. [Google Scholar]
23. Gilbert CD, Wiesel TN. Кластерные внутренние связи в зрительной коре кошек. Дж. Нейроски. 1983; 3: 1116–1133. [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Gilbert CD, Wiesel TN. Столбчатая специфичность внутренних горизонтальных и корково-кортикальных связей в зрительной коре кошек. Дж. Нейроски. 1989;9:2432–2442. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Grieve KL, Sillito AM. Переоценка роли VI слоя зрительной коры в формировании коркового концевого торможения. Опыт Мозг Res. 1991; 87: 521–529. [PubMed] [Google Scholar]
26. Grieve KL, Sillito AM. Дифференциальные свойства клеток первичной зрительной коры кошек, обеспечивающих кортикофугальную обратную связь с латеральным коленчатым телом и зрительным клауструмом. Дж. Нейроски. 1995; 15:4868–4874. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Хирш Дж.А. Синаптическая интеграция в слое 4 зрительной коры хорька. J Physiol (Лондон) 1995; 481: 247–262. [Google Scholar]
28. Hirsch JA, Alonso JM, Reid RC. Визуально вызванные кальциевые потенциалы действия в стриарной коре кошек. Природа. 1995; 378: 612–616. [PubMed] [Google Scholar]
29. Hirsch JA, Alonso JM, Reid RC, Martinez LM. Различия синаптических ответов сложных клеток первого и второго порядка в стриарной коре кошек. Soc Neurosci Abstr. 1997; 23:1666. [Академия Google]
30. Хоффман К.П., Стоун Дж. Скорость проведения афферентов к зрительной коре головного мозга кошки: корреляция со свойствами коркового рецептивного поля. Мозг Res. 1971; 32: 460–466. [PubMed] [Google Scholar]
31. Horikawa K, Armstrong WE. Универсальный способ мечения: инъекция биоцитина и его детекция конъюгатами авидина. J Neurosci Методы. 1988; 25:1–11. [PubMed] [Google Scholar]
32. Hubel DH, Wiesel TN. Рецептивные поля, бинокулярное взаимодействие и функциональная архитектура зрительной коры кошек. J Physiol (Лонд) 1962;160:106–154. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Рецептивные поля, бинокулярное взаимодействие и функциональная архитектура зрительной коры кошек. J Physiol (Лонд) 1962;160:106–154. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Хамфри А.Л., Сур М., Ульрих Д.Дж., Шерман С.М. Паттерны проекции отдельных аксонов X- и Y-клеток от латерального ядра коленчатого тела к корковым областям 17 и 18. J Comp Neurol. 1985; 233: 159–189. [PubMed] [Google Scholar]
34. Джагадиш Б., Ферстер Д. Длина рецептивного поля в стриарной коре кошек может увеличиваться при уменьшении контраста стимула. Soc Neurosci Abstr. 1990;16:293. [Google Scholar]
35. Джонсон Р. Р., Беркхальтер А. Микросхемы прямых и обратных связей в зрительной коре крысы. J Комп Нейрол. 1996;368:383–98. [PubMed] [Google Scholar]
36. Jones JP, Palmer LA. Двумерная пространственная структура простых рецептивных полей в стриарной коре кошек. J Нейрофизиол. 1987; 58: 1187–1211. [PubMed] [Google Scholar]
37. Katz LC. Локальная схема идентифицированных проекционных нейронов в срезах зрительной коры головного мозга кошки. Дж. Нейроски. 1987; 7: 1223–1249. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Нейроски. 1987; 7: 1223–1249. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. LeVay S, Gilbert CD. Ламинарные паттерны геникулокортикальной проекции у кошек. Мозг Res. 1976;113:1–19. [PubMed] [Google Scholar]
39. LeVay S, Sherk H. Зрительная кора кошки. I. Структура и связи. Дж. Нейроски. 1981; 1: 956–980. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Lin CS, Friedlander MJ, Sherman SM. Морфология физиологически идентифицированных нейронов зрительной коры кошки. Мозг Res. 1979; 172: 344–348. [PubMed] [Google Scholar]
41. Лунд Дж. Анатомическая организация стриарной зрительной коры макак. Annu Rev Neurosci. 1988;11:253–288. [PubMed] [Google Scholar]
42. Лунд Дж. С., Бут Р. Интерламинарные связи и организация пирамидных нейронов в зрительной коре, область 17, обезьяны-макаки. J Комп Нейрол. 1975; 164: 287–303. [PubMed] [Google Scholar]
43. Лунд Дж. С., Лунд Р. Д., Хендриксон А. Е., Бунт А. Х., Фукс А. Ф. Происхождение эфферентных путей из первичной зрительной коры, область 17, макаки, как показано ретроградным транспортом пероксидазы хрена. J Комп Нейрол. 1975; 164: 287–303. [PubMed] [Академия Google]
J Комп Нейрол. 1975; 164: 287–303. [PubMed] [Академия Google]
44. Лунд Дж.С., Генри Г.Х., МакКуин К.Л., Харви А.Р. Анатомическая организация первичной зрительной коры (поле 17) кошки. Сравнение с зоной 17 макаки. J Комп Нейрол. 1979; 184: 599–618. [PubMed] [Google Scholar]
45. Малинов Р., Циен Р.В. Пресинаптическое усиление, показанное цельноклеточными записями долговременной потенциации в срезах гиппокампа. Природа. 1990; 346: 177–180. [PubMed] [Google Scholar]
46. Martin KAC, Whitteridge D. Форма, функция и интракортикальные свойства шипиковых нейронов в стриарной зрительной коре кошки. J Physiol (Лонд) 1984;353:463–504. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. McCourt ME, Boyapati J, Henry GH. Наслоение в пластинке 6 стриарной коры кошки. Мозг Res. 1986; 364: 181–185. [PubMed] [Google Scholar]
48. McGuire B, Hornung JP, Gilbert CD, Wiesel TN. Паттерны синаптического входа 4 слоя стриарной коры кошки. Дж. Нейроски. 1984; 4: 3021–3033. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
49. McGuire BA, Gilbert CD, Rivlin PK, Wiesel TN. Цели горизонтальных связей в первичной зрительной коре макаки. J Комп Нейрол. 1991;305:370–92. [PubMed] [Google Scholar]
50. Миллер Дж.В., Бушманн М.Б., Беневенто Л.А. Внеколенчатые таламические проекции на первичную зрительную кору. Мозг Res. 1980; 189: 221–227. [PubMed] [Google Scholar]
51. Мовшон Дж.А., Томпсон И.Д., Толхерст Д.Дж. Организация рецептивного поля сложных клеток в стриарной коре кошек. J Physiol (Лондон) 1978; 283: 79–99. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Murphy PC, Sillito AM. Функциональная морфология пути обратной связи от поля 17 зрительной коры кошки к латеральному коленчатому телу. Дж. Нейроски. 1996;16:1180–1192. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Ohzawa I, Freeman RD. Бинокулярная организация сложных клеток зрительной коры кошек. J Нейрофизиол. 1986; 56: 243–259. [PubMed] [Google Scholar]
54. Палмер Л.А., Дэвис Т.Л. Сравнение ответов на движущиеся и неподвижные стимулы в стриарной коре кошек. J Нейрофизиол. 1981; 46: 260–276. [PubMed] [Google Scholar]
Палмер Л.А., Дэвис Т.Л. Сравнение ответов на движущиеся и неподвижные стимулы в стриарной коре кошек. J Нейрофизиол. 1981; 46: 260–276. [PubMed] [Google Scholar]
55. Робсон Дж.А. Морфология кортикофугальных аксонов в дорсально-латеральном коленчатом теле у кошки. J Комп Нейрол. 1983;216:89–104. [PubMed] [Google Scholar]
56. Sawatari A, Callaway EM. Конвергенция магно- и парвоцеллюлярных путей в слое 4В первичной зрительной коры макака. Природа. 1996; 380:442–446. [PubMed] [Google Scholar]
57. Скоттун Б.С., ДеВалуа Р.Л., Грософ Д.Х., Мовшон Дж.А., Альбрехт Д.Г., Бондс А.Б. Классификация простых и сложных клеток на основе модуляции ответа. Видение Рез. 1991; 31: 1079–1086. [PubMed] [Google Scholar]
58. Somogyi P. Синаптическая организация ГАМКергических нейронов и ГАМК a рецепторы латерального коленчатого тела и зрительной коры. В: Lam DM, Gilbert CD, редакторы. Нейронные механизмы восприятия. Залив; Хьюстон: 1989. стр. 35–62. [Google Scholar]
59. Штюмер В., Робертс В.М., Алмерс В. Свободный пластырь. В: Сакманн Б., Неер Э., редакторы. Одноканальная запись. пленум; Нью-Йорк: 1983. С. 123–132. [Google Scholar]
Штюмер В., Робертс В.М., Алмерс В. Свободный пластырь. В: Сакманн Б., Неер Э., редакторы. Одноканальная запись. пленум; Нью-Йорк: 1983. С. 123–132. [Google Scholar]
60. Usrey WM, Fitzpatrick D. Специфичность аксональных соединений нейронов слоя VI в полосатой коре землеройки: свидетельство существования различных гранулярных и супрагранулярных систем. Дж. Нейроски. 1996;16:1203–1218. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Wiser AK, Callaway EM. Вклад отдельных пирамидных нейронов слоя 6 в локальные схемы первичной зрительной коры макака. Дж. Нейроски. 1996; 15: 2724–2739. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Сложные тепловые карты выявляют закономерности и корреляции в многомерных геномных данных | Биоинформатика
Журнальная статья
Цзугуан Гу,
Цзугуан Гу
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Роланд Эйлс,
Роланд Эйлс
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Матиас Шлеснер
Матиас Шлеснер *
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Примечания автора
Биоинформатика , том 32, выпуск 18, 15 сентября 2016 г. , страницы 2847–2849, https://doi.org/10.1093/bioinformatics/btw313
, страницы 2847–2849, https://doi.org/10.1093/bioinformatics/btw313
Опубликовано:
20 мая 2016 г.
История статьи
Получен:
27 декабря 2015 г.
Полученная ревизия:
12 мая 2016 г.
Принято:
13 мая 2016
Опубликовано:
20 мая 2016
- Разделенный вид
- Содержание статьи
- Рисунки и таблицы
- видео
- Аудио
- Дополнительные данные
Цитировать
Cite
Zuguang Gu, Roland Eils, Matthias Schlesner, Сложные тепловые карты выявляют закономерности и корреляции в многомерных геномных данных, Bioinformatics , Volume 32, Issue 18, 15 September 2016, Pages 2847–2849, https://doi.
 org/10.1093/биоинформатика/btw313
org/10.1093/биоинформатика/btw313Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Разрешения
- Электронная почта
- Твиттер
- Фейсбук
- Подробнее
Фильтр поиска панели навигации БиоинформатикаЭтот выпускЖурналы по биоинформатикеБиоинформатика и вычислительная биологияКнигиЖурналыOxford Academic Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации БиоинформатикаЭтот выпускЖурналы по биоинформатикеБиоинформатика и вычислительная биологияКнигиЖурналыOxford Academic Термин поиска на микросайте
Advanced Search
Abstract
Резюме: Параллельные тепловые карты с тщательно разработанной аннотационной графикой являются мощным инструментом для эффективной визуализации паттернов и взаимосвязей между многомерными геномными данными. Здесь мы представляем Пакет ComplexHeatmap , предоставляющий широкие функциональные возможности для настройки тепловых карт, организации нескольких параллельных тепловых карт и включения графических аннотаций, определяемых пользователем. Мы демонстрируем возможности ComplexHeatmap для простого выявления закономерностей и корреляций между несколькими источниками информации с четырьмя наборами реальных данных.
Здесь мы представляем Пакет ComplexHeatmap , предоставляющий широкие функциональные возможности для настройки тепловых карт, организации нескольких параллельных тепловых карт и включения графических аннотаций, определяемых пользователем. Мы демонстрируем возможности ComplexHeatmap для простого выявления закономерностей и корреляций между несколькими источниками информации с четырьмя наборами реальных данных.
Доступность и реализация: Пакет ComplexHeatmap и документация находятся в свободном доступе в проекте Bioconductor: http://www.bioconductor.org/packages/devel/bioc/html/ComplexHeatmap.html.
Контактное лицо: [email protected]
Дополнительная информация: Дополнительные данные доступны по адресу Биоинформатика онлайн.
1 Введение
Тепловые карты — это фундаментальный метод визуализации, который широко используется для выявления закономерностей, скрытых в геномных данных. Они особенно популярны для анализа экспрессии генов (Eisen et al. , 1998) и профилирования метилирования (Sturm et al. , 2012). С ростом доступности наборов геномных данных срочно необходимы методы визуализации, которые эффективно отображают отношения в многомерных данных. В этой статье мы показываем, как тепловые карты с тщательно разработанной графикой аннотаций могут значительно улучшить раскрытие базовой структуры данных, и как использование параллельных тепловых карт, ориентированных на разные источники информации, дает быстрый и всесторонний обзор данных.
Они особенно популярны для анализа экспрессии генов (Eisen et al. , 1998) и профилирования метилирования (Sturm et al. , 2012). С ростом доступности наборов геномных данных срочно необходимы методы визуализации, которые эффективно отображают отношения в многомерных данных. В этой статье мы показываем, как тепловые карты с тщательно разработанной графикой аннотаций могут значительно улучшить раскрытие базовой структуры данных, и как использование параллельных тепловых карт, ориентированных на разные источники информации, дает быстрый и всесторонний обзор данных.
В среде программирования R традиционные инструменты для рисования тепловых карт, такие как базовая функция тепловой карты или дополнительные пакеты, такие как тепловая карта или тепловая карта плюс , обеспечивают только ограниченную функциональность для отображения графических аннотаций и не поддерживают построение графиков. несколько параллельных тепловых карт. Пакет ComplexHeatmap был разработан для преодоления этих ограничений.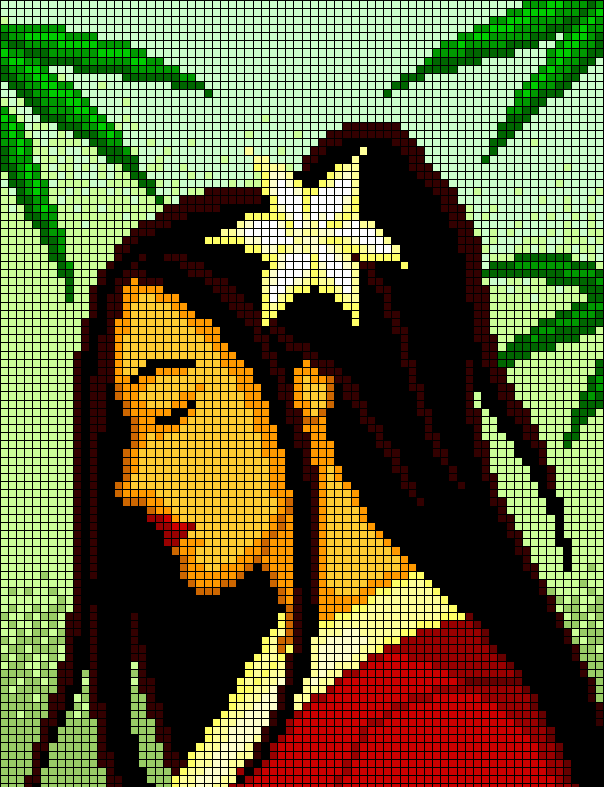 Он обеспечивает общее решение для сопоставления различных источников информации в нескольких параллельных тепловых картах. Каждая тепловая карта может быть дополнена несколькими графическими аннотациями, чтобы завершить отображение набора данных. Графические аннотации столбцов и строк поддерживаются в версии 9.0169 ComplexHeatmap может быть предопределенной графикой, например. точек, гистограмм или коробчатых диаграмм или более общей определяемой пользователем графики. Другие функции ComplexHeatmap включают: (i) гибкую поддержку кластеризации. Например, поддерживаются визуализированные дендрограммы (Galili, 2015) или определяемая пользователем функция расстояния, которая принимает два парных вектора; (ii) разделение строк тепловой карты на срезы для поддержки визуализации подгрупп, где разделение строк может быть выполнено либо методом разделения, например k — означает кластеризацию или фрейм данных, содержащий классификации; (iii) пользовательская настройка сетки тепловой карты для более продвинутой визуализации сложной информации, например.
Он обеспечивает общее решение для сопоставления различных источников информации в нескольких параллельных тепловых картах. Каждая тепловая карта может быть дополнена несколькими графическими аннотациями, чтобы завершить отображение набора данных. Графические аннотации столбцов и строк поддерживаются в версии 9.0169 ComplexHeatmap может быть предопределенной графикой, например. точек, гистограмм или коробчатых диаграмм или более общей определяемой пользователем графики. Другие функции ComplexHeatmap включают: (i) гибкую поддержку кластеризации. Например, поддерживаются визуализированные дендрограммы (Galili, 2015) или определяемая пользователем функция расстояния, которая принимает два парных вектора; (ii) разделение строк тепловой карты на срезы для поддержки визуализации подгрупп, где разделение строк может быть выполнено либо методом разделения, например k — означает кластеризацию или фрейм данных, содержащий классификации; (iii) пользовательская настройка сетки тепловой карты для более продвинутой визуализации сложной информации, например.![]() усовершенствованный OncoPrint (Gao et al. , 2013 г.) на рисунке 1A; (iv) интерактивный выбор на тепловых картах для получения подмножества строк и столбцов, если тепловые карты рисуются на интерактивном устройстве (например, X11) и (v) возможность добавлять более настраиваемую графику после создания тепловых карт.
усовершенствованный OncoPrint (Gao et al. , 2013 г.) на рисунке 1A; (iv) интерактивный выбор на тепловых картах для получения подмножества строк и столбцов, если тепловые карты рисуются на интерактивном устройстве (например, X11) и (v) возможность добавлять более настраиваемую графику после создания тепловых карт.
Рис. 1.
Открыть в новой вкладкеСкачать слайд
Примеры от ComplexHeatmap . ( A ) Геномные изменения и биологические функции ассоциированных мутировавших генов. ( B ) Гетерогенность мышиных Т-клеток по данным одноклеточной РНК-Seq. Источники данных и код R для графиков можно найти в дополнительных S1 и S2
ComplexHeatmap имеет модульную конструкцию с удобным интерфейсом прикладного программирования (API). Гибкость и расширяемость пакета позволяют быстро и легко создавать новые представления многомерных наборов данных и значительно облегчают открытия из геномных и других омических данных.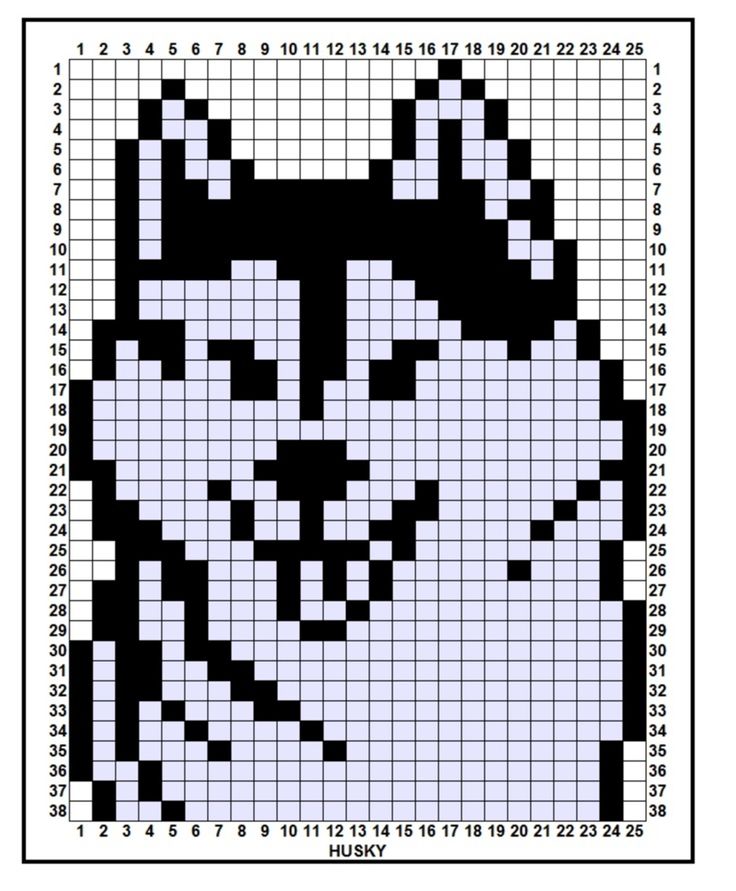
2 Реализация
Комплексная тепловая карта реализована объектно-ориентированным способом. Основные классы:
Тепловая карта : Представление одной тепловой карты. Класс обрабатывает матрицу данных и предоставляет методы для рисования компонентов тепловой карты, то есть тела тепловой карты, имен строк/столбцов, заголовков, дендрограмм и аннотаций столбцов.
HeatmapList : Представление списка тепловых карт. Класс настраивает графические параметры для списка тепловых карт, создает макет и предоставляет методы для рисования таких компонентов, как легенды тепловых карт. Оператор «+» используется для объединения параллельных тепловых карт (т. е. тепловых карт, в которых соответствующие строки во всех матрицах соответствуют одному и тому же объекту):
Тепловая карта(матрица1, …) + Тепловая карта(матрица2, …)
Тепловая картаАннотация: Аннотация тепловой карты — это общая и гибкая концепция.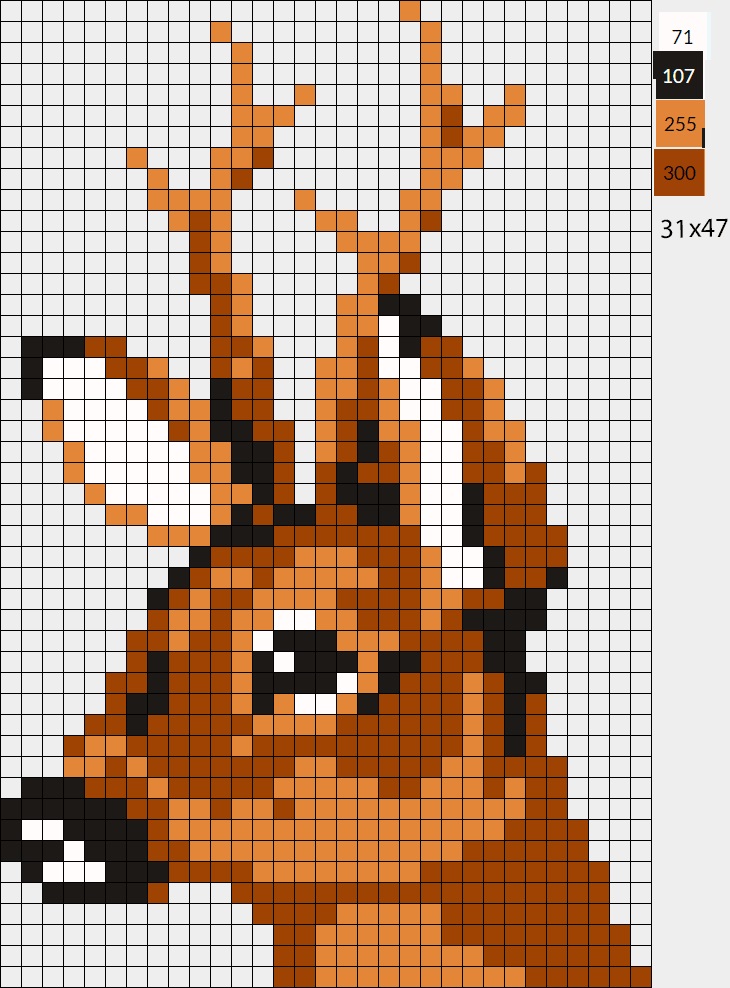 Единственным требованием к аннотациям тепловой карты является то, что графика должна быть выровнена по столбцам или строкам тепловой карты соответственно. Наиболее часто используемые графические аннотации представляют собой список сеток, которые показывают различные группы данных. Однако могут использоваться и другие типы графики, например. это могут быть диаграммы, которые визуализируют распределение данных в соответствующих строках или столбцах. ComplexHeatmap предоставляет несколько фиксированных типов графических аннотаций, например. точек, гистограмм и коробчатых диаграмм. Пакет также предоставляет API, который позволяет пользователям создавать собственные графические аннотации.
Единственным требованием к аннотациям тепловой карты является то, что графика должна быть выровнена по столбцам или строкам тепловой карты соответственно. Наиболее часто используемые графические аннотации представляют собой список сеток, которые показывают различные группы данных. Однако могут использоваться и другие типы графики, например. это могут быть диаграммы, которые визуализируют распределение данных в соответствующих строках или столбцах. ComplexHeatmap предоставляет несколько фиксированных типов графических аннотаций, например. точек, гистограмм и коробчатых диаграмм. Пакет также предоставляет API, который позволяет пользователям создавать собственные графические аннотации.
Аннотации столбцов и аннотаций строк инкапсулируются классом HeatmapAnnotation . Аннотации столбцов являются компонентами одной тепловой карты:
ha = HeatmapAnnotation(…)
Heatmap(matrix, top_annotation = ha, …)
Аннотации строк присоединяются к списку тепловой карты оператором «+» и могут соответствовать к нескольким параллельным тепловым картам:
га = Тепловая картаАннотация(…, которая = ‘строка’)
Тепловая карта(матрица, …) + га
Количество и порядок аннотаций тепловой карты в списке тепловых карт не ограничены.
3 Заявка
На рисунке 1A визуализируются геномные изменения для 38 выбранных генов у 134 пациентов из когорты аденокарциномы легкого TCGA (Collisson et al. , 2014) в виде улучшенного OncoPrint. Гены разделены на две группы в зависимости от скорости амплификации среди пациентов. Гистограммы добавляются к строкам и столбцам, чтобы показать количество изменений у пациентов и генов соответственно. Дополнительная тепловая карта справа указывает на биологические функции мутировавших генов. Комбинация OncoPrint и этой тепловой карты показывает, что сильно амплифицированные гены относительно активны в процессах клеточного ответа, в то время как мутированные гены обогащены процессами, связанными с развитием и метаболизмом.
На рисунке 1B показана гетерогенность мышиных Т-клеток, проанализированная с помощью данных РНК-Seq отдельных клеток (Buettner et al. , 2015). Профили экспрессии 721 выбранного гена классифицируют клетки на две субпопуляции. Левая субпопуляция (выделена светло-красным цветом в столбце дендрограммы) характеризуется относительно высокой экспрессией подмножества генов клеточного цикла, однако абсолютный уровень экспрессии низкий. Субпопуляция справа демонстрирует относительно низкую экспрессию этого подмножества генов клеточного цикла, в то время как другие выбранные гены, включая другие гены клеточного цикла, экспрессируются на высоких уровнях. Группа генов рибонуклеопротеинов демонстрирует очень сильную коэкспрессию. Указаны названия генов высокоэкспрессированных генов клеточного цикла. Тепловая карта справа визуализирует парные корреляции между 721 геном. Значения корреляции были иерархически сгруппированы, и полученный порядок строк использовался для определения порядка строк всех параллельных тепловых карт.
Левая субпопуляция (выделена светло-красным цветом в столбце дендрограммы) характеризуется относительно высокой экспрессией подмножества генов клеточного цикла, однако абсолютный уровень экспрессии низкий. Субпопуляция справа демонстрирует относительно низкую экспрессию этого подмножества генов клеточного цикла, в то время как другие выбранные гены, включая другие гены клеточного цикла, экспрессируются на высоких уровнях. Группа генов рибонуклеопротеинов демонстрирует очень сильную коэкспрессию. Указаны названия генов высокоэкспрессированных генов клеточного цикла. Тепловая карта справа визуализирует парные корреляции между 721 геном. Значения корреляции были иерархически сгруппированы, и полученный порядок строк использовался для определения порядка строк всех параллельных тепловых карт.
Дополнительный файл S3 визуализирует корреляции между метилированием, экспрессией генов, энхансерами и информацией, связанной с генами. Данные генерируются случайным образом на основе шаблонов, найденных в неопубликованной работе. На тепловых картах каждая строка соответствует области дифференциального метилирования (DMR) или другим объектам, связанным с DMR. Этими объектами являются, например, ближайший ген с экспрессией, отрицательно коррелирующей с уровнем метилирования в ассоциированном DMR, или энхансеры, перекрывающиеся с DMR. Сложные тепловые карты показывают, что сильно метилированные DMR обогащены межгенными и внутригенными областями и редко перекрываются с энхансерами. Напротив, низкометилированные DMR обогащены сайтами начала транскрипции (TSS) и энхансерами.
На тепловых картах каждая строка соответствует области дифференциального метилирования (DMR) или другим объектам, связанным с DMR. Этими объектами являются, например, ближайший ген с экспрессией, отрицательно коррелирующей с уровнем метилирования в ассоциированном DMR, или энхансеры, перекрывающиеся с DMR. Сложные тепловые карты показывают, что сильно метилированные DMR обогащены межгенными и внутригенными областями и редко перекрываются с энхансерами. Напротив, низкометилированные DMR обогащены сайтами начала транскрипции (TSS) и энхансерами.
Дополнительный файл S4 повторно реализует рисунок 1 в Sturm et al. (2012 г.), чтобы продемонстрировать способность ComplexHeamtap создавать сложные аннотации. По сравнению с исходным рисунком добавлены две новые тепловые карты: одна визуализирует расстояние между сайтами CpG и ближайшим TSS, а вторая визуализирует аннотации к островам CpG (CGI). Тепловая карта разделена в соответствии с аннотацией CGI, показывая, что сайты CpG, которые находятся на шельфе CGI и в открытом море, имеют более высокие уровни метилирования и большее расстояние до ближайшего TSS.
Финансирование
Эта работа была поддержана Немецким центром исследования рака-Гейдельбергским центром персонализированной онкологии (DKFZ-HIPO) и сетью de.NBI HD-HuB, финансируемой BMBF (#031A537A, #031A537C).
Конфликт интересов : не объявлено.
Каталожные номера
Бюттнер
F.
и другие. . (
2015
)
Компьютерный анализ межклеточной неоднородности в данных секвенирования РНК одной клетки выявляет скрытые субпопуляции клеток
.
Нац. Биотехнолог
.,
33
,
155
–
160
.
Коллиссон
Е.А.
и другие. . (
. (
2014
)
Комплексное молекулярное профилирование аденокарциномы легкого
.
Природа
,
511
,
543
–
550
.
Эйзен
М.Б.
и другие. . (
1998
)
Кластерный анализ и отображение полногеномных паттернов экспрессии
.
Проц. Натл. акад. Наука
,
95
,
14863
–
14868
.
Галили
Т.
(
2015
)
dendextend: пакет R для визуализации, настройки и сравнения деревьев иерархической кластеризации
.
Биоинформатика
,
31
,
3718
–
3720
и другие. . (
2013
)
Комплексный анализ сложной геномики рака и клинических профилей с использованием cBioPortal
.
Науч. Сигнал
.,
6
,
pl1
.
Штурм
Д.
и другие. . (
2012
)
Точечные мутации в h4F3A и IDh2 определяют отдельные эпигенетические и биологические подгруппы глиобластомы
.
Раковая клетка
,
22
,
425
–
437
.
Примечания автора
Заместитель редактора: Джон Хэнкок
© The Author, 2016. Опубликовано Oxford University Press. Все права защищены. Для разрешений, пожалуйста, по электронной почте: [email protected]
© The Author, 2016. Опубликовано Oxford University Press. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
Раздел выпуска:
АНАЛИЗ ГЕНОМА
Скачать все слайды
Дополнительные данные
Дополнительные данные
Дополнительные данные – zip-файл
Реклама
Цитаты
Просмотры
33 766
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи через
Последний
Самые читаемые
Самые цитируемые
Диагностика подтипа рака легкого с использованием слабопарных мультиомных данных
Воспроизводимое получение, управление и метаанализ (мета)данных о нуклеотидной последовательности с использованием q2-fondue
CGV: средство просмотра генома рака, веб-сервис для комплексного анализа генома рака и фармакогеномных данных
EagleImp: быстрое и точное фазирование и импутация всего генома в одном инструменте
MPVNN: Архитектура видимой нейронной сети с мутированными путями для интерпретируемого прогнозирования риска выживания, связанного с раком
Постоянный помощник/доцент эпидемиологии (2 должности)
Колумбия, Южная Каролина
Отделение инфекционных заболеваний Университета Рочестера Должность преподавателя: клинические испытания вакцины против ВИЧ
Рочестер, Нью-Йорк
Врач-инфекционист
Брисбен, Другое / Не США
ПОСЛЕДОКТОРСКАЯ СТИПЕНДИЯ
Хьюстон, Техас
Просмотреть все вакансии
Реклама
Комплекс туберозного склероза: MedlinePlus Genetics
Описание
Комплекс туберозного склероза — это генетическое заболевание, характеризующееся ростом многочисленных незлокачественных (доброкачественных) опухолей во многих частях тела. Эти опухоли могут возникать в головном мозге, почках, сердце, коже и других органах, что в некоторых случаях приводит к серьезным проблемам со здоровьем. Комплекс туберозного склероза также вызывает проблемы с развитием, а признаки и симптомы заболевания варьируются от человека к человеку.
Эти опухоли могут возникать в головном мозге, почках, сердце, коже и других органах, что в некоторых случаях приводит к серьезным проблемам со здоровьем. Комплекс туберозного склероза также вызывает проблемы с развитием, а признаки и симптомы заболевания варьируются от человека к человеку.
Комплекс туберозного склероза часто поражает головной мозг, при этом у некоторых больных наблюдаются доброкачественные новообразования на внешней поверхности головного мозга (кора головного мозга), известные как кортикальные клубни. У людей с комплексом туберозного склероза часто развивается модель поведения, называемая психоневрологическими расстройствами, связанными с TSC (TAND). Эти расстройства включают гиперактивность, агрессию, психические расстройства, умственную отсталость и проблемы с общением и социальным взаимодействием (расстройство аутистического спектра). Кроме того, у людей с комплексом туберозного склероза может быть синдром дефицита внимания/гиперактивности (СДВГ) или судороги.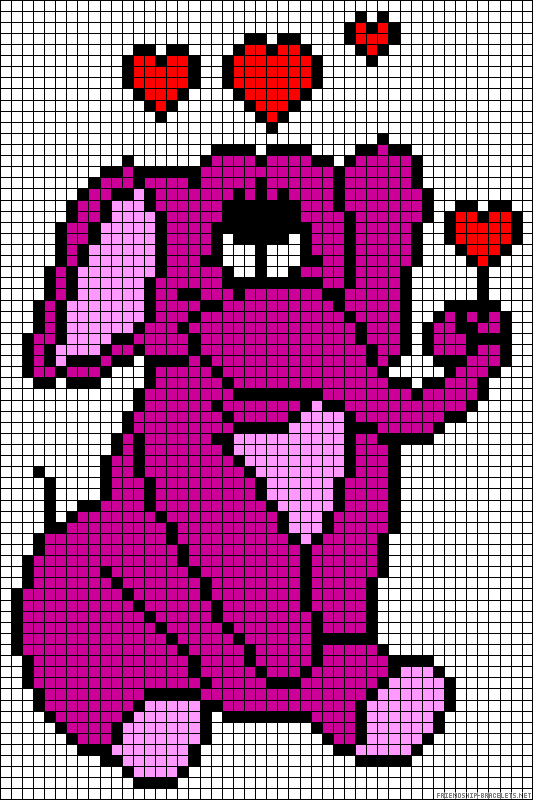
Опухоли почек часто встречаются у людей с комплексом туберозного склероза; эти наросты могут вызвать серьезные проблемы с функцией почек и в некоторых случаях могут быть опасными для жизни. Кроме того, опухоли могут развиваться в сердце (рабдомиома сердца) и в светочувствительной ткани задней части глаза (сетчатке). У некоторых женщин с комплексом туберозного склероза развивается лимфангиолейомиоматоз (ЛАМ), который представляет собой заболевание легких, характеризующееся аномальным разрастанием гладкой мышечной ткани в легких, что вызывает кашель, одышку, боль в груди и коллапс легкого.
Практически все пораженные люди имеют кожные аномалии, в том числе участки необычно светлой кожи, участки приподнятой и утолщенной кожи и наросты под ногтями. Опухоли на лице, называемые ангиофибромами лица, также часто возникают в детстве. Иногда у пораженных людей есть участки повреждения костей или зубов.
Частота
Комплекс туберозного склероза поражает 1 из 6000–10 000 человек.
Причины
Варианты (также известные как мутации) в 9Ген 0169 TSC1 или TSC2 может вызывать комплекс туберозного склероза. Гены TSC1 и TSC2 содержат инструкции по получению белков гамартина и туберина соответственно. Внутри клеток эти два белка работают вместе, помогая регулировать рост и деление клеток (пролиферацию), а также размер клеток. Белки действуют как супрессоры опухолей, которые обычно предотвращают рост и деление клеток слишком быстро или неконтролируемым образом.
Люди с комплексом туберозного склероза рождаются с одной измененной копией гена TSC1 или гена TSC2 в каждой клетке. Вариант в одной копии гена TSC1 не позволяет клеткам вырабатывать функциональный белок гамартин из этой копии; изменение в одной копии гена TSC2 не позволяет клеткам производить функциональный белок туберин из измененной копии. Однако из другой, нормальной копии гена TSC1 или TSC2 обычно вырабатывается достаточное количество белка для эффективного регулирования роста клеток. Чтобы опухоли развивались в комплексе туберозного склероза, второе изменение, затрагивающее другую копию 9Ген 0169 TSC1 или TSC2 должен встречаться в клетках в течение жизни человека.
Чтобы опухоли развивались в комплексе туберозного склероза, второе изменение, затрагивающее другую копию 9Ген 0169 TSC1 или TSC2 должен встречаться в клетках в течение жизни человека.
Клетки, имеющие варианты в обеих копиях гена TSC1 , не могут продуцировать функциональный гамартин; клетки с двумя измененными копиями гена TSC2 неспособны продуцировать какой-либо функциональный туберин. Потеря любого из этих белков позволяет клетке неконтролируемо расти и делиться, образуя опухоль. У людей с комплексом туберозного склероза второй вариант в 9Ген 0169 TSC1 или TSC2 обычно встречается во многих клетках на протяжении жизни больного человека. Отсутствие гамартина или туберина в различных типах клеток приводит к росту опухолей во многих различных органах и тканях, как это наблюдается при туберозном склерозе.
Наследование
Комплекс туберозного склероза имеет аутосомно-доминантный тип наследования, а это значит, что одной копии измененного гена в каждой клетке достаточно для увеличения риска развития опухолей и других проблем с развитием. Примерно в одной трети случаев больной наследует измененный TSC1 или ген TSC2 от родителя, страдающего заболеванием. Остальные две трети людей с комплексом туберозного склероза рождаются с новыми вариантами гена TSC1 или TSC2 . Эти случаи, которые описываются как спорадические, возникают у людей, в семье которых не было туберозного склероза. Варианты гена TSC1 , по-видимому, более распространены в семейных случаях комплекса туберозного склероза, тогда как варианты гена TSC2 чаще встречаются в спорадических случаях.
Примерно в одной трети случаев больной наследует измененный TSC1 или ген TSC2 от родителя, страдающего заболеванием. Остальные две трети людей с комплексом туберозного склероза рождаются с новыми вариантами гена TSC1 или TSC2 . Эти случаи, которые описываются как спорадические, возникают у людей, в семье которых не было туберозного склероза. Варианты гена TSC1 , по-видимому, более распространены в семейных случаях комплекса туберозного склероза, тогда как варианты гена TSC2 чаще встречаются в спорадических случаях.
В редких случаях люди с комплексом туберозного склероза не имеют идентифицированного варианта в гене TSC1 или TSC2 . Исследования показывают, что в этих случаях состояние может быть вызвано случайным вариантом в гене TSC1 или TSC2 , который возникает на очень раннем этапе развития. В результате одни клетки организма имеют нормальную версию гена, а другие — измененную. Такая ситуация называется мозаицизмом.
Такая ситуация называется мозаицизмом.
Другие названия для этого состояния
- Болезнь Бурневиля
- Факоматоз Бурневиля
- Церебральный склероз
- Туберозный склероз
- Туберозный склероз
Дополнительная информация и ресурсы
Информация о генетическом тестировании
- Реестр генетического тестирования: Туберозный склероз 1
- Реестр генетического тестирования: Туберозный склероз 2
Информационный центр генетических и редких заболеваний
- Комплекс туберозного склероза
Ресурсы поддержки и защиты интересов пациентов
- Информационный поиск по болезням
- Национальная организация редких заболеваний (NORD)
Научные исследования от ClinicalTrials.gov
- ClinicalTrials.gov
Каталог генов и болезней от OMIM
- ТУБЕРОЗНЫЙ СКЛЕРОЗ 1
- ТУБЕРОЗНЫЙ СКЛЕРОЗ 2
Научные статьи в PubMed
- PubMed
Ссылки
- Crino PB, Nathanson KL, Henske EP.
 Комплекс туберозного склероза. Н англ J
Мед. 2006 г., 28 сентября; 355(13):1345-56. Обзор. Цитата в PubMed
Комплекс туберозного склероза. Н англ J
Мед. 2006 г., 28 сентября; 355(13):1345-56. Обзор. Цитата в PubMed - Франц Д.Н., Бисслер Дж.Дж., МакКормак FX. Комплекс туберозного склероза: неврологический, почечные и легочные проявления. нейропедиатрия. 2010 Октябрь; 41 (5): 199-208. дои: 10.1055/s-0030-1269906. Epub 2011 5 января. Обзор. Цитирование в PubMed или бесплатная статья в PubMed Central
- Хайман М.Х., Уиттемор В.Х. Консенсусная конференция национальных институтов здравоохранения: комплекс туберозного склероза. Арх Нейрол. 2000 г., май; 57(5):662-5. Обзор. Цитата в PubMed
- Маром Д. Генетика комплекса туберозного склероза: обновление. Чайлдс Нерв Сист. 2020 окт; 36 (10): 2489-2496. doi: 10.1007/s00381-020-04726-z. Epub 2020 6 августа. Обзор. Цитата в PubMed
- Нортруп Х., Аронов М.Э., Бебин Э.М., Бисслер Дж., Дарлинг Т.Н., де Врис П.Дж., Фрост М.Д.,
Фукс З., Госнелл Э.С., Гупта Н.
 , Янсен А.С., Юзвяк С., Кингсвуд Дж.К., Книланс Т.К.,
McCormack FX, Pounders A, Roberds SL, Rodriguez-Buritica DF, Roth J, Sampson JR,
Спарагана С., Тиле Э.А., Вайнер Х.Л., Уилесс Дж.В., Таубин А.Дж., Крюгер Д.А.;
Международная группа консенсуса по комплексу туберозного склероза. Обновленный международный
Комплексные диагностические критерии туберозного склероза, наблюдение и лечение
Рекомендации. Педиатр Нейрол. 2021 окт; 123:50-66. дои:
10.1016/ж.педиатрнейрол.2021.07.011. Epub 2021, 24 июля. Цитирование на PubMed
, Янсен А.С., Юзвяк С., Кингсвуд Дж.К., Книланс Т.К.,
McCormack FX, Pounders A, Roberds SL, Rodriguez-Buritica DF, Roth J, Sampson JR,
Спарагана С., Тиле Э.А., Вайнер Х.Л., Уилесс Дж.В., Таубин А.Дж., Крюгер Д.А.;
Международная группа консенсуса по комплексу туберозного склероза. Обновленный международный
Комплексные диагностические критерии туберозного склероза, наблюдение и лечение
Рекомендации. Педиатр Нейрол. 2021 окт; 123:50-66. дои:
10.1016/ж.педиатрнейрол.2021.07.011. Epub 2021, 24 июля. Цитирование на PubMed - Northrup H, Koenig MK, Pearson DA, Au KS. Комплекс туберозного склероза. 1999 июль 13 [обновлено 9 декабря 2021 г.]. Пришли: Адам М.П., Эверман Д.Б., Мирзаа Г.М., Пагон Р.А., Уоллес SE, Bean LJH, Gripp KW, Amemiya A, редакторы. GeneReviews® [Интернет]. Сиэтл (WA): Вашингтонский университет, Сиэтл; 1993-2022 гг. Доступна с http://www.ncbi.nlm.nih.gov/books/NBK1220/ Цитата в PubMed
- Рэндл СК. Комплекс туберозного склероза: обзор.
 Педиатр Энн. 2017 апр
1;46(4):е166-е171. дои: 10.3928/19382359-20170320-01. Обзор. Цитата в PubMed
Педиатр Энн. 2017 апр
1;46(4):е166-е171. дои: 10.3928/19382359-20170320-01. Обзор. Цитата в PubMed - Salussolia CL, Klonowska K, Kwiatkowski DJ, Sahin M. Генетическая этиология, Диагностика и лечение туберозно-склеротического комплекса. Annu Rev Genomics Hum Жене. 2019 31 августа; 20: 217-240. doi: 10.1146/annurev-genom-083118-015354. Epub 2019 24 апр. Обзор. Цитата в PubMed
Программирование сложных структур тканей в 3D
Одновременное программируемое создание и организация типов клеток различного происхождения и функций в искусственных тканях может вдохнуть новую жизнь в будущую биомедицинскую трехмерную инженерию органов и тканей
Автор Benjamin Boettner
(Бостон) — Тканевые инженеры разработали множество подходов к воссозданию органов и тканей de novo вне человеческого тела для использования в регенеративной терапии, скрининге лекарств и моделировании заболеваний. В качестве двух ярких примеров: человеческие «органоиды» собираются из клеток для формирования крошечных искусственных конструкций в чашке, которые напоминают человеческие органы, и биопринтируются более крупные ткани человека. Оба подхода используют так называемые индуцированные плюрипотентные стволовые клетки (ИПСК) в качестве источника клеток, которые получают из крови, кожи и других клеток, предоставленных донорами-людьми, а затем дифференцируют в типы клеток, типичные для интересующего органа.
Оба подхода используют так называемые индуцированные плюрипотентные стволовые клетки (ИПСК) в качестве источника клеток, которые получают из крови, кожи и других клеток, предоставленных донорами-людьми, а затем дифференцируют в типы клеток, типичные для интересующего органа.
Препятствия на пути тканевой инженерии
На этом изображении показана органоидная структура, полученная из плюрипотентных стволовых клеток человека (iPCS), которые были запрограммированы на создание организованной структуры с сосудистой сетью (красный) и нейронными клетками (синий), подобными тем, которые находятся в мозге. кора. Предоставлено: Институт Висса при Гарвардском университете, С помощью этих методов становится доступным все большее число различных типов клеток и тканей, и исследователи даже начинают решать главную проблему интеграции системы поддержки крови в виде сосудистых сетей. Однако чего им все еще не хватает, так это точного контроля над точным составом типов клеток и их пространственной организацией в функциональные единицы в трехмерных пространствах их конструкций. Кроме того, чтобы перейти к биопроизводству целых органов, эти искусственные тканевые конструкции также должны быть созданы с большей эффективностью, скоростью и масштабируемостью, чем позволяют текущие протоколы.
Кроме того, чтобы перейти к биопроизводству целых органов, эти искусственные тканевые конструкции также должны быть созданы с большей эффективностью, скоростью и масштабируемостью, чем позволяют текущие протоколы.
Теперь совместная многопрофильная исследовательская группа из Института биологического инжиниринга Висса при Гарвардском университете, Гарвардской школы инженерии и прикладных наук им. Джона А. Полсона (SEAS) и Гарвардской медицинской школы (HMS) разработала комплексный подход. для преодоления этих проблем. Путем независимого и одновременного программирования различных популяций иПСК внутри органоидов и 3D-биопечатных тканевых конструкций с факторами транскрипции (TF), определяющими клеточную судьбу, они заставляют их дифференцироваться в желаемые типы клеток, которые могут быть собраны в in vivo -подобные «органотипические» паттерны с беспрецедентным контролем. Их результаты опубликованы в Nature Biomedical Engineering .
«Мы используем генетическое программирование стволовых клеток и органоидов в сочетании с биопечатью, чтобы расширить границы программируемости и многоклеточной организации в трехмерных тканевых конструкциях для моделирования заболеваний и регенеративной медицины», — сказала старший автор и член основного факультета Wyss Дженнифер Льюис, научный сотрудник. .Д. Льюис также является руководителем инициативы Института Wyss по 3D-инженерии органов и Хансйорг Висс Профессор биологической инженерии в SEAS.
.Д. Льюис также является руководителем инициативы Института Wyss по 3D-инженерии органов и Хансйорг Висс Профессор биологической инженерии в SEAS.
Jennifer LewisМы используем генетическое программирование стволовых клеток и органоидов в сочетании с биопечатью, чтобы расширить границы программируемости и многоклеточной организации трехмерных тканевых конструкций для моделирования заболеваний и регенеративной медицины.
Контроль дифференцировки клеток в трехмерных искусственных тканях
Чтобы дифференцировать ИПСК в сторону определенного типа клеток, на них необходимо воздействовать определенными сигналами. Это могут быть химические и биологические факторы, добавленные к среде, в которой культивируются иПСК, подталкивая их к принятию специфической клеточной идентичности. В качестве альтернативы, они снабжают iPCS транскрипционными факторами (TF), инструктирующими судьбу клетки, которые включают программы экспрессии генов, заставляющие iPSC дифференцироваться в сторону определенного типа клеток, минуя химические сигналы в условиях среды.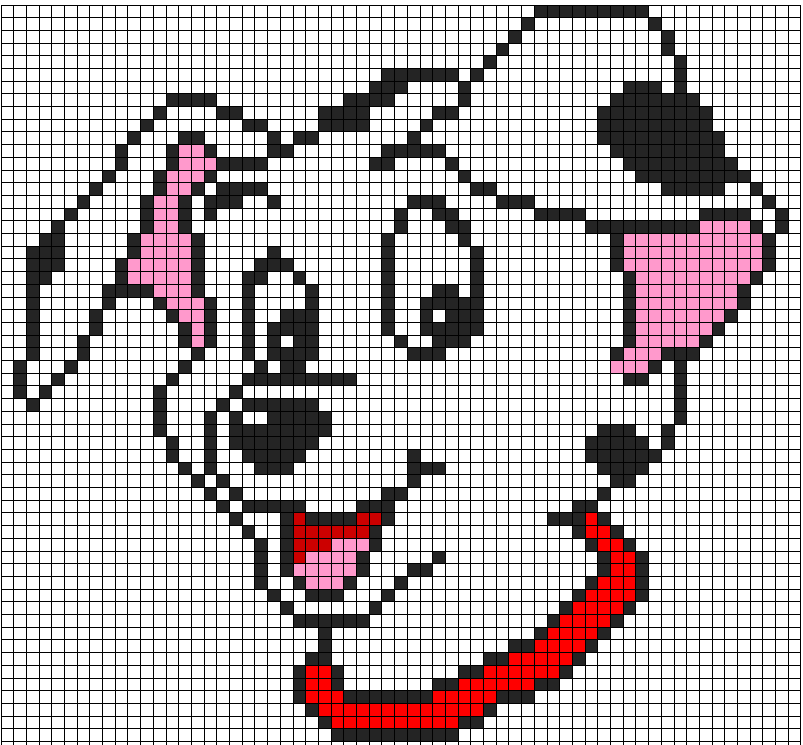 Соавтор Джордж Черч, доктор философии, и его группа собрали наиболее полную коллекцию ТФ, способных генерировать многие типы дифференцированных клеток человека из ИПСК.
Соавтор Джордж Черч, доктор философии, и его группа собрали наиболее полную коллекцию ТФ, способных генерировать многие типы дифференцированных клеток человека из ИПСК.
«Основная проблема создания тканевых конструкций, состоящих из нескольких типов клеток, которые организованы в структурированные структуры, заключалась в том, что среды для культивирования клеток могут определять только генерацию одного или нескольких типов клеток», — заявили co-first и co- автор-корреспондент Марк Скайлар-Скотт, доктор философии. В 2016 году в качестве постдокторского научного сотрудника, работающего с Льюисом, он обсуждал потенциал TF для расширения диапазона и организации клеток в инженерных тканях с Алексом Нг, доктором философии, бывшим постдокторским научным сотрудником в команде Черча Нг, который возглавил открытие TF. усилия, и в настоящее время занимается коммерциализацией технологической платформы на основе TF в качестве CSO GC Therapeutics Inc., соучредителем которой он, как и Черч.
«Мы предположили, что экспрессия ТФ, определяющих тип клеток, может обеспечивать параллельные и независимые программы дифференцировки в клеточных линиях, даже когда они смешаны, и, таким образом, преодолевать ограничивающие условия среды», — сказал Скайлар-Скотт, который сейчас является доцентом биоинженерии в Стэнфорде. Университет.
Университет.
Чтобы подтвердить свою идею, сотрудники смешали и агрегировали ИПСК, которые включают TF ETV2 при добавлении препарата, который индуцирует их дифференцировку в эндотелиальные клетки (клетки iEndo), ИПСК, которые посредством экспрессии TF NGN1 генерируют нейронные клетки ( инейронные клетки) и обычные иПСК, которые в выбранных средах по умолчанию генерируют нейрональные стволовые клетки. Клетки iEndo и iNeuron быстро и более эффективно дифференцировались в эндотелиальные и нейрональные клетки, чем позволяли более ранние методы, и в адаптированных соотношениях. Затем они продолжали формировать интегрированные сосудистые и нейрональные сети, тогда как неиндуцированные iPSCs автономно дифференцировались в маленькие нейросферы в развивающихся органоидах, которые напоминали желудочки мозга.
«Это показало, что разработанный нами метод обеспечивает высокопрограммируемую совместную дифференцировку типов клеток с совершенно разным происхождением в нормальном эмбриогенезе и органогенезе, что в противном случае невозможно легко осуществить в одной тканевой конструкции», — сказал соавтор Джереми Хуанг. , доктор философии, наставником которого в проекте в качестве аспиранта были Льюис и Черч. «Поэтому мы выбрали для него «ортогонально индуцированную дифференциацию» (OID)». Сейчас Хуанг является постдокторантом Массачусетского технологического института.
Рисование паттернов с помощью запрограммированных клеток
Затем команда использовала те же клеточные линии, созданные с помощью OID, для создания тканевых конструкций с паттернами с использованием технологий 3D-органоидов и биопечати. Различные популяции нервных клеток и микрососудистых клеток образуют отдельные слои в областях мозга и вокруг него. Возможность структурировать клетки аналогично in vitro позволила бы исследователям изучить их взаимоотношения крупным планом, например, в случае коры головного мозга и формирования в ней нейронных сетей, миграции клеток между ее различными слоями и специфических для слоев взаимодействий.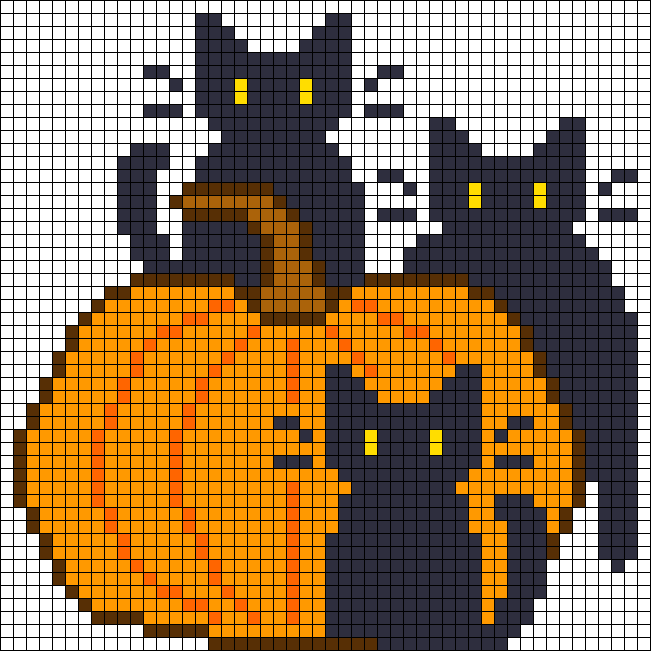 с микроциркуляторным руслом головного мозга. Команда применила метод наслоения, чтобы объединить различные клетки в архитектуру «многоядерная оболочка» с внутренним ядром, содержащим нормальные иПСК, окружающим слоем, содержащим клетки iNeuron, и внешним слоем, содержащим клетки iEndo. При индукции экспрессии различных TF органоиды с многоядерной оболочкой дифференцировались как единое целое в структуру с внутренним «центром зародышевой зоны», в котором находятся и размножаются стволовые клетки, а внешняя зона состоит из дифференцированных нейронов, которые при нормальном развитии мозга происходят из стволовые клетки в зародышевом центре, которые приступили к своей программе дифференцировки и вышли из зародышевого центра, и сосудистая сеть, которая инкапсулирует нейрональный компартмент.
с микроциркуляторным руслом головного мозга. Команда применила метод наслоения, чтобы объединить различные клетки в архитектуру «многоядерная оболочка» с внутренним ядром, содержащим нормальные иПСК, окружающим слоем, содержащим клетки iNeuron, и внешним слоем, содержащим клетки iEndo. При индукции экспрессии различных TF органоиды с многоядерной оболочкой дифференцировались как единое целое в структуру с внутренним «центром зародышевой зоны», в котором находятся и размножаются стволовые клетки, а внешняя зона состоит из дифференцированных нейронов, которые при нормальном развитии мозга происходят из стволовые клетки в зародышевом центре, которые приступили к своей программе дифференцировки и вышли из зародышевого центра, и сосудистая сеть, которая инкапсулирует нейрональный компартмент.
«Важно, что эти структурированные органоиды развивались очень последовательно и с дифференцированными нейронами после восьми дней культивирования, в то время как мозговым органоидам обычно требуется несколько недель культивирования для образования нейронов. Анализируя отдельные клетки в органоидах на экспрессию их генов, мы подтвердили, что они следуют независимым траекториям, не мешая друг другу. Этот уровень скорости и надежности предлагает огромные преимущества для будущих биомедицинских подходов. Программируя клеточную идентичность и распределение в органоидах мозга, мы теперь можем контролировать состав органоидов способами, которые раньше были невозможны», — сказал соавтор Арик Лу, аспирант, работающий с Льюисом.
Анализируя отдельные клетки в органоидах на экспрессию их генов, мы подтвердили, что они следуют независимым траекториям, не мешая друг другу. Этот уровень скорости и надежности предлагает огромные преимущества для будущих биомедицинских подходов. Программируя клеточную идентичность и распределение в органоидах мозга, мы теперь можем контролировать состав органоидов способами, которые раньше были невозможны», — сказал соавтор Арик Лу, аспирант, работающий с Льюисом.
Наконец, команда разработала биологические чернила на основе своей системы OID для 3D-биопечати. Поскольку нормальные иПСК и их аналоги, сконструированные для экспрессии индуцируемых лекарственным средством ТФ, но еще не индуцированные лекарственным средством, значительно размножаются в клеточной культуре, можно легко получить достаточное количество клеточного материала для крупномасштабных приложений биопечати. В частности, команда создала биочернила, содержащие концентрированные нормальные клетки iPSC, iEndo и iNeuron, которые они заполнили тремя независимыми каналами доставки печатающей головки. После укладки узора из параллельных полос сформировалась непрерывная нить, в которой исследователи затем индуцировали экспрессию TF. Через шесть дней иПСК приобрели идентичность нейрональных стволовых клеток. Клетки iEndo продолжали дифференцироваться в эндотелиальные клетки, которые образовывали микрососудистую сеть, а клетки iNeuron дифференцировались в нейроны, демонстрируя, что эти клеточные линии могут использоваться вместе для создания структурированных многоклеточных структур, которые содержат аспекты развивающегося мозга.
Поскольку нормальные иПСК и их аналоги, сконструированные для экспрессии индуцируемых лекарственным средством ТФ, но еще не индуцированные лекарственным средством, значительно размножаются в клеточной культуре, можно легко получить достаточное количество клеточного материала для крупномасштабных приложений биопечати. В частности, команда создала биочернила, содержащие концентрированные нормальные клетки iPSC, iEndo и iNeuron, которые они заполнили тремя независимыми каналами доставки печатающей головки. После укладки узора из параллельных полос сформировалась непрерывная нить, в которой исследователи затем индуцировали экспрессию TF. Через шесть дней иПСК приобрели идентичность нейрональных стволовых клеток. Клетки iEndo продолжали дифференцироваться в эндотелиальные клетки, которые образовывали микрососудистую сеть, а клетки iNeuron дифференцировались в нейроны, демонстрируя, что эти клеточные линии могут использоваться вместе для создания структурированных многоклеточных структур, которые содержат аспекты развивающегося мозга.
Джордж ЧерчЭто важный шаг на пути к моделированию определенных областей мозга, поскольку обнаруживается больше TF, которые могут помочь нам в дальнейшем определить субидентичность нейронов в этих слоях. Тем не менее […] наш подход предлагает потенциал для конструирования гораздо большего количества тканей и органов человека, просто беря страницы из учебника по развитию и используя инженерию генома и другие инструменты, которые мы разрабатываем.
«Это важный шаг на пути к моделированию конкретных областей мозга, поскольку обнаруживается все больше TF, которые могут помочь нам в дальнейшем определить субидентичность нейронов в этих слоях», — сказал Черч. «Однако, хотя мы начали наш проект OID, сосредоточив внимание на нейронных клетках, поскольку к ним относительно легко получить доступ с помощью TF, наш подход предлагает потенциал для конструирования гораздо большего количества тканей и органов человека, просто беря страницы из учебника по развитию и используя инженерию генома и другие инструменты. которые мы развиваем». Черч является руководителем платформы синтетической биологии Института Висса, а также профессором генетики в HMS, а также профессором технологий медицинских наук в Гарварде и Массачусетском технологическом институте.
которые мы развиваем». Черч является руководителем платформы синтетической биологии Института Висса, а также профессором генетики в HMS, а также профессором технологий медицинских наук в Гарварде и Массачусетском технологическом институте.
Дополнительными авторами исследования являются Томоя Дуенки, бывший аспирант группы Льюиса, который сыграл важную роль в анализе системы OID, многоядерной оболочки и анализе биопечати, а также Сонглей Лю, Люси Нам и Сарита Дамараджу. Исследование проводилось за счет грантов Национального института исследований генома человека Национального института здравоохранения (NIH) (№ RM1HG008525), Инициативы NIH Brain (№ RO1Mh223977-01), Программы стипендий факультета Ванневара Буша, спонсируемой Управлением военно-морских исследований ( #N00014-16-1-2823 и N00014-21-1-2958).
наведение моста от большого к малому
Сложные узоры: наведение моста от большого к малому 18 августа 2022 г. | |
| ( Nanowerk News ) Для многих важных для жизни процессов, таких как клеточное деление, клеточная миграция и развитие органов, необходимо пространственно и временно правильное формирование биологических паттернов. | |
| Для понимания этих процессов основная задача состоит не в объяснении того, как из однородного начального состояния формируются закономерности, а в объяснении того, как простые закономерности превращаются во все более сложные. | |
| Выяснение механизмов этой сложной самоорганизации в различных пространственных и временных масштабах является ключевой задачей науки. Так называемые методы «крупнозернистой детализации» позволяют упростить такие многомасштабные системы, чтобы их можно было описать с помощью редуцированной модели в больших масштабах длины и времени. | |
Нелинейная динамика между белками MinE и MinD приводит к образованию сложных узоров вдоль плоской поверхности мембраны. (Изображение: Л. Вюртнер) (Изображение: Л. Вюртнер) | |
| «Однако цена, которую вы платите за крупнозернистость, заключается в том, что важная информация о паттернах на малых масштабах — например, о типе паттерна — теряется. Но дело в том, что эти закономерности играют решающую роль в биологических системах. Например, они контролируют важные клеточные процессы», — объясняет Лаэшкир Вюртнер, член группы под руководством физика LMU профессора Эрвина Фрея и ведущий автор нового исследования ( PNAS , «Соединительные чешуйки в многомасштабной системе формирования паттернов». “), который решает эту проблему. | |
В сотрудничестве с исследовательской группой профессора Сеса Деккера (Технический университет Делфта) команда Фрея разработала новый подход к крупномасштабному анализу так называемых систем реакции-диффузии с сохранением массы, в котором крупномасштабный анализ полных плотностей вовлеченные частицы позволяют предсказывать закономерности в малых масштабах.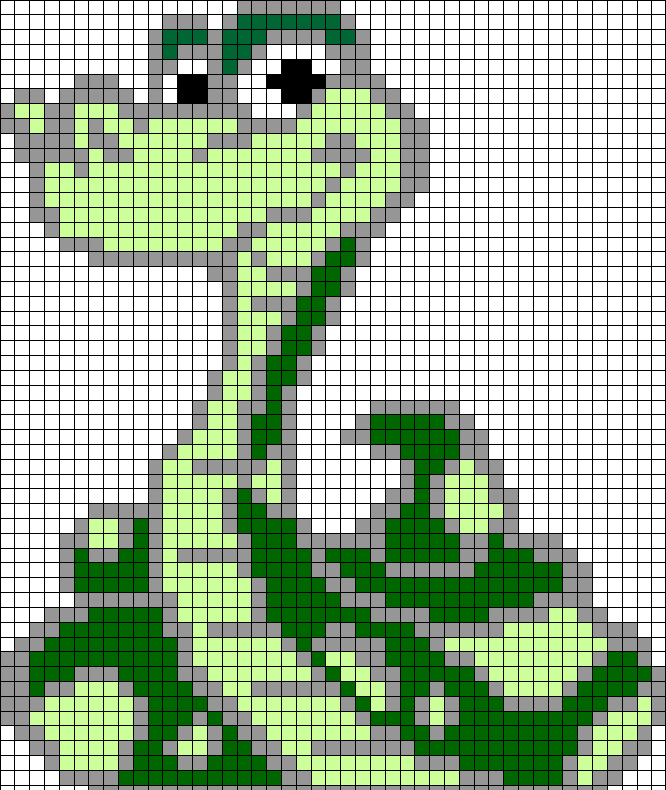 | |
Числовые вычисления в минутах вместо месяцев | |
| Ученые проиллюстрировали потенциал своего подхода с помощью белковой системы Min, парадигмальной модели формирования биологического паттерна. Бактерия E. coli использует различные белки Min, циркулирующие в клетке, чтобы определить, в каком месте происходит деление клетки. Решающим фактором здесь является то, что задействованные белки встречаются с разной частотой в зависимости от их местоположения в клетке и химического состояния, то есть они имеют разную плотность. | |
| «Теперь нам удалось уменьшить сложность этой системы, разработав теорию, основанную исключительно на общей плотности белков, так что мы можем полностью отразить динамику формирования паттерна», — говорит Фрей. «Это огромное сокращение. Численные расчеты теперь выполняются за минуты, а не за месяцы». | |
Исследователям удалось экспериментально подтвердить теоретические предсказания модели, согласно которой распределение белков зависит от геометрии среды.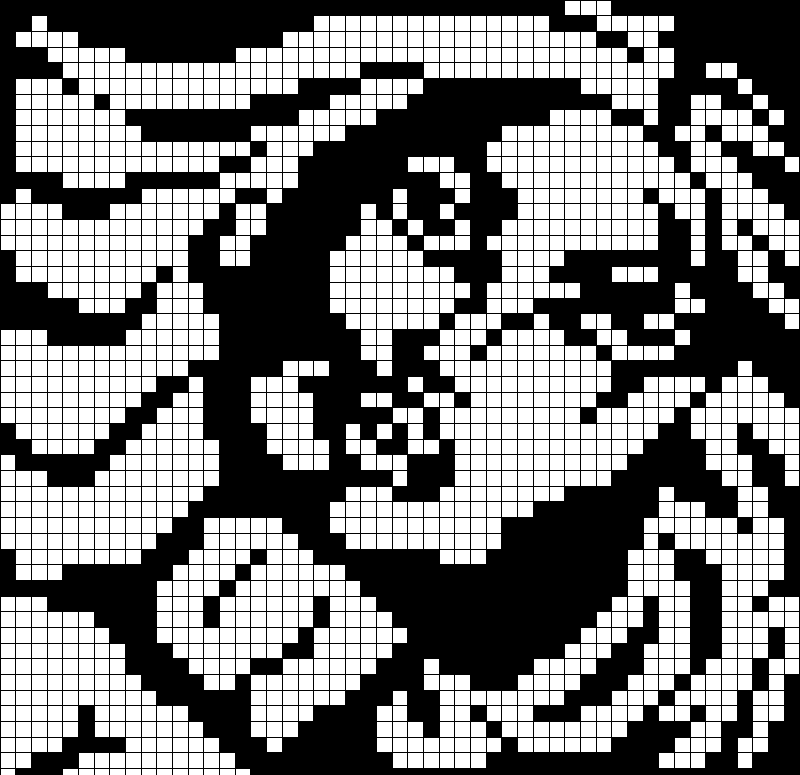 Они сделали это, реконструировав белковую систему Min в проточной ячейке in vitro, и результаты показали те же белковые структуры, которые были выявлены при моделировании. Они сделали это, реконструировав белковую систему Min в проточной ячейке in vitro, и результаты показали те же белковые структуры, которые были выявлены при моделировании. | |
| «Такая реконструкция информации в малом масштабе из уменьшенной динамики на макроскопическом уровне открывает новые пути для лучшего понимания сложных многомасштабных систем, которые встречаются в широком диапазоне физических систем», — говорит Фрей. |
| Источник: Ludwig-Maximilians-Universität München | |
| Поделись этим: | |
Информационный бюллетень Nanowerk
Получайте наши новости о биотехнологии на свой почтовый ящик!
Новости биотехнологии
Исследователи продвигают усилия по адаптации доставки лекарств к «электростанциям» клеток
20 сентября 2022 г.
Ученые внедряют в клетки «неканонический» путь для производства собственных лекарств
19 сентября 2022 г.
Beyond AlphaFold: AI. превосходен в создании новых белков
15 сентября 2022 г.
Новаторские исследования с использованием бактерий приближают ученых к созданию искусственных клеток с функциями, приближенными к живым
15.09.2022
Диагностика рака по миллилитру крови
10 сентября 2022 г.
Движение ДНК, связанное с ее реакцией на повреждение, способность к самовосстановлению
03 сентября 2022 г.
Новое исследование подтверждает предсказанную в 1953 году структуру белка «рифленый лист»
02.09.2022
Алгоритмы производства фармацевтических препаратов на основе мРНК
01 сентября 2022 г.
Новый алгоритм раскрывает секреты клеточных фабрик
31 августа 2022 г.
Метод микроскопии позволяет выявить скрытые наноструктуры в клетках и тканях
29 августа 2022 г.
Ученые вырастили «синтетический» эмбрион с мозгом и бьющимся сердцем из стволовых клеток
27 августа 2022 г.
Белковые структуры не высечены в камне
26 августа 2022 г.
Прорыв в клеточной инженерии: высокопроизводительный CRISPR без вирусных векторов
25 августа 2022 г.
Ученые разрабатывают новый метод для раскрытия скрытого генома
25 августа 2022 г.
Технология, основанная на CRISPR, направлена на борьбу с глобальными вредителями сельскохозяйственных культур
18 августа 2022 г.
Сложные узоры: построение моста от большого к малому
18 августа 2022 г.
Ученые разработали гель из белков шелка паука для биомедицинских целей
15 августа 2022 г.
Разработка ферментов для решения пластиковой проблемы планеты
13 августа 2022 г.
Химики совершили подвиг «молекулярного редактирования»
09 августа 2022 г.
Редактирование генов с помощью CRISPR/Cas9 может привести к клеточной токсичности и нестабильности генома
09 августа 2022 г.
Инструменты искусственного интеллекта предсказывают регуляторную роль ДНК и трехмерную структуру
09 августа 2022 г.
Без яйцеклетки, спермы или матки: синтетические модели эмбрионов мыши, созданные исключительно из стволовых клеток
06 августа 2022 г.
Манипуляции с хромосомами в живых клетках показывают, что они жидкие
02 августа 2022 г.
Новый метод стимулирования образования биопленки и повышения эффективности биокатализа
01 августа 2022 г.
«Почтовые индексы» сообщают РНК, куда идти
30 июля 2022 г.
Программирование pH
27 июля 2022 г.
Ученые создают живые биосенсоры для отслеживания генетической активности клеток
27 июля 2022 г.
…ещё новости биотехнологииКлеточная стенка растений: сложная и динамичная структура, выявленная ответами генов в условиях стресса
Введение
Клеточная стенка растений представляет собой сложную структуру, выполняющую разнообразные функции на протяжении всего жизненного цикла растения. В дополнение к поддержанию структурной целостности за счет сопротивления внутреннему гидростатическому давлению клеточная стенка обеспечивает гибкость для поддержки клеточного деления, биохимический каркас, обеспечивающий дифференцировку, а также патологический и экологический барьер, защищающий от стресса (Scheller and Ulvskov, 2010; Hamann, 2012; Такер и Колтунов, 2014). Клеточная стенка содержит широкий спектр рецепторов, пор и каналов, которые регулируют движение молекул и ответы на локальные и дальнодействующие элиситоры, включая гормоны, сахара, белки и РНК. В соответствии с ролью во многих процессах, структура клеточной стенки растений невероятно разнообразна не только между видами растений, но и между типами тканей. В общем, два типа стенок, окружающих растительные клетки, часто называют первичной стенкой и вторичной стенкой. Динамическая первичная стенка устанавливается в молодых клетках во время деления и обеспечивает гибкость и базовую структурную поддержку, защищая клетку и опосредуя межклеточные взаимодействия.
В дополнение к поддержанию структурной целостности за счет сопротивления внутреннему гидростатическому давлению клеточная стенка обеспечивает гибкость для поддержки клеточного деления, биохимический каркас, обеспечивающий дифференцировку, а также патологический и экологический барьер, защищающий от стресса (Scheller and Ulvskov, 2010; Hamann, 2012; Такер и Колтунов, 2014). Клеточная стенка содержит широкий спектр рецепторов, пор и каналов, которые регулируют движение молекул и ответы на локальные и дальнодействующие элиситоры, включая гормоны, сахара, белки и РНК. В соответствии с ролью во многих процессах, структура клеточной стенки растений невероятно разнообразна не только между видами растений, но и между типами тканей. В общем, два типа стенок, окружающих растительные клетки, часто называют первичной стенкой и вторичной стенкой. Динамическая первичная стенка устанавливается в молодых клетках во время деления и обеспечивает гибкость и базовую структурную поддержку, защищая клетку и опосредуя межклеточные взаимодействия. Более толстая и прочная вторичная стенка находится между первичной стенкой и плазматической мембраной и откладывается на более поздней стадии, когда клетка перестает расти и делиться. Вторичная стенка рассматривается как важная адаптация, которая позволяет наземным растениям выдерживать и способствовать вертикальному росту.
Более толстая и прочная вторичная стенка находится между первичной стенкой и плазматической мембраной и откладывается на более поздней стадии, когда клетка перестает расти и делиться. Вторичная стенка рассматривается как важная адаптация, которая позволяет наземным растениям выдерживать и способствовать вертикальному росту.
Типичные компоненты клеточной стенки включают целлюлозу, нецеллюлозные и пектиновые полисахариды, белки, фенольные соединения и воду. Основными компонентами (> 90%) являются полисахариды, структура и биосинтез которых были подробно изучены в последнее время (Atmodjo et al., 2013; Pauly et al., 2013; Rennie and Scheller, 2014; Kumar et al. , 2016). Вкратце, целлюлоза представляет собой нерастворимый в воде углевод, присутствующий как в первичных, так и во вторичных клеточных стенках, волокнистая структура которого позволяет поддерживать структурную целостность. Пектины, которые, возможно, являются наиболее сложными и гетерогенными полисахаридами клеточной стенки, существуют преимущественно в первичной клеточной стенке и играют роль в расширении, прочности, пористости, адгезии и межклеточной передаче сигналов.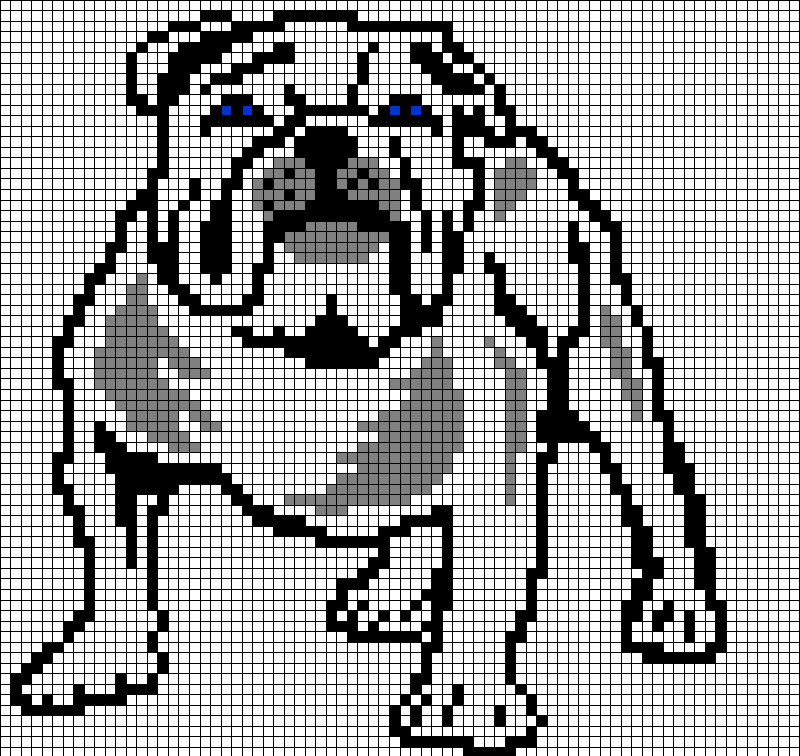 Другие распространенные нецеллюлозные полисахариды включают ксилоглюкан, β-1,3:1,4-глюкан, ксилан, маннан и каллозу, которые выполняют различные роли в механической поддержке, запасании и развитии. В отличие от целлюлозы пектиновые и нецеллюлозные полисахариды можно дополнительно отличить по замещению сахара и боковым цепям, которые присоединяются к полисахаридному остову во время биосинтеза (Шеллер и Ульвсков, 2010). Эти заместители влияют на растворимость, вязкость и взаимодействие с другими полисахаридами и белками в клеточной стенке.
Другие распространенные нецеллюлозные полисахариды включают ксилоглюкан, β-1,3:1,4-глюкан, ксилан, маннан и каллозу, которые выполняют различные роли в механической поддержке, запасании и развитии. В отличие от целлюлозы пектиновые и нецеллюлозные полисахариды можно дополнительно отличить по замещению сахара и боковым цепям, которые присоединяются к полисахаридному остову во время биосинтеза (Шеллер и Ульвсков, 2010). Эти заместители влияют на растворимость, вязкость и взаимодействие с другими полисахаридами и белками в клеточной стенке.
Функция различных компонентов клеточной стенки и то, как они взаимодействуют с экзогенными стимулами, такими как патогены и стресс окружающей среды, вызывает интерес в течение многих лет, особенно в связи с поиском механизмов, с помощью которых можно повысить устойчивость к патогенам, стрессоустойчивость и повысить урожайность сельскохозяйственных культур. быть достигнуто. В недавних обзорах изучалось, как абиотические сигналы изменяют биосинтез целлюлозы (Wang et al. , 2016), как экспансины и пероксидазы влияют на жесткость стенок во время стресса (Tenhaken, 2014) и как модификации нецеллюлозных полисахаридов, таких как ксилоглюкан, сопровождают реакции на стресс (Le Галл и др., 2015). Только недавно в исследованиях начали рассматривать более широкое представление о том, как различные стрессы могут индуцировать сходные изменения в обилии транскриптов (Coolen et al., 2016). Вопрос о том, реагируют ли компоненты клеточной стенки, семейства генов, связанных с клеточной стенкой, или отдельные ортологичные гены клеточной стенки одинаковым образом на различные стрессы у разных видов, подробно не рассматривался, но может дать новые мишени с широкой специфичностью для модификация стрессовых реакций. Некоторые ответы могут быть скрыты в общедоступных наборах данных транскриптомов однодольных и двудольных видов, в которых подробно описаны глобальные транскрипционные реакции на патогены, такие как бактерии, грибы, оомицеты, насекомые и нематоды, а также на абиотические стрессы, такие как засуха, холод и жара.
, 2016), как экспансины и пероксидазы влияют на жесткость стенок во время стресса (Tenhaken, 2014) и как модификации нецеллюлозных полисахаридов, таких как ксилоглюкан, сопровождают реакции на стресс (Le Галл и др., 2015). Только недавно в исследованиях начали рассматривать более широкое представление о том, как различные стрессы могут индуцировать сходные изменения в обилии транскриптов (Coolen et al., 2016). Вопрос о том, реагируют ли компоненты клеточной стенки, семейства генов, связанных с клеточной стенкой, или отдельные ортологичные гены клеточной стенки одинаковым образом на различные стрессы у разных видов, подробно не рассматривался, но может дать новые мишени с широкой специфичностью для модификация стрессовых реакций. Некоторые ответы могут быть скрыты в общедоступных наборах данных транскриптомов однодольных и двудольных видов, в которых подробно описаны глобальные транскрипционные реакции на патогены, такие как бактерии, грибы, оомицеты, насекомые и нематоды, а также на абиотические стрессы, такие как засуха, холод и жара. Эти наборы данных предоставляют ресурсы для идентификации семейств генов, связанных с углеводами, которые кодируют белки с аналогичными функциональными доменами База данных углеводно-активных ферментов (CAZy); (Lombard et al., 2014), а также определить, могут ли определенные семейства, такие как гликозилтрансферазы (GTs), гликозилгидролазы (GHs) и другие ферменты, модифицирующие углеводы, играть ключевую роль в синтезе и модификации клеточной стенки во время стресса. Поэтому после первых двух разделов этого обзора, где мы рассмотрели новые и исторические данные о роли полимеров и генов, связанных с клеточной стенкой, во время биотического и абиотического стресса, мы пересмотрели несколько наборов транскрипционных данных, чтобы обобщить реакцию клеточной стенки и гены и семейства, связанные с углеводами, при стрессе, подчеркивая замечательный уровень сохранения реакций, вызванных различными типами стресса.
Эти наборы данных предоставляют ресурсы для идентификации семейств генов, связанных с углеводами, которые кодируют белки с аналогичными функциональными доменами База данных углеводно-активных ферментов (CAZy); (Lombard et al., 2014), а также определить, могут ли определенные семейства, такие как гликозилтрансферазы (GTs), гликозилгидролазы (GHs) и другие ферменты, модифицирующие углеводы, играть ключевую роль в синтезе и модификации клеточной стенки во время стресса. Поэтому после первых двух разделов этого обзора, где мы рассмотрели новые и исторические данные о роли полимеров и генов, связанных с клеточной стенкой, во время биотического и абиотического стресса, мы пересмотрели несколько наборов транскрипционных данных, чтобы обобщить реакцию клеточной стенки и гены и семейства, связанные с углеводами, при стрессе, подчеркивая замечательный уровень сохранения реакций, вызванных различными типами стресса.
Биотический стресс и клеточная стенка растений
На поле совместной эволюции между растениями и микробами на протяжении миллионов лет растения выработали многоуровневую систему защиты, в которой клеточная стенка служит нескольким целям. Растительная клеточная стенка может служить предварительно сформированным или пассивным структурным барьером, а также индуцированным или активным защитным барьером. Микробы должны обойти клеточную стенку и другие заранее сформированные барьеры, чтобы установить желаемые патогенные отношения с растениями-хозяевами. Это требует соответствующих стратегий распознавания хозяина и разработки подходящих инфекционных структур и/или химического оружия (Zentmyer, 19).61; Турра и др., 2015). Неспособность разработать соответствующие стратегии для разрушения стенки хозяина и других предварительно сформированных структур приводит к тому, что микробы становятся непатогенными и неадаптированными патогенами.
Растительная клеточная стенка может служить предварительно сформированным или пассивным структурным барьером, а также индуцированным или активным защитным барьером. Микробы должны обойти клеточную стенку и другие заранее сформированные барьеры, чтобы установить желаемые патогенные отношения с растениями-хозяевами. Это требует соответствующих стратегий распознавания хозяина и разработки подходящих инфекционных структур и/или химического оружия (Zentmyer, 19).61; Турра и др., 2015). Неспособность разработать соответствующие стратегии для разрушения стенки хозяина и других предварительно сформированных структур приводит к тому, что микробы становятся непатогенными и неадаптированными патогенами.
Растение-хозяин также может использовать клеточную стенку в качестве активного защитного барьера для тех микробов, которые развили механизм преодоления предварительно сформированных барьеров. Во время инфекции элиситоры олигосахаридов высвобождаются из клеточной стенки растения-хозяина (ассоциированные с повреждением молекулярные паттерны, DAMP) или из клеточной стенки патогена (патоген-ассоциированные молекулярные паттерны, PAMP) в результате деградации (Boller and Felix, 2009). ). Растения воспринимают эти элиситоры через иммунные рецепторы плазматической мембраны, которые запускают сигнальные каскады для активации многочисленных защитных реакций, называемых иммунитетом, запускаемым DAMP или PAMP (DTI или PTI; Jones and Dangl, 2006). Одним из распространенных защитных ответов, связанных с DTI или PTI, является укрепление клеточной стенки для создания большей устойчивости к физическому давлению и/или ферментативному гидролизу, генерируемому патогенами (Boller and Felix, 2009; Ringli, 2010; Malinovsky et al., 2014). . В зависимости от типа взаимодействия процесс укрепления клеточной стенки может происходить несколькими различными способами, включая перестройку и сшивание ранее существовавших материалов клеточной стенки, включение легко сшиваемых полимеризованных материалов в существующую клеточную стенку и локальное отложение клеточной стенки. материалов в местах заражения (Moerschbacher and Mendgen, 2012).
). Растения воспринимают эти элиситоры через иммунные рецепторы плазматической мембраны, которые запускают сигнальные каскады для активации многочисленных защитных реакций, называемых иммунитетом, запускаемым DAMP или PAMP (DTI или PTI; Jones and Dangl, 2006). Одним из распространенных защитных ответов, связанных с DTI или PTI, является укрепление клеточной стенки для создания большей устойчивости к физическому давлению и/или ферментативному гидролизу, генерируемому патогенами (Boller and Felix, 2009; Ringli, 2010; Malinovsky et al., 2014). . В зависимости от типа взаимодействия процесс укрепления клеточной стенки может происходить несколькими различными способами, включая перестройку и сшивание ранее существовавших материалов клеточной стенки, включение легко сшиваемых полимеризованных материалов в существующую клеточную стенку и локальное отложение клеточной стенки. материалов в местах заражения (Moerschbacher and Mendgen, 2012).
Состав сосочков и биотрофные патогены
Локальное отложение материалов клеточной стенки, также известное как сосочки, является ранней защитной реакцией, обычно формируемой против инфекции рядом биотрофных, гемибиотрофных и бактериальных патогенов (Bellincampi et al. , 2014). Крошечная структура микрометрового масштаба, образующаяся в месте заражения, часто бывает достаточно большой, чтобы остановить проникновение грибка. У некоторых видов-нехозяев и хозяев резистентность достигается на стадии до инвазии за счет образования сосочков в местах заражения. Однако точная роль сосочков не совсем понятна. Они могут действовать как физический барьер, который эффективно останавливает проникновение патогенов или замедляет процесс проникновения, так что другие защитные механизмы могут быть активированы раньше времени (Stone and Clarke, 19).92; Хюкельховен, 2005). Они также могут функционировать в качестве химического барьера, вмещающего различные виды химического оружия, такие как противомикробные токсины, фитоалексины и дефенсины, которые необходимы для непосредственной атаки на патогены или ингибирования ферментов, разрушающих клеточную стенку, вырабатываемых патогенами (Albersheim et al., 2011). .
, 2014). Крошечная структура микрометрового масштаба, образующаяся в месте заражения, часто бывает достаточно большой, чтобы остановить проникновение грибка. У некоторых видов-нехозяев и хозяев резистентность достигается на стадии до инвазии за счет образования сосочков в местах заражения. Однако точная роль сосочков не совсем понятна. Они могут действовать как физический барьер, который эффективно останавливает проникновение патогенов или замедляет процесс проникновения, так что другие защитные механизмы могут быть активированы раньше времени (Stone and Clarke, 19).92; Хюкельховен, 2005). Они также могут функционировать в качестве химического барьера, вмещающего различные виды химического оружия, такие как противомикробные токсины, фитоалексины и дефенсины, которые необходимы для непосредственной атаки на патогены или ингибирования ферментов, разрушающих клеточную стенку, вырабатываемых патогенами (Albersheim et al., 2011). .
Было высказано предположение, что устойчивость к проникновению, опосредованная сосочками, представляет собой способность генотипа хозяина развивать эффективные сосочки с правильным составом и в нужное время (Aist and Israel, 1977; Иноуэ и др. , 1994). Следовательно, понимание состава сосочков и факторов, влияющих на развитие эффективных сосочков, было в центре внимания многих исследователей. В последние три десятилетия в исследованиях предпринимались попытки идентифицировать компоненты сосочков, образующихся против разных биотрофных патогенов на разных культурах. В то время как некоторые физико-химические изменения, которые происходят во время укрепления клеточных стенок, опосредованного сосочками, в настоящее время хорошо описаны, многие аспекты плохо изучены. Например, было проведено множество исследований, посвященных накоплению и лигнификации папиллярной каллозы из-за наличия флуоресцентных красителей для каллозы и присущей аутофлуоресценции соединений лигнина, в то время как потенциальная роль многих других компонентов клеточной стенки остается неизвестной.
, 1994). Следовательно, понимание состава сосочков и факторов, влияющих на развитие эффективных сосочков, было в центре внимания многих исследователей. В последние три десятилетия в исследованиях предпринимались попытки идентифицировать компоненты сосочков, образующихся против разных биотрофных патогенов на разных культурах. В то время как некоторые физико-химические изменения, которые происходят во время укрепления клеточных стенок, опосредованного сосочками, в настоящее время хорошо описаны, многие аспекты плохо изучены. Например, было проведено множество исследований, посвященных накоплению и лигнификации папиллярной каллозы из-за наличия флуоресцентных красителей для каллозы и присущей аутофлуоресценции соединений лигнина, в то время как потенциальная роль многих других компонентов клеточной стенки остается неизвестной.
Недавняя разработка специфичных к клеточной стенке антител, модулей связывания углеводов и красителей с малыми молекулами дает новую возможность получить информацию о трехмерных изменениях полисахаридов в инфицированных участках клеточной стенки. Чоудхури и др. (2014) использовали эти новые инструменты, чтобы показать, что основные полисахариды, обнаруженные в сосочках ячменя, индуцируются в ответ на грибковый патоген Blumeria graminis f.sp. hordei ( Bgh ) представляют собой каллозу, арабиноксилан и целлюлозу. Эффективные сосочки, успешно предотвращающие попытки проникновения Bgh содержат значительно более высокие концентрации этих полисахаридов по сравнению с неэффективными сосочками. Сосочки состоят из внутреннего ядра, состоящего из каллозы и арабиноксилана, и внешнего слоя, содержащего арабиноксилан и целлюлозу. Сочетание арабиноксилана и целлюлозы с устойчивостью к проникновению открывает новые возможности для улучшения состава сосочков и создания линий с улучшенной устойчивостью к болезням. Предыдущие исследования описывали профили экспрессии генов-кандидатов во время формирования сосочков и обсуждали их вероятные защитные функции (Bhuiyan et al., 2009).). Однако, кроме того, что участие семейства генов glucan synthase-like ( GSL ) в синтезе каллозы сосочков (Jacobs et al.
Чоудхури и др. (2014) использовали эти новые инструменты, чтобы показать, что основные полисахариды, обнаруженные в сосочках ячменя, индуцируются в ответ на грибковый патоген Blumeria graminis f.sp. hordei ( Bgh ) представляют собой каллозу, арабиноксилан и целлюлозу. Эффективные сосочки, успешно предотвращающие попытки проникновения Bgh содержат значительно более высокие концентрации этих полисахаридов по сравнению с неэффективными сосочками. Сосочки состоят из внутреннего ядра, состоящего из каллозы и арабиноксилана, и внешнего слоя, содержащего арабиноксилан и целлюлозу. Сочетание арабиноксилана и целлюлозы с устойчивостью к проникновению открывает новые возможности для улучшения состава сосочков и создания линий с улучшенной устойчивостью к болезням. Предыдущие исследования описывали профили экспрессии генов-кандидатов во время формирования сосочков и обсуждали их вероятные защитные функции (Bhuiyan et al., 2009).). Однако, кроме того, что участие семейства генов glucan synthase-like ( GSL ) в синтезе каллозы сосочков (Jacobs et al. , 2003), гены, участвующие в синтезе остальных полисахаридов сосочков, еще не охарактеризованы. .
, 2003), гены, участвующие в синтезе остальных полисахаридов сосочков, еще не охарактеризованы. .
Некротрофные патогены
Взаимодействие между растительными и некротрофными патогенами происходит на более высоком уровне, чем наблюдаемое с биотрофными патогенами. Хотя целью растительной клетки по-прежнему является предотвращение проникновения патогенов, патогены с некротрофической фазой своего жизненного цикла развили арсенал ферментов, разрушающих клеточную стенку, предназначенных для разрушения клеточной стенки растения, наряду с рядом факторов вирулентности или токсинов. чтобы убить клетки-хозяева и высвободить питательные вещества внутри, а не забрать их тайком (van Kan, 2006). Растения обычно реагируют на некротрофные патогены сильнее, но аналогично биотрофным патогенам, укрепляя клеточную стенку в точке атаки и модифицируя клеточную стенку, чтобы она была более устойчивой к ферментативному перевариванию. Патоген часто использует этот процесс в своих интересах, заставляя растение изменять свою клеточную стенку, чтобы сделать ее более усвояемой (Hok et al. , 2010). Учитывая широко распространенное повреждение, которое может быть вызвано токсинами, можно было бы ожидать большой реакции на ранение из-за потери целостности клеточной стенки (Ferrari et al., 2013). Большинство некротрофных патогенов проникает в ткани растений через устьица и открытые раны, распространяясь между клеточными контактами.
, 2010). Учитывая широко распространенное повреждение, которое может быть вызвано токсинами, можно было бы ожидать большой реакции на ранение из-за потери целостности клеточной стенки (Ferrari et al., 2013). Большинство некротрофных патогенов проникает в ткани растений через устьица и открытые раны, распространяясь между клеточными контактами.
Нафиси и др. (2015) рассмотрели роль клеточной стенки во взаимодействии растение:некротроф, сосредоточив внимание на передаче сигналов фитогормонов ниже по течению. Распознавание сигналов PAMP приводит к активации сигнальных каскадов, которые взаимодействуют с ауксином, цитокинином, брассиностероидами и абсцизовой кислотой, чтобы активировать экспрессию генов, связанных с защитой. Таким образом, восприимчивость клеточной стенки к деградации и последующему производству PAMP важна для устойчивости растения к патогенам. Об этом свидетельствует влияние метилэстерификации пектина на взаимодействие растений и патогенов. Лионетти и др. (2012) рассмотрели роль пектинметилэстераз в ответ на ряд патогенов растений, включая некротрофы, подчеркнув, что деэтерификация пектина влияет на восприимчивость клеточной стенки к ферментам, разрушающим клеточную стенку грибов. Был проведен метаанализ ферментов, модифицирующих пектин, у арабидопсиса, однако семейства гликозилтрансфераз, участвующих в синтезе пектина, не были включены.
Был проведен метаанализ ферментов, модифицирующих пектин, у арабидопсиса, однако семейства гликозилтрансфераз, участвующих в синтезе пектина, не были включены.
Нематоды, паразитирующие на растениях
Ремоделирование клеточной стенки при заражении паразитическими нематодами корней растений, вероятно, является важным компонентом для успешного завершения жизненного цикла нематод (обзор в Bohlmann and Sobczak, 2014). Паразитические нематоды должны проникать, мигрировать и устанавливать питательные структуры (синцитии или гигантские клетки), все из которых требуют определенного взаимодействия со стенками клеток корня. Ранние исследования цистообразующих нематод изучали роль ферментов, разрушающих клеточную стенку, которые секретируются для того, чтобы проникнуть и мигрировать к оптимальному месту питания (рассмотрено в Deubert and Rohde, 19).71), а недавние исследования подтверждают, что смесь ферментов, таких как целлюлазы, 1,3-β-глюканазы и пектинлиазы, обычно связанные с патогенезом растений, сохраняются у различных видов паразитических нематод (Rai et al. , 2015). Совсем недавно акцент сместился на реакцию клеточной стенки растений, поскольку она ремоделируется, чтобы приспособиться к формированию места питания (обзор в Wieczorek, 2015) и различиям, наблюдаемым в восприимчивом и резистентном взаимодействии. Несколько исследований показали специфические изменения полисахаридов стенки, таких как пектин, во время инфекции (Davies and Urwin, 2012) и выдвинули гипотезу о том, что компоненты клеточной стенки, такие как 1,3-β-глюкан или 1,3:1,4-β- глюкан может влиять на поток растворенных веществ между нематодой и хозяином (Hofmann et al., 2010; Aditya et al., 2015).
, 2015). Совсем недавно акцент сместился на реакцию клеточной стенки растений, поскольку она ремоделируется, чтобы приспособиться к формированию места питания (обзор в Wieczorek, 2015) и различиям, наблюдаемым в восприимчивом и резистентном взаимодействии. Несколько исследований показали специфические изменения полисахаридов стенки, таких как пектин, во время инфекции (Davies and Urwin, 2012) и выдвинули гипотезу о том, что компоненты клеточной стенки, такие как 1,3-β-глюкан или 1,3:1,4-β- глюкан может влиять на поток растворенных веществ между нематодой и хозяином (Hofmann et al., 2010; Aditya et al., 2015).
Травоядные насекомые
Реакция растений на нападение травоядных насекомых в значительной степени регулируется реакцией на ранение, вызванной распознаванием DAMP (Boller and Felix, 2009). Механические повреждения, вызванные кормлением насекомыми, могут быть уменьшены за счет утолщения клеточной стенки, однако устойчивость, скорее всего, примет форму химической защиты, такой как фенолы, алкалоиды, терпеноиды или глюкозинолаты (обзор в van Dam, 2009). Прямое воздействие растительных гликозилгидролаз или лектинов на хитин или другие углеводные структуры, присутствующие в пищевых структурах и средней кишке насекомых, играет важную роль в защите травоядных, препятствуя поглощению патогена питательными веществами (rev. Vandenborre et al., 2011).
Прямое воздействие растительных гликозилгидролаз или лектинов на хитин или другие углеводные структуры, присутствующие в пищевых структурах и средней кишке насекомых, играет важную роль в защите травоядных, препятствуя поглощению патогена питательными веществами (rev. Vandenborre et al., 2011).
Последствия модифицированного состава клеточной стенки для патогенеза
Трансгенные и генетические подходы как с усилением, так и с утратой функции использовались для изучения влияния измененного состава клеточной стенки на устойчивость растений к болезням, некоторые из которых обобщены в таблице 1. Эти исследования показывают, что модифицированный состав клеточной стенки действительно может приводить к повышению или снижению фенотипов устойчивости к болезням у растений-хозяев, в зависимости от целевого полисахарида и того, был ли ген, связанный с клеточной стенкой, сверхэкспрессирован или мутирован. Во многих случаях гены-мишени, связанные с клеточной стенкой, идентифицировали с помощью транскриптомных методов после применения специфического биотического стресса (см. ссылки в таблицах 1, 2). Однако также важно отметить, что ряд этих исследований был направлен на улучшение усвояемости кормовых культур, чтобы сделать лигноцеллюлозу менее неподатливой для биопереработки, и есть некоторые опасения, что растения с повышенной усвояемостью из-за измененных свойств клеточных стенок могут быть более восприимчивы. к вредителям и болезням. Данные исследований трансгенных линий с измененными уровнями транскриптов генов-кандидатов, участвующих в путях биосинтеза целлюлозы, нецеллюлозных полисахаридов и лигнина, предполагают, что это может быть не так. Например, снижение биосинтеза целлюлозы с помощью генетики или химических веществ приводит к компенсаторным эффектам целостности клеточной стенки, что приводит к увеличению лигнификации и повышению устойчивости к болезням (Hamann, 2012).
ссылки в таблицах 1, 2). Однако также важно отметить, что ряд этих исследований был направлен на улучшение усвояемости кормовых культур, чтобы сделать лигноцеллюлозу менее неподатливой для биопереработки, и есть некоторые опасения, что растения с повышенной усвояемостью из-за измененных свойств клеточных стенок могут быть более восприимчивы. к вредителям и болезням. Данные исследований трансгенных линий с измененными уровнями транскриптов генов-кандидатов, участвующих в путях биосинтеза целлюлозы, нецеллюлозных полисахаридов и лигнина, предполагают, что это может быть не так. Например, снижение биосинтеза целлюлозы с помощью генетики или химических веществ приводит к компенсаторным эффектам целостности клеточной стенки, что приводит к увеличению лигнификации и повышению устойчивости к болезням (Hamann, 2012).
Таблица 1. Растение: фенотипы устойчивости к биотическому стрессу с измененным составом клеточной стенки .
Таблица 2. Растение:системы стресса, собранные из PLEXdb для метаанализа .
Абиотический стресс
Другим типом внешнего раздражителя, который может повлиять на клеточную стенку растения, является абиотический стресс. Этот тип стресса включает ряд факторов, таких как экстремальные температуры, засуха, наводнения, соленость, атмосферные загрязнители и загрязнители тяжелыми металлами. Часто растение одновременно подвергается нескольким абиотическим стрессам, что может затруднить определение того, какой стресс вызывает наблюдаемую реакцию. Различные изменения в составе клеточных стенок растений при различных абиотических стрессах были изучены и недавно подробно рассмотрены. Ван и др. (2016) обсудили влияние четырех типов абиотического стресса; солевой стресс, доступность воды, световые условия и температура, на одном из аспектов клеточной стенки растений, целлюлозе. Среди подробно обсуждаемых генов есть представители семейства генов CesA , которые, как известно, синтезируют целлюлозу, и другие, которые ранее были идентифицированы как взаимодействующие с CesA s. Ле Галл и др. (2015) дают обзор влияния засухи, жары, холода, солей, тяжелых металлов, света и загрязнителей воздуха на основные компоненты клеточной стенки растений как у однодольных, так и у двудольных растений. Напротив, Tenhaken (2014) сосредоточился на влиянии активных форм кислорода (АФК), которые являются реакцией растений на стресс, на компоненты клеточной стенки растений, такие как XTH и расширения. Члены семейств генов экспансина и XTH часто проявляют дифференциальную экспрессию в условиях абиотического стресса и, следовательно, повышенное присутствие АФК, что приводит к потенциальной паузе в росте. В этом разделе текущего обзора (и в таблице 3) мы приводим краткий обзор исследований, раскрывающих динамику транскрипции генов клеточной стенки во время абиотического стресса, прежде чем сосредоточить внимание на исследованиях однодольных и двудольных растений, которые напрямую связывают влияние ген или семейство генов, связанных с клеточной стенкой, к измененным реакциям на абиотический стресс.
Ле Галл и др. (2015) дают обзор влияния засухи, жары, холода, солей, тяжелых металлов, света и загрязнителей воздуха на основные компоненты клеточной стенки растений как у однодольных, так и у двудольных растений. Напротив, Tenhaken (2014) сосредоточился на влиянии активных форм кислорода (АФК), которые являются реакцией растений на стресс, на компоненты клеточной стенки растений, такие как XTH и расширения. Члены семейств генов экспансина и XTH часто проявляют дифференциальную экспрессию в условиях абиотического стресса и, следовательно, повышенное присутствие АФК, что приводит к потенциальной паузе в росте. В этом разделе текущего обзора (и в таблице 3) мы приводим краткий обзор исследований, раскрывающих динамику транскрипции генов клеточной стенки во время абиотического стресса, прежде чем сосредоточить внимание на исследованиях однодольных и двудольных растений, которые напрямую связывают влияние ген или семейство генов, связанных с клеточной стенкой, к измененным реакциям на абиотический стресс.
Таблица 3. Растение: фенотип реакции на абиотический стресс с измененным составом клеточной стенки .
Глобальное профилирование реакции на абиотический стресс
Транскрипционные изменения, сопровождающие различные абиотические стрессы, обсуждались достаточно подробно (обзор в Santos et al., 2011; Gehan et al., 2015), но удивительно немногие рассматривали эти изменения в контекст специфических генов, связанных с клеточной стенкой. Детальный анализ генетических реакций на засуху отдельных органов колоса ячменя был проведен Abebe et al. (2010). Профили транскрипции ости, семени, цветковой чешуи и цветковой чешуи сравнивали между растениями, пережившими засушливый стресс из-за отсутствия воды в течение 4 дней во время налива зерна, и контрольными растениями. Было обнаружено, что для всех тканей, кроме семян, множественные гены, связанные с клеточной стенкой, по-разному регулируются между контрольными растениями и растениями, подвергшимися засухе. Гены, кодирующие представителей целлюлозсинтазы (GT2, CesA), UDP-ксилозилтрансферазы, семейства гликозилгидролаз 1 (Gh2), эндо-бета-1,4-глюканазы (GH9) и семейства ксилоглюкан-эндотрансгликозилазы (Gh26, XTH/XET) были среди генов, связанных с клеточной стенкой, которые, как наблюдалось, подавлялись в условиях засухи. Дополнительный XET активировался в условиях засухи, а также предполагаемый ингибитор ксиланазы, эндо-1,3-бета-глюкозидаза и бета-D-глюканэкзогидролаза. Подобные исследования у арабидопсиса выявили более 500 генов, которые реагируют на засуху, холод и стресс от засоления (Seki et al., 2002), включая несколько членов семейств экстенсинов, пектинэстераз и XTH/XET, которые были подавлены. Ван и др. (2013) показали, что только в случае засоления более 140 генов, связанных с клеточной стенкой, реагируют на солевой стресс, а иногда и по-разному между экотипами Arabidopsis. Как ранее было установлено другими авторами в условиях засухи (Wu and Cosgrove, 2000; Moore et al., 2008), в зависимости от того, какая ткань наблюдается, клеточная стенка растения либо ослабевает, либо сжимается для поддержания роста.
Гены, кодирующие представителей целлюлозсинтазы (GT2, CesA), UDP-ксилозилтрансферазы, семейства гликозилгидролаз 1 (Gh2), эндо-бета-1,4-глюканазы (GH9) и семейства ксилоглюкан-эндотрансгликозилазы (Gh26, XTH/XET) были среди генов, связанных с клеточной стенкой, которые, как наблюдалось, подавлялись в условиях засухи. Дополнительный XET активировался в условиях засухи, а также предполагаемый ингибитор ксиланазы, эндо-1,3-бета-глюкозидаза и бета-D-глюканэкзогидролаза. Подобные исследования у арабидопсиса выявили более 500 генов, которые реагируют на засуху, холод и стресс от засоления (Seki et al., 2002), включая несколько членов семейств экстенсинов, пектинэстераз и XTH/XET, которые были подавлены. Ван и др. (2013) показали, что только в случае засоления более 140 генов, связанных с клеточной стенкой, реагируют на солевой стресс, а иногда и по-разному между экотипами Arabidopsis. Как ранее было установлено другими авторами в условиях засухи (Wu and Cosgrove, 2000; Moore et al., 2008), в зависимости от того, какая ткань наблюдается, клеточная стенка растения либо ослабевает, либо сжимается для поддержания роста. Это иллюстрирует сложность реакции клеточной стенки на абиотические стрессы.
Это иллюстрирует сложность реакции клеточной стенки на абиотические стрессы.
Растения могут испытывать абиотический стресс разной степени тяжести, поэтому многие исследования включают несколько уровней лечения стресса, чтобы понять, как это влияет на реакцию. Харб и др. (2010) оценили влияние прогрессирующей засухи и умеренной засухи на рост растений с помощью ряда биохимических и физиологических анализов, а изменения в экспрессии генов отслеживали с помощью эксперимента с микроматрицами. В условиях умеренной засухи рост растений значительно снижался как в плане накопления биомассы, так и в плане расширения листьев, а также устьичной проводимости. В этих условиях несколько генов, кодирующих экспансины клеточной стенки, активировались; однако в условиях прогрессирующей засухи экспансины клеточной стенки подавлялись. Экспансины представляют собой белки, которые, как было показано ранее, ослабляют и модифицируют клеточную стенку растений во время роста и адаптации к стрессу путем модификации целлюлозных и нецеллюлозных компонентов клеточной стенки (Cosgrove, 2005). Применяя аналогичный подход, Mangelsen et al. (2011) подвергали молодые зерновки ячменя тепловому стрессу в течение 0,5, 3 и 6 часов и использовали микрочипы для идентификации дифференциально экспрессируемых генов. Гены с пониженной регуляцией, связанные с клеточной стенкой, были статистически чрезмерно представлены, особенно после 3 и 6 часов воздействия теплового стресса, которые авторы описали как фазы первичной тепловой реакции и адаптации к тепловому стрессу соответственно. Этот набор включал гены, функционально аннотированные как пектатлиазы, полигалактуроназы и пектинэстеразы.
Применяя аналогичный подход, Mangelsen et al. (2011) подвергали молодые зерновки ячменя тепловому стрессу в течение 0,5, 3 и 6 часов и использовали микрочипы для идентификации дифференциально экспрессируемых генов. Гены с пониженной регуляцией, связанные с клеточной стенкой, были статистически чрезмерно представлены, особенно после 3 и 6 часов воздействия теплового стресса, которые авторы описали как фазы первичной тепловой реакции и адаптации к тепловому стрессу соответственно. Этот набор включал гены, функционально аннотированные как пектатлиазы, полигалактуроназы и пектинэстеразы.
В других подходах сравнивали транскриптомные данные чувствительных к стрессу и толерантных сортов. Кэл и др. (2013) сравнили данные транскриптома для зоны удлинения листа (LEZ) двух сортов риса, Moroberekan, который является засухоустойчивым, и IR64, который чувствителен к засухе, в условиях водного дефицита. Эта ткань была выбрана потому, что изменения в экспансии в LEZ часто являются одним из более ранних ответов на дефицит воды (Cutler et al. , 1980). Эти наборы транскриптомных данных идентифицировали набор генов, которые показали более чем двукратное изменение экспрессии в обоих сортах, включая 27 генов, связанных с клеточной стенкой, большинство из которых были подавлены в засухоустойчивом сорте Мороберекан. Список подавленных генов включает гены, участвующие в производстве вторичной клеточной стенки, включая циннамоил-КоА-редуктазу, ферулат-5-гидроксилазу, лакказу и апопластные пероксидазы класса III. Гены, кодирующие арабиногалактановые белки и экстенсины, участвующие в передаче сигналов и структуре клеточной стенки, XTH/XET и GT, включая два генов CesA по-разному экспрессировались между двумя сортами. Два гена, экспрессия которых была обнаружена в Мороберекане, являются членами семейства гликозилгидролаз Gh38, кодирующего полигалактуроназы. В аналогичном исследовании Zheng et al. (2010) сравнили данные по полногеномной экспрессии генов для линий кукурузы Han21 и Ye478, устойчивых к засухе и неустойчивых к засухе, соответственно, в условиях засушливого стресса.
, 1980). Эти наборы транскриптомных данных идентифицировали набор генов, которые показали более чем двукратное изменение экспрессии в обоих сортах, включая 27 генов, связанных с клеточной стенкой, большинство из которых были подавлены в засухоустойчивом сорте Мороберекан. Список подавленных генов включает гены, участвующие в производстве вторичной клеточной стенки, включая циннамоил-КоА-редуктазу, ферулат-5-гидроксилазу, лакказу и апопластные пероксидазы класса III. Гены, кодирующие арабиногалактановые белки и экстенсины, участвующие в передаче сигналов и структуре клеточной стенки, XTH/XET и GT, включая два генов CesA по-разному экспрессировались между двумя сортами. Два гена, экспрессия которых была обнаружена в Мороберекане, являются членами семейства гликозилгидролаз Gh38, кодирующего полигалактуроназы. В аналогичном исследовании Zheng et al. (2010) сравнили данные по полногеномной экспрессии генов для линий кукурузы Han21 и Ye478, устойчивых к засухе и неустойчивых к засухе, соответственно, в условиях засушливого стресса. В общей сложности 15 наборов зондов, которые кодируют предполагаемые гены, связанные с клеточной стенкой, по-разному экспрессировались между двумя линиями. К ним относятся зонды, аннотированные как субъединицы целлюлозосинтазы, предшественники эндо-1,3-β-глюкозидазы и предшественники белка COBRA-подобного 3.
В общей сложности 15 наборов зондов, которые кодируют предполагаемые гены, связанные с клеточной стенкой, по-разному экспрессировались между двумя линиями. К ним относятся зонды, аннотированные как субъединицы целлюлозосинтазы, предшественники эндо-1,3-β-глюкозидазы и предшественники белка COBRA-подобного 3.
Как упоминалось ранее в этом обзоре, растения часто одновременно подвергаются множественным стрессам, в том числе абиотическим и биотическим, и их эффекты не обязательно просто аддитивны (Puranik et al., 2012; Coolen et al., 2016). Поэтому важно, несмотря на очевидную сложность подобных экспериментов, изучить влияние на растения многократных одновременных стрессов. Было обнаружено, что AtRALFL8 активируется в корнях 10-дневных растений, когда применялись двойные стрессы заражения нематодами и водного дефицита (Atkinson et al., 2013). Последующий анализ микрочипов показал, что в этих условиях AtRALFL8 в высокой степени ко-экспрессируется с пектиназами, известными своей способностью способствовать ремоделированию клеточной стенки, ранее было показано, что они играют роль в нескольких реакциях на стресс, включая заражение нематодами (Pelloux et al. , 2007; An et al., 2008). Кулен и др. (2016) подвергали растения арабидопсиса одиночному и двойному сочетанию стресса от засухи, грибковой инфекции Botrytis cinerea и травоядной инфекции Pieris rapae . В общей сложности 41 ген, связанный с клеточной стенкой, включая PECTIN METHYLESTERASE 3 ( PME3 ), EXPANSIN A6 ( EXPA6 ), XTh20 , and XTh42 responded to at least one stress, while 12 genes including CELLULOSE SYNTHASE LIKE G2 ( CSLG2 ), АРАБИНОГАЛАКТАН БЕЛОК 2 ( AGP2 ), БЕТА ГЛЮКОЗИДАЗА 46 ( BGLU46 ), 1,3-β-глюканаза и EXPANSIN A8 ( ) реагировали на все одинаково. три. Это указывает на то, что общие транскрипционные ответы и, возможно, последующие эффекты на состав клеточной стенки используются в ответ на разные стрессы.
, 2007; An et al., 2008). Кулен и др. (2016) подвергали растения арабидопсиса одиночному и двойному сочетанию стресса от засухи, грибковой инфекции Botrytis cinerea и травоядной инфекции Pieris rapae . В общей сложности 41 ген, связанный с клеточной стенкой, включая PECTIN METHYLESTERASE 3 ( PME3 ), EXPANSIN A6 ( EXPA6 ), XTh20 , and XTh42 responded to at least one stress, while 12 genes including CELLULOSE SYNTHASE LIKE G2 ( CSLG2 ), АРАБИНОГАЛАКТАН БЕЛОК 2 ( AGP2 ), БЕТА ГЛЮКОЗИДАЗА 46 ( BGLU46 ), 1,3-β-глюканаза и EXPANSIN A8 ( ) реагировали на все одинаково. три. Это указывает на то, что общие транскрипционные ответы и, возможно, последующие эффекты на состав клеточной стенки используются в ответ на разные стрессы.
Генетические и трансгенные данные, подтверждающие роль генов клеточной стенки в реакции на абиотический стресс абиотический стресс.
 У арабидопсиса AtCesA8 / IRX1 способствует синтезу вторичной клеточной стенки и влияет на устойчивость растений к засухе и осмотическому стрессу (Chen et al., 2005). Мутантные аллели AtCesA8, увядание листьев 2-1 ( lew2-1 ) и lew2-2 продемонстрировали более высокую устойчивость к осмотическим стрессам, вызванным воздействием NaCl и маннита, и стрессу от засухи по сравнению с растениями дикого типа. Другие компоненты клеточной стенки растений влияют на устойчивость растений к холоду или морозу. Например, Такета и др. (2012) провели скрининг мутантов ячменя, индуцированных азидом натрия, которые были чувствительны к охлаждению. Было обнаружено, что из 11 идентифицированных линий в 2 отсутствует (1,3:1,4)-β-глюкан, и они содержат мутации в HvCslF6 , ген, связанный с клеточной стенкой, который ранее был вовлечен в синтез (1,3:1,4)-β-глюкана (Burton et al., 2006). Хотя эти линии не содержали мутаций, которые продуцировали бы преждевременный стоп-кодон, одна мутация располагалась в непосредственной близости от консервативного каталитического мотива HvCslF6 гликозилтрансферазы (GT2).
У арабидопсиса AtCesA8 / IRX1 способствует синтезу вторичной клеточной стенки и влияет на устойчивость растений к засухе и осмотическому стрессу (Chen et al., 2005). Мутантные аллели AtCesA8, увядание листьев 2-1 ( lew2-1 ) и lew2-2 продемонстрировали более высокую устойчивость к осмотическим стрессам, вызванным воздействием NaCl и маннита, и стрессу от засухи по сравнению с растениями дикого типа. Другие компоненты клеточной стенки растений влияют на устойчивость растений к холоду или морозу. Например, Такета и др. (2012) провели скрининг мутантов ячменя, индуцированных азидом натрия, которые были чувствительны к охлаждению. Было обнаружено, что из 11 идентифицированных линий в 2 отсутствует (1,3:1,4)-β-глюкан, и они содержат мутации в HvCslF6 , ген, связанный с клеточной стенкой, который ранее был вовлечен в синтез (1,3:1,4)-β-глюкана (Burton et al., 2006). Хотя эти линии не содержали мутаций, которые продуцировали бы преждевременный стоп-кодон, одна мутация располагалась в непосредственной близости от консервативного каталитического мотива HvCslF6 гликозилтрансферазы (GT2). Такета и др. (2012) предположили, что повышенная чувствительность вегетативных тканей к охлаждению у линий, содержащих мутации в HvCslF6 может быть связано с более тонкими клеточными стенками, поскольку (1,3:1,4)-β-глюкан обычно является основным компонентом этой структуры у трав. Эта гипотеза была подкреплена работой Cu et al. (2016), которые наблюдали более тонкие клеточные стенки в линиях нокдауна CslF6 , полученных с помощью РНКи, по сравнению с диким типом, используя как метод окрашивания Calcofluor, так и иммуноцитологическое окрашивание с помощью BG1 (1,3:1,4)-β-глюкан-специфического антитела. . Интересно, что содержание (1,3:1,4)-β-глюкана в зерне злаков, по-видимому, особенно чувствительно к условиям окружающей среды, хотя неясно, зависит ли это изменение только от модифицированного HvCslF6 функция. Суонстон и др. (1997) и Wallwork et al. (1998) выявили значительные различия в содержании (1,3:1,4)-β-глюкана в зерне ячменя в зависимости от участка поля или температуры во время созревания зерна.
Такета и др. (2012) предположили, что повышенная чувствительность вегетативных тканей к охлаждению у линий, содержащих мутации в HvCslF6 может быть связано с более тонкими клеточными стенками, поскольку (1,3:1,4)-β-глюкан обычно является основным компонентом этой структуры у трав. Эта гипотеза была подкреплена работой Cu et al. (2016), которые наблюдали более тонкие клеточные стенки в линиях нокдауна CslF6 , полученных с помощью РНКи, по сравнению с диким типом, используя как метод окрашивания Calcofluor, так и иммуноцитологическое окрашивание с помощью BG1 (1,3:1,4)-β-глюкан-специфического антитела. . Интересно, что содержание (1,3:1,4)-β-глюкана в зерне злаков, по-видимому, особенно чувствительно к условиям окружающей среды, хотя неясно, зависит ли это изменение только от модифицированного HvCslF6 функция. Суонстон и др. (1997) и Wallwork et al. (1998) выявили значительные различия в содержании (1,3:1,4)-β-глюкана в зерне ячменя в зависимости от участка поля или температуры во время созревания зерна. Аналогичным образом у нескольких сортов пшеницы, выращенных в различных условиях жары и засухи, наблюдалось снижение содержания (1,3:1,4)-β-глюкана в зерне у линий, выращенных в стрессовых условиях (Rakszegi et al., 2014). И наоборот, в тех же условиях сообщалось об увеличении содержания арабиноксилана в зерне, что, возможно, способствовало снижению содержания (1,3:1,4)-β-глюкана. Генетическая основа этих переменных реакций на абиотический стресс еще не раскрыта.
Аналогичным образом у нескольких сортов пшеницы, выращенных в различных условиях жары и засухи, наблюдалось снижение содержания (1,3:1,4)-β-глюкана в зерне у линий, выращенных в стрессовых условиях (Rakszegi et al., 2014). И наоборот, в тех же условиях сообщалось об увеличении содержания арабиноксилана в зерне, что, возможно, способствовало снижению содержания (1,3:1,4)-β-глюкана. Генетическая основа этих переменных реакций на абиотический стресс еще не раскрыта. Трансгенные растения также использовались для проверки роли генов-кандидатов, потенциально участвующих в продукции/модификации клеточной стенки и абиотическом стрессе. Обычно считается, что ферменты XET/XTH играют роль в ослаблении клеточной стенки и, следовательно, в расширении клеток (Rose et al., 2002). Трансгенные линии Arabidopsis, экспрессирующие XTH из Capsicum annuum , демонстрируют аномальные фенотипы листьев, включая неправильный рисунок клеток на поперечных срезах и скрученные листья (Cho et al. , 2006). Кроме того, трансгенные линии арабидопсиса и томатов (Choi et al., 2011), экспрессирующие Capsicum XTH продемонстрировал повышенную солеустойчивость и более длинные корни, чем контрольные растения без трансгена, что указывает на роль гибкости стенок в смягчении реакции на стресс. Было обнаружено, что в тканях корней кукурузы множественные гены, связанные с клеточной стенкой, по-разному экспрессируются при солевом стрессе (Li et al., 2014), включая ZmXET1. ZmXET1 , как полагают, участвует в удлинении клеточной стенки, поскольку он способен гидролизовать и повторно соединять молекулы ксилоглюкана (Fry et al., 1992). Другие гены, идентифицированные Li et al. (2014), поскольку экспансины 9 активировались, когда растения подвергались воздействию повышенной засоленности, и, следовательно, возможно, участвовали в опосредовании устойчивости к токсичности, связанной с засолением.1100 ZmEXPA1, ZmEXPA3, ZmEXPA5, ZmEXPB1, ZmEXPB2 . Экспрессия этих генов, связанных с клеточной стенкой, может находиться под эпигенетическим контролем, поскольку повышенная экспрессия генов ZmHATB и ZmGCN5 гистоновых ацетилтрансфераз увеличивалась после солевого стресса и сопровождалась повышенным ацетилированием гистонов h4K9 и h5K5.
, 2006). Кроме того, трансгенные линии арабидопсиса и томатов (Choi et al., 2011), экспрессирующие Capsicum XTH продемонстрировал повышенную солеустойчивость и более длинные корни, чем контрольные растения без трансгена, что указывает на роль гибкости стенок в смягчении реакции на стресс. Было обнаружено, что в тканях корней кукурузы множественные гены, связанные с клеточной стенкой, по-разному экспрессируются при солевом стрессе (Li et al., 2014), включая ZmXET1. ZmXET1 , как полагают, участвует в удлинении клеточной стенки, поскольку он способен гидролизовать и повторно соединять молекулы ксилоглюкана (Fry et al., 1992). Другие гены, идентифицированные Li et al. (2014), поскольку экспансины 9 активировались, когда растения подвергались воздействию повышенной засоленности, и, следовательно, возможно, участвовали в опосредовании устойчивости к токсичности, связанной с засолением.1100 ZmEXPA1, ZmEXPA3, ZmEXPA5, ZmEXPB1, ZmEXPB2 . Экспрессия этих генов, связанных с клеточной стенкой, может находиться под эпигенетическим контролем, поскольку повышенная экспрессия генов ZmHATB и ZmGCN5 гистоновых ацетилтрансфераз увеличивалась после солевого стресса и сопровождалась повышенным ацетилированием гистонов h4K9 и h5K5. В отдельном исследовании Liu et al. (2014) было обнаружено, что сверхэкспрессия OsBURP16 увеличивает количество полигалактуроназы (PG), фермента, гидролизующего пектин, и изменяет состав клеточной стенки растений. Следовательно, растения риса, сверхэкспрессирующие OsBURP16 продемонстрировали меньшую устойчивость к засухе (количественно определяемую как выживаемость после лишения 2-недельных растений воды), при этом растения дикого типа показали выживаемость 42% по сравнению с <10% для линий сверхэкспрессии OsBURP16 . Измерение уровней H 2 O 2 , индикатора стресса, показало, что линии OsBURP16 со сверхэкспрессией также были более восприимчивы к солевому стрессу, чем растения дикого типа.
В отдельном исследовании Liu et al. (2014) было обнаружено, что сверхэкспрессия OsBURP16 увеличивает количество полигалактуроназы (PG), фермента, гидролизующего пектин, и изменяет состав клеточной стенки растений. Следовательно, растения риса, сверхэкспрессирующие OsBURP16 продемонстрировали меньшую устойчивость к засухе (количественно определяемую как выживаемость после лишения 2-недельных растений воды), при этом растения дикого типа показали выживаемость 42% по сравнению с <10% для линий сверхэкспрессии OsBURP16 . Измерение уровней H 2 O 2 , индикатора стресса, показало, что линии OsBURP16 со сверхэкспрессией также были более восприимчивы к солевому стрессу, чем растения дикого типа.
Выявление сети реагирования на стресс клеточной стенки
По мере того, как мы узнаем больше о сетях генов, регулирующих синтез и гидролиз клеточной стенки растений, возможно, что путем ассоциации будет выявлено больше генов, участвующих в реакции на стресс. Недавнее подробное исследование, в котором использовались методы in vitro и in vivo для всесторонней характеристики сети генов, регулирующих синтез вторичной клеточной стенки у арабидопсиса, также показало, как на одну часть этой сети влиял абиотический стресс (Taylor-Teeples et al. др., 2015). Авторы описали регуляторную сеть ксилемы и то, как изменения солености и железа могут вызывать возмущения, которые, в свою очередь, вызывают фенотипические изменения во вторичной клеточной стенке.
Недавнее подробное исследование, в котором использовались методы in vitro и in vivo для всесторонней характеристики сети генов, регулирующих синтез вторичной клеточной стенки у арабидопсиса, также показало, как на одну часть этой сети влиял абиотический стресс (Taylor-Teeples et al. др., 2015). Авторы описали регуляторную сеть ксилемы и то, как изменения солености и железа могут вызывать возмущения, которые, в свою очередь, вызывают фенотипические изменения во вторичной клеточной стенке.
Имеется большой объем данных предыдущих исследований абиотического стресса, которые подробно описывают глобальную реакцию транскрипции генов, а в некоторых случаях – оценку изменений фенотипов клеточных стенок в качестве реакции. Аналогичные наборы данных доступны для биотических стрессов, применяемых к различным видам и тканям. Из генетических исследований, рассмотренных выше, видно, что в реакцию на разные стрессы часто вовлечены сходные члены семейств генов (например, семейства XET/XTH, экспансинов и пектиновых модификаторов). Однако присущая стенке сложность и большое количество генов, участвующих в ее синтезе и модификации, означает, что многие детали в отношении генетической и биохимической основы ответа клеточной стенки на стресс остаются неясными. Несмотря на очевидные трудности, связанные со сравнением экспериментов между разными видами, стрессами и тканями, в заключительном разделе этого обзора мы пересмотрели общедоступные наборы данных транскриптома, чтобы подчеркнуть широкое сходство между различными типами стресса и подумать, можно ли уделить больше внимания предполагаемой клеточной стенке. родственные гены, которые ранее не учитывались.
Однако присущая стенке сложность и большое количество генов, участвующих в ее синтезе и модификации, означает, что многие детали в отношении генетической и биохимической основы ответа клеточной стенки на стресс остаются неясными. Несмотря на очевидные трудности, связанные со сравнением экспериментов между разными видами, стрессами и тканями, в заключительном разделе этого обзора мы пересмотрели общедоступные наборы данных транскриптома, чтобы подчеркнуть широкое сходство между различными типами стресса и подумать, можно ли уделить больше внимания предполагаемой клеточной стенке. родственные гены, которые ранее не учитывались.
Общедоступные наборы данных освещают сложные реакции на уровне транскриптома на абиотический и биотический стресс
В предыдущих разделах этого обзора обобщены исследования, проведенные по различным аспектам укрепления и модификации клеточной стенки во время патогенной инфекции и абиотического стресса. Укрепление клеточной стенки в виде сосочков — относительно распространенный механизм, определяющий исход инфекции. Однако, учитывая разнообразие биотических стрессов, которым может подвергаться растение, любые общие черты в формировании сосочков, вероятно, будут сопровождаться рядом различных защитных реакций, связанных с клеточной стенкой. То же самое можно ожидать и в отношении различных абиотических стрессов, таких как экстремальные температуры, соленость и наводнения. Что касается перекрытия биотических и абиотических стрессов, недавнее исследование арабидопсиса показало, что ~ 25% транскриптов, связанных с клеточной стенкой, которые реагировали на грибковую инфекцию, растительноядность или засуху, давали сходную реакцию при каждом воздействии (Coolen et al. , 2016). Хотя в настоящее время невозможно провести подробный обзор всех изменений клеточных стенок, вызванных реакцией на ряд различных биотических и абиотических стрессов, можно провести метаанализ с использованием общедоступных данных об экспрессии транскриптов растений-патогенов и взаимодействия растений и стресса, чтобы выявить перекрытия в реакциях механизмов клеточной стенки.
Данные об экспрессии генов доступны в Базе данных экспрессии растений (PLEXdb; Dash et al., 2012), включая множество наборов данных микрочипов арабидопсиса и ячменя, в которых подробно описаны изменения в количестве транскриптов после воздействия различных абиотических или биотических стрессов (таблица 2). Дополнительным источником значительного использования является база данных Carbohydrate-Active EnZYmes (CAZy) (Lombard et al., 2014), в которой описаны семейства структурно родственных ферментов, которые гидролизуют, модифицируют или создают гликозидные связи. Используя эту информацию, предполагаемый 9Было отобрано 1100 генов CAZy Arabidopsis , присутствующих в массиве генома Affymetrix 22K ATh2. Домены семейства белков (Pfam), связанные с аннотациями базы данных CAZy, использовались для идентификации генов, связанных с углеводами ячменя, присутствующих на 22K Barley1 GeneChip. Нормализованные уровни транскриптов для каждого гена, связанного с углеводами, из Arabidopsis и ячменя сравнивали после каждого стресса (относительно необработанных контролей) в каждом эксперименте и представляли как логарифмическую (2)-кратную индукцию. Как и следовало ожидать из предыдущих разделов этого обзора, многие гены клеточной стенки продемонстрировали выраженную реакцию на различные стрессы.
Чтобы проверить, могут ли эти ответы быть более консервативными на уровне семейства генов CAZy, была рассчитана и проанализирована средняя кратность индукции, наблюдаемая для всех членов семейства, с использованием средства просмотра мультиэкспериментов TIGR (МэВ). Иерархическая кластеризация использовалась для упорядочения семейств генов в соответствии со сходством паттерна экспрессии генов (рис. 1А, В) (Eisen et al., 1998). На рисунках 1A, B ясно показано, что большинство семейств генов CAZy активируются в ответ на абиотический или биотический стресс у 9 животных.1100 арабидопсис и ячмень. Хотя не все семейства CAZy содержат членов, которые действуют на один и тот же субстрат, и вероятность того, что все специализированные члены семейства реагируют одинаковым образом, невелика, этот подход был нацелен на предоставление простых средств идентификации ключевых видов активности, связанных с углеводами, которые являются общими для разных групп. разные стрессы. Сходное поведение хорошо охарактеризованных и плохо охарактеризованных семейств CAZy может дать полезную информацию о новых связанных со стрессом изменениях клеточной стенки и связанных с углеводами. Выявить тенденции, сохраняющиеся в ответ на напряжения между Arabidopsis и ячменя кратность индукции для каждого семейства генов была усреднена для всех абиотических и всех биотических стрессов и представлена на рисунке 1C. Сравнительные реакции этих семейств генов на абиотический и биотический стресс показаны на рисунке 2 у обоих видов.
Рисунок 1. Анализ транскриптов, связанных с клеточной стенкой, после абиотических и биотических стрессов у арабидопсиса (A) и ячменя (B) . Обилие транскриптов определяли путем метаанализа наборов данных микрочипов, собранных из базы данных экспрессии растений (PLEXdb; Dash et al., 2012), с использованием экспериментов, перечисленных в таблице 2. Значения показывают среднее логарифмическое (2)-кратное увеличение индукции для представителей каждого Семейство генов CAZy присутствует на массиве генома ATh2 Arabidopsis Affymetrix 22K ATh2 и на генетическом чипе 22K Barley1. Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. Тенденции, законсервированные в ответ на стрессы между арабидопсисом и ячменем, наблюдаются в (C) , который показывает среднюю кратность индукции для каждого семейства генов для всех абиотических и всех биотических стрессов у арабидопсиса и ячменя. Звездочками отмечены семейства генов, экспрессия которых усиливается как абиотическими, так и биотическими стрессами у арабидопсиса и ячменя.
Рисунок 2. Графическое представление средней логарифмической (2)-кратной индукции для каждого семейства генов (представлено на рисунке 1C), которая показывает средний абиотический (ось x ) и средний биотический ( y 9ось 1101) реакция на стресс у Arabidopsis (A) и ячменя (B) . Семейства CAZy, которые активируются в ответ на абиотические, но не биотические стрессы, окрашены в красный цвет, семейства CAZy, которые активируются в ответ на биотические стрессы, но не в ответ на абиотические стрессы, окрашены в желтый цвет, а семейства CAZy, которые активируются в ответ как на абиотические, так и на абиотические стрессы, биотические стрессы окрашены в оранжевый цвет.
Были рассмотрены два различных метода кластеризации. Внутри каждого вида семейства генов могут быть сгруппированы на основе корреляции их профилей транскриптов в каждом эксперименте. Во-вторых, наборы экспериментальных данных также могут быть сгруппированы на основе корреляции профилей транскриптов семейства генов. Интересно отметить, что абиотические стрессы обычно образуют кластер вместе, как и биотические стрессы, даже несмотря на то, что в каждом кластере существует огромная разница в типе стресса (рис. 1А, В). Догма о том, что стандартная защитная реакция клеточной стенки в первую очередь обусловлена каллозой и похожих на глюкансинтазу генов семейства GT48 CAZy трудно поддерживать, учитывая большое количество семейств генов, которые, по-видимому, активируются при большинстве стрессов. Несмотря на то, что отдельные гены, индуцируемые в каждом эксперименте, различаются, кластеризация семейств CAZy в экспериментах предполагает, что существует сходная защитная реакция независимо от конкретного типа стресса. Мы можем видеть примеры биотрофных грибов, вызывающих аналогичные реакции на некротрофные грибы, стресс, вызванный засухой, вызывающий аналогичные реакции на холодовой стресс, и даже примеры в разных тканях с нематодами в корнях по сравнению с листьями, зараженными белокрылкой.
Основанная на знаниях идентификация родственных углеводам семейств, которые реагируют на биотический и абиотический стресс
Кластеризация реакции семейств CAZy позволяет выявить активность, которая, по-видимому, обычно активируется в большинстве экспериментов, и, следовательно, клеточная стенка или углеводные компоненты могут быть изменены аналогично во время взаимодействия. На фигуре 1C показана средняя кратность индукции для каждого семейства генов CAZy при всех абиотических стрессах арабидопсиса, биотических стрессах арабидопсиса, абиотических стрессах ячменя и биотических стрессах ячменя. Существуют примеры семейств CAZy, которые, по-видимому, активируются (в среднем) только у Arabidopsis, включая арабиногалактановые белки (AGP), экспансины, фасциклин-подобные арабиногалактановые белки (FLA), пектинлиазу (PL1), пектин-ацетилэстеразу (CE13). , гликозилгидролазы (GH9, GH85) и ряд гликозилтрансфераз (GT4, GT20, GT21, GT47 и GT64). Было обнаружено, что число семейств CAZy, в среднем выше в ячмене, включает ксилан-ацетилэстеразу (CE6) и гликозилтрансферазу (GT14). Учитывая различия между клеточными стенками ячменя и арабидопсиса: стенки ячменя содержат больше арабиноксилана и (1,3:1,4)-β-глюкана, а стенки арабидопсиса содержат больше пектина и ксилоглюкана (Burton et al., 2010), неожиданным было увидеть большую представленность ферментов, модифицирующих пектин, в наборе данных Arabidopsis.
Двенадцать семейств генов CAZy активируются (в среднем) при абиотических и биотических стрессах у арабидопсиса и ячменя. К ним относятся ферменты, расщепляющие и модифицирующие полисахариды, такие как пектинметилэстераза (CE8), углеводная эстераза (CE10) и гликозилгидролазы (Gh2, Gh27, Gh28 и Gh29), которые нацелены на ряд полисахаридов и олигосахаридов, содержащих 1,3-β-глюкан. и хитин. Некоторые из этих семейств CAZy уже вовлечены в реакцию на стресс как белки, связанные с патогенезом (PR). Гены Gh27 были классифицированы как белки PR-2, разлагающие 1,3-β-глюкан (Leubner-Metzger and Meins, 19).99), в то время как Gh28 и Gh29 представляют пять из 17 семейств растительных PR белков (Minic, 2008). Члены семейства Gh2 вовлечены в активацию защитных соединений посредством удаления β-глюкозида (Poulton, 1990; Duroux et al., 1998). Метилэстеразы пектина модифицируют статус этерификации пектина в стенке, влияя на восприимчивость барьера клеточной стенки к грибковым и бактериальным CWDE (Collmer and Keen, 1986). Деэстерификация пектина также влияет на пористость плазмодесм, что может изменить распространение сигнальных молекул во время защитной реакции (Chen et al., 2000).
Роль семейств CAZy гликозилтрансфераз во время защитной реакции менее изучена, чем роль гидролитических ферментов. Примечательно, что существует пять семейств CAZy GT, которые в среднем активируются при абиотических и биотических стрессах у Arabidopsis и ячменя, включая GT1, GT8, GT61, GT75 и GT92. Семейство GT1 включает большое количество генов с широким спектром предполагаемых функций, включая активность UDP-глюкуронозилтрансферазы. Перенося сахара в широкий спектр вторичных метаболитов, UGT повышают стабильность и растворимость агликонов и, следовательно, изменяют их биологическую активность и эффективность в качестве регуляторов защитного ответа (Lim and Bowles, 2004; Langlois-Meurinne et al., 2005). Семейство GT8 катализирует перенос различных сахаров (Glc, Gal, GlcNAC, GalA) на акцепторы липоолигосахаридов, белков, инозитола, олигосахаридов или полисахаридов с использованием нуклеотидных сахарных субстратов (Yin et al., 2011). Члены семейства участвуют в нескольких различных функциях, включая синтез пектинов и ксилана, а также семейство олигосахаридов рафинозы, которые играют роль в реакции на стресс (Kim et al., 2008). На сегодняшний день GT61, GT75 и GT9Сообщается, что 2 семьи не участвовали в ответных мерах по защите растений. Члены семейства GT61 охарактеризовали функции переноса замен арабинозы и ксилозы на 1,4-β-ксилановый бэкон (Anders et al. , 2012). Члены GT75 аннотированы как мутазы UDP-Ara (UAM), участвующие в превращении UDP-арабинопиранозы в UDP-арабинофуранозу, что необходимо для образования субстрата UDP-Ara f для арабиноксилана, белка арабиногалактана и пектинового полисахарида. биосинтез (Hsieh et al., 2015). С недавним обнаружением арабиноксилана в сосочках ячменя в ответ на попытку проникновения Blumeria graminis f.sp. hordei (Chowdhury et al., 2014), заманчиво предположить, что члены семейства GT61 и GT75 широко вовлечены в защитные реакции. Члены семейства GT92 играют роль в синтезе 1,4-β-галактана (Liwanag et al., 2012), которого относительно много в растянутой древесине, образующейся в ответ на механическое напряжение (Andersson-Gunnerås et al., 2006). . Поэтому, хотя этот общий метаанализ семейств CAZy во время абиотического и биотического стресса не принимает во внимание различия в активности отдельных членов семейства, количестве копий генов или тканеспецифических паттернах экспрессии, он идентифицирует набор семейств CAZy, которые хорошо охарактеризованы с точки зрения реакции на стресс (например, Gh27, Gh28), а также те, которые менее хорошо охарактеризованы (GT61, GT75).
Вопрос о том, оказывают ли члены этих семейств CAZy специфические или сходные эффекты на мишени клеточной стенки, можно определить, охарактеризовав функцию лежащих в их основе генов. Например, повышенная экспрессия семейств GT8 и GT61 подчеркивает потенциальную роль синтеза пектина и ксилана в реакции растений на стресс у обоих видов. Однако эти семейства содержат членов, которые участвуют во многих различных процессах, и важно более подробно оценить экспрессию и функцию каждого гена. На рис. 3 показаны уровни экспрессии каждого гена из семейства генов GT8 арабидопсиса (рис. 3А) и ячменя (рис. 3В). Большинство генов арабидопсиса GT8 активируются в ответ по крайней мере на один стресс, но, по-видимому, существуют подгруппы, которые реагируют на специфические стрессы. И наоборот, семейство GT8 ячменя разделено на две группы: одна содержит гены, которые не меняются или подавляются в ответ на стресс, а другая содержит гены, которые активируются большинством стрессов. Сравнение генов GT8 чувствительного к стрессу ячменя с охарактеризованными членами семейства GT8 от Arabidopsis (рис. 3C) позволяет предположить, что общие гены чувствительного к стрессу ячменя не ограничиваются кладами с единственной предполагаемой функцией, а распределены между галактинолсинтазой (GolS), ксилановой активности глюкуронозилтрансферазы (GUX) и галактуронозилтрансферазы (GAUT и GATL). На рисунке 4 показаны уровни экспрессии каждого гена из семейства генов GT61 арабидопсиса (рис. 4A) и семейства генов GT61 ячменя (рис. 4B). Нет четкой кластеризации генов, чувствительных и не реагирующих на стресс, как это наблюдается для семейства GT8, при этом гены GT61 активируются при ряде различных стрессов как у арабидопсиса, так и у ячменя. Сравнение чувствительных к стрессу генов ячменя и арабидопсиса GT61 с другими членами семейства GT61, которые были функционально охарактеризованы (рис. 4C), показывает, что чувствительные к стрессу гены не ограничиваются кладами с единственной предполагаемой функцией, т. е. β-(1,2)- активности ксилозилтрансферазы (XylT), ксиланксилозилтрансферазы (XXT) и ксиланарабинофуранозилтрансферазы (XAT). Это еще раз подчеркивает необходимость дальнейшей характеристики генов, связанных с клеточной стенкой, в реакциях на стресс. Сохраняющиеся изменения в экспрессии гена CAZy у разных видов могут указывать на то, что родственные гены рекрутируются для действия на сходные субстраты во время стрессовых реакций. Альтернативно, гены из одного и того же семейства могут быть рекрутированы для модификации разных субстратов, но сходным образом.
Рисунок 3. Анализ представителей семейства GT8 после абиотических и биотических стрессов у арабидопсиса (A) и ячменя (B) . Обилие транскриптов определяли путем метаанализа наборов данных микрочипов, собранных из базы данных экспрессии растений (PLEXdb; Dash et al., 2012), с использованием экспериментов, перечисленных в таблице 2. Значения показывают среднее логарифмическое (2)-кратное увеличение индукции для представителей каждого Семейство генов CAZy присутствует на массиве генома ATh2 Arabidopsis Affymetrix 22K ATh2 и на генетическом чипе 22K Barley1. Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. (C) Филогенетическое древо представителей семейства GT8 из арабидопсиса и ячменя с предполагаемыми функциями, присвоенными каждой кладе. Красными точками отмечены гены ячменя, которые активируются в ответ на стресс (B) .
Рисунок 4. Анализ представителей семейства GT61 после абиотических и биотических стрессов у арабидопсиса (A) и ячменя (B) . Обилие транскриптов определяли путем метаанализа наборов данных микрочипов, собранных из базы данных экспрессии растений (PLEXdb; Dash et al., 2012), с использованием экспериментов, перечисленных в таблице 2. Значения показывают среднее логарифмическое (2)-кратное увеличение индукции для представителей каждого Семейство генов CAZy присутствует на массиве генома ATh2 Arabidopsis Affymetrix 22K ATh2 и на генетическом чипе 22K Barley1. Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. (C) Филогенетическое древо членов семейства GT61 из арабидопсиса, ячменя и риса с предполагаемыми функциями, присвоенными каждой кладе.
Перспективы и резюме
Основой для этого обзора было рассмотрение активности клеточных стенок и полисахаридов, которые влияют на биотические и абиотические реакции на стресс, и выделение тех, которые могут выполнять общую функцию, стимулируя ремоделирование клеточной стенки как прямую реакцию. к абиотическому стрессу или атаке патогенов. Генетические и трансгенные данные свидетельствуют о том, что модификация активности специфических клеточных стенок оказывает выраженное влияние на толерантность к стрессу. В некоторых случаях сходные семейства генов, по-видимому, модулируют эффект различных биотических и абиотических стрессов внутри и между разными видами, что подразумевает, что общие механизмы могли быть задействованы для воздействия на, казалось бы, несопоставимые типы стресса. Это подтверждается более широким анализом всего транскриптома, который указывает на сходные ответы отдельных генов, связанных с клеточной стенкой, и даже семейств CAZy на различные абиотические и биотические стрессы. Приводят ли эти перекрытия в экспрессии генов к аналогичным изменениям в структуре клеточных стенок, в большинстве случаев еще предстоит подтвердить, особенно в случае пектинов и ксиланов, которые демонстрируют явные различия в количестве между моделями однодольных и двудольных растений. В самом деле, функции многих генов, связанных с клеточной стенкой, еще не описаны во время нормального роста и развития, не говоря уже о реакциях на стресс. Это подчеркивает необходимость дальнейшего распространения технологий редактирования генома на целые семейства CAZy и разработки высокопроизводительных методологий химического анализа клеточных стенок, способных быть нацеленными на отдельные типы клеток.
Вклад авторов
KH, AL, MT, NS и JC задумали этот обзор, подготовили и отредактировали рукопись, одобрили окончательную версию перед публикацией и соглашаются нести ответственность за все аспекты работы, гарантируя, что вопросы, связанные с точность или целостность любой части работы должным образом исследованы и решены.
Финансирование
KH благодарит за финансирование исследовательской программы правительства Шотландии. AL, NS и JC были поддержаны грантами Австралийского исследовательского совета. MT был поддержан ARC Future Fellowship.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Абебе Т., Мелмаи К., Берг В. и Уайз Р. П. (2010). Реакция колоса ячменя на засуху: экспрессия генов в чешуе, цветковом чешуе, ости и семени. Функц. интегр. Геномика 10, 191–205. doi: 10.1007/s10142-009-0149-4
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Aditya, J., Lewis, J., Shirley, N.J., Tan, H.T., Henderson, M., Fincher, G.B., et al. (2015). Динамика заражения злаковыми цистообразными нематодами различается между чувствительными и устойчивыми сортами ячменя и приводит к изменениям уровня (1, 3; 1, 4)-β-глюкана и количества транскриптов гена HvCslF. Новый Фитол . 207, 135–147. doi: 10.1111/nph.13349
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Аист, Дж. Р., и Исраэль, Х. В. (1977). Формирование сосочков: сроки и значение при проникновении Erysiphe graminis hordei в колеоптилы ячменя. Фитопатология 67, 455–461. doi: 10.1094/Phyto-67-455
CrossRef Полный текст | Google Scholar
Albersheim, P., Darvill, A., Roberts, K., Sederoff, R., and Staehelin, A. (2011). Клеточные стенки и взаимодействие растений и микробов: Garland Science . Нью-Йорк, штат Нью-Йорк: Тейлор и Фрэнсис.
Ан, С. Х., Сон, К. Х., Чой, Х. В., Хван, И. С., Ли, С. К., и Хван, Б. К. (2008). Белок-ингибитор метилэстеразы пектина перца CaPMEI1 необходим для противогрибковой активности, устойчивости к базальным заболеваниям и толерантности к абиотическим стрессам. Планта 228, 61–78. doi: 10.1007/s00425-008-0719-z
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Андерс Н. , Уилкинсон М. Д., Лавгроув А., Фриман Дж., Трифона Т., Пеллни Т. К. и др. (2012). Гликозилтрансферазы семейства 61 опосредуют перенос арабинофуранозила на ксилан в травах. проц. Натл. акад. науч. США 109, 989–993. doi: 10.1073/pnas.1115858109
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Андерссон-Гуннерос С., Меллерович Э. Дж., Лав Дж., Сегерман Б., Омия Ю., Коутиньо П. М. и др. (2006). Биосинтез обогащенной целлюлозой натяжной древесины у Populus: глобальный анализ транскриптов и метаболитов идентифицирует биохимические регуляторы и регуляторы развития в биосинтезе вторичной стенки. Завод Ж. 45, 144–165. doi: 10.1111/j.1365-313X.2005.02584.x
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Аткинсон, Нью-Джерси, Лилли, Си-Джей, и Урвин, П.Е. (2013). Выявление генов, участвующих в реакции арабидопсиса на одновременные биотические и абиотические стрессы. Завод физиол. 162, 2028–2041 гг. doi: 10.1104/pp.113.222372
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Атмоджо, М. А., Хао, З. и Монен, Д. (2013). Эволюция взглядов на биосинтез пектина. год. Преподобный завод биол. 64, 747–779. doi: 10.1146/annurev-arplant-042811-105534
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Беллинкампи Д., Червоне Ф. и Лионетти В. (2014). Динамика клеточных стенок растений и связанная со стенками восприимчивость к взаимодействиям растений и патогенов. Фронт. Растениевод. 5:228. doi: 10.3389/fpls.2014.00228
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бхаттараи К.К., Атамиан Х.С., Калошян И. и Юлгем Т. (2010). 9Факторы транскрипции типа 1100 WRKY72 вносят вклад в базальный иммунитет у томата и арабидопсиса, а также в резистентность между генами, опосредованную R-геном Mi-1 томата. Завод Ж. 63, 229–240. doi: 10.1111/j.1365-313X.2010.04232.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бхуиян Н. Х., Селварадж Г., Вей Ю. и Кинг Дж. (2009). Профилирование экспрессии генов и замалчивание показывают, что биосинтез монолигнола играет критическую роль в защите пшеницы от проникновения мучнистой росы. Дж. Экспл. Бот. 60, 509–521. doi: 10.1093/jxb/ern290
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бодд Дж., Чо С., Крюгер В. М. и Мюльбауэр Г. Дж. (2006). Транскриптомный анализ взаимодействия ячменя и Fusarium graminearum. мол. Взаимодействие растительных микробов. 19, 407–417. doi: 10.1094/MPMI-19-0407
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Болманн Х. и Собчак М. (2014). Клеточная стенка растений в местах питания цистообразующих нематод. Фронт. Растениевод. 5:89. doi: 10.3389/fpls.2014.00089
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Боллер, Т., и Феликс, Г. (2009). Возрождение элиситоров: восприятие связанных с микробами молекулярных паттернов и сигналов опасности рецепторами распознавания паттернов. год. Преподобный завод биол. 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бертон, Р. А., Гидли, М. Дж., и Финчер, Дж. Б. (2010). Неоднородность химического состава, строения и функции клеточных стенок растений. нац. хим. биол. 6, 724–732. doi: 10.1038/nchembio.439
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бертон Р. А., Уилсон С. М., Хрмова М., Харви А. Дж., Ширли Н. Дж., Медхерст А. и др. (2006). Целлюлозосинтазоподобные гены CslF опосредуют синтез (1, 3; 1, 4)-β-D-глюканов клеточной стенки. Наука 311, 1940–1942. doi: 10.1126/science.1122975
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кэл А. Дж., Лю Д., Молеон Р., Син Ю. И. К. и Серрадж Р. (2013). Транскриптомное профилирование зоны удлинения листьев в условиях засухи у контрастных сортов риса. PLoS ONE 8:e54537. doi: 10. 1371/journal.pone.0054537
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Каньо-Дельгадо А., Пенфилд С., Смит К., Кэтли М. и Беван М. (2003). Снижение синтеза целлюлозы вызывает лигнификацию и защитные реакции у Arabidopsis thaliana . Завод Ж. 34, 351–362. doi: 10.1046/j.1365-313X.2003.01729.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чен М. Х., Шэн Дж., Хинд Г., Ханда А. К. и Цитовски В. (2000). Взаимодействие между белком перемещения вируса табачной мозаики и пектинметилэстеразами клетки-хозяина необходимо для перемещения вируса от клетки к клетке. EMBO J. 19, 913–920. doi: 10.1093/emboj/19.5.913
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чен З., Хун Х., Чжан Х., Ван Ю., Ли Х., Чжу Дж. К. и др. (2005). Нарушение гена синтазы целлюлозы, AtCesA8/IRX1, повышает устойчивость арабидопсиса к засухе и осмотическому стрессу. Завод Ж. 43, 273–283. doi: 10.1111/j.1365-313X.2005.02452.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чо, С. К., Ким, Дж. Э., Парк, Дж. А., Эом, Т. Дж., и Ким, В. Т. (2006). Конститутивное выражение индуцируемого абиотическим стрессом острого перца CaXTh4 , который кодирует гомолог ксилоглюкан эндотрансглюкозилазы/гидролазы, улучшает засухоустойчивость и солеустойчивость трансгенных растений арабидопсиса. ФЕБС Письмо. 580, 3136–3144. doi: 10.1016/j.febslet.2006.04.062
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чой, Дж. Ю., Сео, Ю. С., Ким, С. Дж., Ким, В. Т., и Шин, Дж. С. (2011). Конститутивная экспрессия CaXTh4, ксилоглюкановой эндотрансглюкозилазы/гидролазы острого перца, повышала устойчивость растений томата к солевым и засушливым стрессам без фенотипических дефектов (9).1100 Solanum lycopersicum сорт. Дотаранг). Представитель растительных клеток 30, 867–877. doi: 10.1007/s00299-010-0989-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чоудхури Дж. , Хендерсон М., Швейцер П., Бертон Р. А., Финчер Г. Б. и Литтл А. (2014). Дифференциальное накопление каллозы, арабиноксилана и целлюлозы в непроникающих и проникающих сосочках на листьях ячменя, зараженных Blumeria graminis f.sp. хорд. Новый Фитол. 204, 650–660. doi: 10.1111/nph.12974
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Коулбрук Э. Х., Крейссен Г., МакГранн Г. Р. Д., Дреос Р., Лэмб К. и Бойд Л. А. (2012). Приобретенная устойчивость широкого спектра действия у ячменя, индуцированная патосистемой Pseudomonas, имеет общие транскрипционные компоненты с системной приобретенной устойчивостью Arabidopsis. мол. Взаимодействие растительных микробов. 25, 658–667. doi: 10.1094/MPMI-09-11-0246
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Коллмер Н. и Кин Т. (1986). Роль пектиновых ферментов в патогенезе растений. год. Преподобный Фитопат. 24, 383–409. doi: 10.1146/annurev. py.24.0
CrossRef Full Text | Google Scholar
Коминелли Э., Сала Т., Кальви Д., Гусмароли Г. и Тонелли К. (2008). Сверхэкспрессия гена AtMYB41 Arabidopsis изменяет рост клеток и проницаемость поверхности листа. Завод Ж. 53, 53–64. doi: 10.1111/j.1365-313X.2007.03310.x
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Coolen, S., Proietti, S., Hickman, R., Davila Olivas, N. H., Huang, P.-P., Van Verk, M. C., et al. (2016). Динамика транскриптома арабидопсиса при последовательных биотических и абиотических стрессах. Завод Ж. 86, 249–267. doi: 10.1111/tpj.13167
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Cosgrove, DJ (2005). Рост клеточной стенки растений. Нац. Преподобный Мол. Клеточная биол. 6, 850–861. doi: 10.1038/nrm1746
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Craigon, D.J., James, N., Okyere, J., Higgins, J. , Jotham, J., and May, S. (2004). NASCArrays: репозиторий данных микрочипов, созданных службой транскриптомики NASC. Рез. нуклеиновых кислот. 32, Д575–Д577. doi: 10.1093/nar/gkh233
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Cu, S., Collins, H.M., Betts, N.S., March, T.J., Janusz, A., Stewart, D.C., et al. (2016). Поглощение воды зерном ячменя: физиология; генетика и промышленное применение. Растениеводство . 242, 260–269. doi: 10.1016/j.plantsci.2015.08.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Катлер Дж. М., Шахан К. В. и Степонкус П. Л. (1980). Влияние водного дефицита и осмотической адаптации на удлинение листьев риса. Растениеводство. 20, 314–318. doi: 10.2135/cropsci1980.0011183X002000030006x
CrossRef Full Text | Google Scholar
Дэш С., Ван Хемерт Дж., Хонг Л., Уайз Р. П. и Дикерсон Дж. А. (2012). PLEXdb: ресурсы экспрессии генов для растений и фитопатогенов. Рез. нуклеиновых кислот. 40, Д1194–Д1201. doi: 10.1093/nar/gkr938
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дэвис, Л.Дж., и Урвин, П.Е. (2012). Клеточные стенки синцитиев, образованные Heterodera schachtii в Arabidopsis thaliana , богаты метилэтерифицированным пектином. Сигнал завода. Поведение 7, 1404–1406. doi: 10.4161/psb.21925
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Де Вос, М., Ван Остен, В. Р., Ван Поке, Р. М., Ван Пелт, Дж. А., Позо, М. Дж., Мюллер, М. Дж., и др. (2005). Изменения сигнальной сигнатуры и транскриптома арабидопсиса во время атаки патогенов и насекомых. мол. Взаимодействие растительных микробов. 18, 923–937. doi: 10.1094/MPMI-18-0923
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дельгадо-Сересо М., Санчес-Родригес К., Эскудеро В., Миедес Э., Фернандес П.В., Хорда Л. и др. (2012). Гетеротримерный G-белок арабидопсиса регулирует защиту клеточной стенки и устойчивость к некротрофным грибам. мол. Завод. 5, 98–114. doi: 10.1093/mp/ssr082
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Дельп, Г., Градин, Т., Ахман, И., и Йонссон, Л.М. (2009). Микроматричный анализ взаимодействия между тлей Rhopalosiphum padi и растениями-хозяевами выявляет как различия, так и сходства между восприимчивыми и частично устойчивыми линиями ячменя. мол. Жене. Геномика 281, 233–248. doi: 10.1007/s00438-008-0409-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Денансе Н., Раноча П., Ориа Н., Барле X., Ривьер М.П., Ядета К.А. и др. (2013). Арабидопсис wat1 ( стенки тонкие1 )-опосредованная резистентность к бактериальному сосудистому патогену Ralstonia solanacearum сопровождается перекрестной регуляцией метаболизма салициловой кислоты и триптофана. Завод Ж. 73, 225–239. doi: 10.1111/tpj.12027
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Деуберт, К. Х., и Роде, Р. А. (1971). «Ферменты нематод», в Паразитические нематоды растений: цитогенетика, взаимодействия хозяин-паразит и физиология , Том. 2, ред. Б. М. Цукерман, В. Ф. Май и Р. А. Роде (Нью-Йорк, штат Нью-Йорк: Academic Press), 73–90.
Google Scholar
Дюру Л., Дельмотт Ф. М., Ланселин Дж.-М., Керавис Г. и Джей-Аллеанд К. (1998). Взгляд на метаболизм нафтохинона: катализируемый β-глюкозидазой гидролиз гидроюглона β-D-глюкопиранозида. Биохим. J. 333, 275–283. doi: 10.1042/bj3330275
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Эйзен, М. Б., Спеллман, П. Т., Браун, П. О., и Ботштейн, Д. (1998). Кластерный анализ и отображение полногеномных паттернов экспрессии. проц. Натл. акад. науч. США 95, 14863–14868. doi: 10.1073/pnas.95.25.14863
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Эллингер Д., Науманн М., Фальтер К., Звикович К., Джамроу Т., Маниссери К. и др. (2013). Повышенное раннее отложение каллозы приводит к полной устойчивости арабидопсиса к проникновению мучнистой росы. Завод физиол. 161, 1433–1444. doi: 10.1104/pp.112.211011
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Эллис, К., и Тернер, Дж. Г. (2001). Мутант арабидопсиса cev1 имеет конститутивно активные жасмонатные и этиленовые сигнальные пути и повышенную устойчивость к патогенам. Растительная клетка 13, 1025–1033. doi: 10.1105/tpc.13.5.1025
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фэн Л., Линкер Р., Гепштейн С., Танимото Э., Ямамото Р. и Нойманн П. М. (2006). Прогрессирующее ингибирование водным дефицитом растяжимости и роста клеточных стенок вдоль зоны растяжения корней кукурузы связано с повышенным метаболизмом лигнина и прогрессирующим накоплением фенольных смол в стенках. Завод Физиол. 140, 603–612. doi: 10.1104/pp.105.073130
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Феррари С. , Саватин Д. В., Сицилия Ф., Граменья Г., Червоне Ф. и Де Лоренцо Г. (2013). Олигогалактурониды: молекулярные паттерны, связанные с повреждением растений, и регуляторы роста и развития. Фронт. Растениевод. 4:49. doi: 10.3389/fpls.2013.00049
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фрай, С. К., Смит, Р. К., Ренвик, К. Ф., Мартин, Д. Дж., Ходж, С. К., и Мэтьюз, К. Дж. (1992). Ксилоглюкан эндотрансгликозилаза, новый фермент, разрыхляющий стенки растений. Биохим. Дж. 282, 821–828. doi: 10.1042/bj2820821
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Gehan, M.A., Greenham, K., Mockler, T.C., and McClung, C.R. (2015). Транскрипционные сети — урожай, часы и абиотический стресс. Курс. мнение Растение Биол . 24, 39–46. doi: 10.1016/j.pbi.2015.01.004
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Гонсалес-Перес, С., Гутьеррес, Дж., Гарсия-Гарсия, Ф. , Осуна, Д., Допазо, Дж., Лоренцо, О., и др. (2011). Ранние защитные реакции транскрипции в культуре суспензии клеток арабидопсиса в условиях яркого света. Завод физиол. 156, 1439–1456. doi: 10.1104/pp.111.177766
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хаманн, Т. (2012). Поддержание целостности клеточных стенок растений как важнейший компонент механизмов реакции на биотический стресс. Фронт. Растениевод. 3:77. doi: 10.3389/fpls.2012.00077
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Харб А., Кришнан А., Амбаварам М. М. и Перейра А. (2010). Молекулярно-физиологический анализ стресса от засухи у арабидопсиса выявляет ранние реакции, ведущие к акклиматизации роста растений. Завод физиол. 154, 1254–1271. doi: 10.1104/pp.110.161752
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Эрнандес-Бланко, К., Фэн, Д. X., Ху, Дж., Санчес-Валлет, А., Десландес, Л. , Льоренте, Ф., и др. (2007). Нарушение синтазы целлюлозы, необходимой для образования вторичной клеточной стенки арабидопсиса, повышает устойчивость к болезням. Растительная клетка 19, 890–903. doi: 10.1105/tpc.106.048058
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хофманн Дж., Юссеф-Банора М., де Алмейда-Энглер Дж. и Грундлер Ф. М. (2010). Роль отложения каллозы вдоль плазмодесм в местах питания нематод. мол. Взаимодействие растительных микробов. 23, 549–557. doi: 10.1094/MPMI-23-5-0549
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хок С., Аттард А. и Келлер Х. (2010). Получение максимума от хозяина: как патогены заставляют растения сотрудничать при болезни. мол. Взаимодействие растительных микробов. 23, 1253–1259. doi: 10.1094/MPMI-04-10-0103
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Се Ю. С., Чжан К., Яп К., Ширли Н. Дж., Ланштайн Дж. , Нельсон С. Дж. и др. (2015). Генетика, профили транскрипции и каталитические свойства семейства udp-арабинозомутазы из ячменя. Биохимия 55, 322–334. doi: 10.1021/acs.biochem.5b01055
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Хукельховен, Р. (2005). Восприимчивость к мучнистой росе и стратегии биотрофического заражения. FEMS микробиол. лат. 245, 9–17. doi: 10.1016/j.femsle.2005.03.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Иноуэ, С., Аист, Дж. Р., и Мако, В. (1994). Более раннее образование сосочков и устойчивость к мучнистой росе ячменя, индуцированные экстрактом, регулирующим сосочков. Физиол. Мол. Завод Патол. 44, 433–440. дои: 10.1016/S0885-5765(05)80099-2
Полнотекстовая перекрестная ссылка | Google Scholar
Джейкобс А. К., Липка В., Бертон Р. А., Панструга Р., Стрижов Н., Шульце-Леферт П. и др. (2003). Синтаза каллозы Arabidopsis, GSL5 , необходима для образования мозолей в ранах и сосочках. Растительная клетка. 15, 2503–2513. doi: 10.1105/tpc.016097
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дженсен М. К., Хагедорн П. Х., Торрес-Забала Д., Грант М. Р., Рунг Дж. Х., Коллиндж Д. Б. и др. (2008). Регуляция транскрипции фактором транскрипции NAC (NAM-ATAF1, 2-CUC2) ослабляет передачу сигналов ABA для эффективной базальной защиты от Blumeria graminis f.sp. hordei у арабидопсиса. Завод J. 56, 867–880. doi: 10.1111/j.1365-313X.2008.03646.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джонс, Дж. Д. Г., и Дангл, Дж. Л. (2006). Иммунная система растений. Природа 444, 323–329. doi: 10.1038/nature05286
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кемпема, Л. А., Цуй, X., Хольцер, Ф. М., и Уоллинг, Л. Л. (2007). Транскриптом арабидопсиса изменяется в ответ на питание нимф белокрылки серебристолистной флоэмой. сходства и различия в реакциях на тлю. Завод Физиол. 143, 849–865. doi: 10.1104/pp.106.0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ким М.С., Чо С.М., Канг Э.Ю., Им Ю.Дж., Хванбо Х., Ким Ю.К. и др. (2008). Галактинол является сигнальным компонентом индуцированной системной резистентности, вызванной колонизацией корня Pseudomonas chlororaph is O6. мол. Взаимодействие растительных микробов. 21, 1643–1653. doi: 10.1094/MPMI-21-12-1643
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Кимпара Т., Аохара Т., Сога К., Вакабаяси К., Хосон Т., Цумурая Ю. и др. (2008). Активность β-1,3:1,4-глюкансинтазы в проростках риса под водой. Энн. Бот. 102, 221–226. doi: 10.1093/aob/mcn077
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кумар М., Кэмпбелл Л. и Тернер С. (2016). Вторичные клеточные стенки: биосинтез и манипуляции. Дж. Экспл. Бот. 67, 515–531. doi: 10.1093/jxb/erv533
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Квон, Ю. , Ким, С. Х., Юнг, М. С., Ким, М. С., О, Дж. Э., Джу, Х. В. и др. (2007). Arabidopsis hot2 кодирует эндохитиназоподобный белок, необходимый для устойчивости к жаре, соли и засухе. Завод Ж. 49, 184–193. doi: 10.1111/j.1365-313X.2006.02950.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Langlois-Meurinne, M., Gachon, CMM, and Saindrean, P. (2005). Патоген-чувствительная экспрессия генов гликозилтрансфераз UGT73B3 и UGT73B5 необходима для устойчивости к Pseudomonas syringae pv томат в арабидопсисе. Завод физиол. 139, 1890–1901 гг. doi: 10.1104/pp.105.067223
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ле Галль Х., Филипп Ф., Домон Дж. М., Жилле Ф., Пеллу Дж. и Район К. (2015). Метаболизм клеточной стенки в ответ на абиотический стресс. Растения 4, 112–166. doi: 10.3390/plants4010112
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лейбнер-Мецгер, Г. , и Мейнс, Ф. Дж. (1999). «Функции и регуляция растительных β-1,3-глюканаз (PR-2)», в «Связанные с патогенезом белки в растениях », редакторы С. Датта и С. Мутукришнан (Флорида, Флорида: CRC Press LLC Boca Raton), 49–76.
Google Scholar
Li, H., Yan, S., Zhao, L., Tan, J., Zhang, Q., Gao, F., et al. (2014). Активация генов, связанных с клеточной стенкой, связанная с ацетилированием гистонов, участвует в набухании корней кукурузы, вызванном солевым стрессом. BMC Растение Биол. 14:105. дои: 10.1186/1471-2229-14-105
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Лим, Э. К., и Боулз, Д. Дж. (2004). Класс растительных гликозилтрансфераз, участвующих в клеточном гомеостазе. EMBO J. 23, 2915–2922. doi: 10.1038/sj.emboj.7600295
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лионетти В., Червоне Ф. и Беллинкампи Д. (2012). Метиловая этерификация пектина играет роль во взаимодействиях растений и патогенов и влияет на устойчивость растений к болезням. J. Физиол растений. 169, 1623–1630. doi: 10.1016/j.jplph.2012.05.006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лю Х., Ма Ю., Чен Н. А., Го С., Лю Х., Го X. и др. (2014). Сверхэкспрессия индуцируемого стрессом OsBURP16 , β-субъединицы полигалактуроназы 1, снижает содержание пектина и адгезию клеток и повышает чувствительность к абиотическому стрессу у риса. Окружающая среда растительных клеток. 37, 1144–1158. doi: 10.1111/pce.12223
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Liwanag, A.J.M., Ebert, B., Verhertbruggen, Y., Rennie, E.A., Rautengarten, C., Oikawa, A., et al. (2012). Биосинтез пектина: GALS1 в Arabidopsis thaliana представляет собой β-1,4-галактан β-1,4-галактозилтрансферазу. Растительная клетка 24, 5024–5036. doi: 10.1105/tpc.112.106625
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ломбард В., Голаконда Рамулу Х. , Друла Э., Коутиньо П. М. и Хенриссат Б. (2014). База данных углеводно-активных ферментов (CAZy) в 2013 г. Рез. нуклеиновых кислот. 42, Д490–Д495. doi: 10.1093/nar/gkt1178
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Малиновский Ф.Г., Фангель Дж.У. и Уиллатс В.Г. (2014). Роль клеточной стенки в иммунитете растений. Фронт. Растениевод. 5:178. doi: 10.3389/fpls.2014.00178
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Manabe, Y., Nafisi, M., Verhertbruggen, Y., Orfila, C., Gille, S., Rautengarten, C., et al. (2011). Мутация с потерей функции СНИЖЕНИЕ АЦЕТИЛИРОВАНИЯ СТЕНКИ2 у Arabidopsis приводит к снижению ацетилирования клеточной стенки и повышению устойчивости к Botrytis cinerea . Завод физиол. 155, 1068–1078. doi: 10.1104/pp.110.168989
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мангельсен Э., Килиан Дж., Хартер К., Янссон К. , Ванке Д. и Сандберг Э. (2011). Транскриптомный анализ высокотемпературного стресса в развивающихся зерновках ячменя: ранние реакции на стресс и влияние на биосинтез запасных соединений. мол. Завод 4, 97–115. doi: 10.1093/mp/ssq058
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
МакГранн Г. Р., Таунсенд Б. Дж., Антонив Дж. Ф., Ашер М. Дж. и Мутаса-Гёттгенс Э. С. (2009). Ячмень вызывает аналогичную раннюю базальную защитную реакцию во время взаимодействия хозяина и не хозяина с корневыми паразитами Polymyxa. евро. Дж. Плант Патол. 123, 5–15. doi: 10.1007/s10658-008-9332-z
CrossRef Полный текст | Google Scholar
Миллет Б. П., Сюн Ю., Даль С. К., Стеффенсон Б. Дж. и Мюльбауэр Г. Дж. (2009 г.)). Дикий ячмень накапливает разные наборы транскриптов в ответ на патогены с разным трофическим образом жизни. Физиол. Мол. Завод Патол . 74, 91–98. doi: 10.1016/j.pmpp.2009.09.006
CrossRef Full Text | Google Scholar
Минич, З. (2008). Физиологическая роль растительных гликозидгидролаз. Планта 227, 723–740. doi: 10.1007/s00425-007-0668-y
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мершбахер Б. и Мендген К. (2012). «Структурный аспект защиты растений», в Механизмы устойчивости к болезням растений , ред. А. Дж. Слюсаренко, Р. С. Фрейзер и Л. К. ван Лун (Берлин: Springer Science & Business Media), 231–277.
Google Scholar
Мур, Дж. П., Викре-Гибуэн, М., Фаррант, Дж. М., и Дриуич, А. (2008). Адаптация клеточных стенок высших растений к потере воды: засуха против высыхания. Физиол. Завод . 134, 237–245. doi: 10.1111/j.1399-3054.2008.01134.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Mouille, G., Robin, S., Lecomte, M., Pagant, S., and Höfte, H. (2003). Классификация и идентификация мутантов клеточной стенки арабидопсиса с помощью микроспектроскопии с преобразованием Фурье в инфракрасном диапазоне (FT-IR). Завод Ж. 35, 393–404. doi: 10.1046/j.1365-313X.2003.01807.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нафиси М., Фимогнари Л. и Сакураги Ю. (2015). Взаимодействия между клеточной стенкой и фитогормонами при взаимодействии растений и некротрофных патогенов. Фитохимия 112, 63–71. doi: 10.1016/j.phytochem.2014.11.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нисимура, М. Т., Штейн, М., Хоу, Б. Х., Фогель, Дж. П., Эдвардс, Х., и Сомервилль, С. К. (2003). Потеря синтазы каллозы приводит к устойчивости к салициловой кислоте. Наука 301, 969–972. doi: 10.1126/science.1086716
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Оз М. Т., Йилмаз Р., Эйдоган Ф., Де Грааф Л. и Юсел М. Октем Х. А. (2009 г.)). Микроматричный анализ поздней реакции листьев ячменя ( Hordeum vulgare L.) на токсичность бора. тюрк. Дж. Агрик. За. 33, 191–202. doi: 10.3906/tar-0806-22
Полный текст CrossRef | Google Scholar
Поли М., Гилле С., Лю Л., Мансури Н., де Соуза А., Шултинк А. и др. (2013). Биосинтез гемицеллюлозы. Планта 238, 627–642. doi: 10.1007/s00425-013-1921-1
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Пеллу, Дж., Рустеруччи, К., и Меллерович, Э. Дж. (2007). Новое понимание структуры и функции пектинметилэстеразы. Trends Plant Sci. 12, 267–277. doi: 10.1016/j.tplants.2007.04.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Poulton, JE (1990). Цианогенез у растений. Завод физиол. 94, 401–405. doi: 10.1104/pp.94.2.401
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пураник С., Саху П. П., Шривастава П. С. и Прасад М. (2012). Белки NAC: регуляция и роль в стрессоустойчивости. Trends Plant Sci. 17, 369–381. doi: 10.1016/j.tplants.2012.02.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Рай К. М., Баласубраманян В.К., Велкер К.М., Панг М., Хии М.М. и Менду В. (2015). Комплексный анализ всего генома и разработка веб-ресурсов о ферментах, разрушающих клеточную стенку фитопаразитических нематод. BMC Растение Биол. 15:187. doi: 10.1186/s12870-015-0576-4
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Раксеги М., Лавгроув А., Балла К., Ланг Л., Бедо З., Вейс О. и др. (2014). Влияние теплового и засушливого стресса на структуру и состав арабиноксилана и β-глюкана в зерне пшеницы. Углеводы Полим. 102, 557–565. doi: 10.1016/j.carbpol.2013.12.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ренни, Э. А., и Шеллер, Х. В. (2014). Биосинтез ксилана. Курс. мнение Биотехнолог. 26, 100–107. doi: 10.1016/j.copbio.2013.11.013
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Ringli, C. (2010). Мониторинг снаружи: механизмы восприятия клеточной стенки. Завод физиол. 153, 1445–1452. doi: 10.1104/pp.110.154518
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Роуз Дж. К., Браам Дж., Фрай С. К. и Нишитани К. (2002). Семейство ферментов XTH, участвующих в эндотрансглюкозилировании и эндогидролизе ксилоглюкана: современные перспективы и новая объединяющая номенклатура. Физиол клеток растений. 43, 1421–1435. doi: 10.1093/pcp/pcf171
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сантос А. П., Серра Т., Фигейредо Д. Д., Баррос П., Лоренсу Т., Чандер С. и др. (2011). Транскрипционная регуляция реакции на абиотический стресс у риса: комбинированное действие факторов транскрипции и эпигенетических механизмов. ОМИКС 15, 839–857. doi: 10.1089/omi.2011.0095
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Шеллер, Х.В., и Ульвсков, П. (2010). Гемицеллюлозы. год. Преподобный завод биол. 61, 263–289. doi: 10.1146/annurev-arplant-042809-112315
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Секи М. , Нарусака М., Исида Дж., Нанджо Т., Фудзита М., Ооно Ю. и др. (2002). Мониторинг профилей экспрессии 7000 генов арабидопсиса в условиях засухи, холода и сильного засоления с использованием полноразмерного микрочипа кДНК. Завод Ж. 31, 279–292. doi: 10.1046/j.1365-313X.2002.01359.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Стоун, Б. А., и Кларк, А. Э. (1992). Химия и биология (1 → 3)-β-D-глюканов . Мельбурн, Виктория: Издательство Университета Ла Троб.
Google Scholar
Свенссон Дж. Т., Кросатти К., Камполи К., Басси Р., Станка А. М., Клоуз Т. Дж. и др. (2006). Транскриптомный анализ адаптации к холоду у мутантов ячменя Albina и Xantha. Завод физиол. 141, 257–270. doi: 10.1104/pp.105.072645
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Суонстон Дж. С., Эллис Р. П., Перес-Вендрель А., Вольтас Дж. и Молина-Кано Дж. Л. (1997). Закономерности развития зерна ячменя в Испании и Шотландии и их влияние на качество солода. Зерновые хим. 74, 456–461. doi: 10.1094/CCHEM.1997.74.4.456
Полный текст CrossRef | Google Scholar
Такета С., Юо Т., Тонука Т., Цумурая Ю., Инагаки Ю., Харуяма Н. и др. (2012). Функциональная характеристика бетаглюкановых мутантов ячменя демонстрирует уникальную роль CsF6 в биосинтезе (1, 3; 1, 4)-β-D-глюкана. Дж. Экспл. Бот. 63, 381–392. doi: 10.1093/jxb/err285
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Taylor-Teeples, M., Lin, L., de Lucas, M., Turco, G., Toal, T.W., Gaudinier, A., et al. (2015). Регуляторная сеть генов арабидопсиса для синтеза вторичной клеточной стенки. Природа 517, 571–575. doi: 10.1038/nature14099
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Тенхакен, Р. (2014). Ремоделирование клеточной стенки при абиотическом стрессе. Фронт. Растениевод. 5:771. doi: 10.3389/fpls.2014.00771
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Такер, М. Р., и Колтунов, А. М. (2014). Мониторы трафика на периферии клетки: роль клеточных стенок во время ранней дифференцировки женских половых клеток у растений. Курс. мнение биол. растений 17, 137–145. doi: 10.1016/j.pbi.2013.11.015
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Тернер С. Р. и Сомервилл С. Р. (1997). Фенотип коллапсированной ксилемы арабидопсиса идентифицирует мутантов с дефицитом отложения целлюлозы во вторичной клеточной стенке. Растительная клетка 9, 689–701. doi: 10.1105/tpc.9.5.689
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Турра Д., Эль Галид М., Росси Ф. и Ди Пьетро А. (2015). Грибной патоген использует рецептор полового феромона для хемотропного восприятия сигналов растения-хозяина. Природа 527, 521–524. doi: 10.1038/nature15516
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
ван Дам, Нью-Мексико (2009). Подземные травоядные и защита растений. год. Преподобный Экол. Эвол. Сист. 40, 373–391. doi: 10.1146/annurev.ecolsys.110308.120314
Полный текст CrossRef | Google Scholar
Vandenborre, G., Smaghe, G., and Van Damme, EJ (2011). Лектины растений как защитные белки от насекомых-фитофагов. Фитохимия 72, 1538–1550. doi: 10.1016/j.phytochem.2011.02.024
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Ван Кан, Дж. А. (2006). Лицензия на убийство: образ жизни некротрофного патогена растений. Trends Plant Sci. 11, 247–253. doi: 10.1016/j.tplants.2006.03.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Вега-Санчес, М. Э., Верхертбрюгген, Ю., Кристенсен, У., Чен, X., Шарма, В., Варанаси, П., и др. (2012). Потеря функции F6 , подобной целлюлозосинтазе , влияет на отложение глюкана со смешанной связью, механические свойства клеточной стенки и защитные реакции в вегетативных тканях риса. Завод Физиол. 159, 56–69. doi: 10.1104/pp.112.195495
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Vogel, JP, Raab, T.K., Somerville, CR, and Somerville, S.C. (2002). PMR6 , ген, подобный пектатлиазе, необходимый для восприимчивости арабидопсиса к мучнистой росе. Растительная клетка 14, 2095–2106. doi: 10.1105/tpc.003509
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Vogel, JP, Raab, T.K., Somerville, CR, and Somerville, SC (2004). Мутации в PMR5 приводят к устойчивости к мучнистой росе и изменению состава клеточных стенок. Завод J. 40, 968–978. doi: 10.1111/j.1365-313X.2004.02264.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уоллворк, М. А. Б., Дженнер, К. Ф., Лог, С. Дж., и Седжли, М. (1998). Влияние высокой температуры при наливе зерна на структуру формирующегося и соложеного зерна ячменя. Энн. Бот. 82, 587–599. doi: 10.1006/anbo.1998.0721
Полный текст CrossRef | Академия Google
Ван Т. , Макфарлейн Х. Э. и Перссон С. (2016). Влияние абиотических факторов на синтез целлюлозы. Дж. Экспл. Бот. 67, 543–552. doi: 10.1093/jxb/erv488
PubMed Abstract | CrossRef Full Text
Wang, Y., Yang, L., Zheng, Z., Grumet, R., Loescher, W., Zhu, J.K., et al. (2013). Транскриптомные и физиологические вариации трех экотипов арабидопсиса в ответ на солевой стресс. PLoS ONE 8:e69036. doi: 10.1371/journal.pone.0069036
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Wieczorek, K. (2015). Глава Трехклеточная стенка пораженных нематодами корней. Доп. Бот. Рез. 73, 61–90. doi: 10.1016/bs.abr.2014.12.002
CrossRef Full Text | Google Scholar
Wieczorek, K., Elashry, A., Quentin, M., Grundler, F.M.W., Favery, B., Seifert, G.J., et al. (2014). Определена роль пектатлиаз в формировании питающих структур, индуцируемых цистными и галловыми нематодами. мол. Взаимодействие растительных микробов. 27, 901–912. doi: 10.1094/MPMI-01-14-0005-R
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ву М. О., Бирд Х., Макдональд М. Х., Брюэр Э. П., Юссеф Р. М., Ким Х. и др. (2014). Манипуляции с двумя генами α-эндо-β-1,4-глюканазы, AtCel6 и GmCel7, снижают восприимчивость корней сои к глицинам Heterodera. мол. Завод Патол. 15, 927–939. doi: 10.1111/mpp.12157
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Ву, Ю., и Косгроув, Д. Дж. (2000). Адаптация корней к низкому водному потенциалу за счет изменения растяжимости клеточной стенки и белков клеточной стенки. Дж. Экспл. Бот. 51, 1543–1553. doi: 10.1093/jexbot/51.350.1543
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Инь Ю. Б., Монен Д., Гелинео-Альбершайм И., Сюй Ю. и Хан М. Г. (2011). Гликозилтрансферазы семейства GT8. год. Заводской номер . 41, 167–211. дои: 10.1002/97814443
.ch6
Полнотекстовая перекрестная ссылка | Google Scholar
Зентмайер, Г.