Развивать корень: Страница не найдена
Корень слова. 2-й класс
- Паночкина Ольга Александровна, учитель начальных классов
Разделы: Начальная школа
Класс: 2
Цели:
- познакомить с термином «корень» слова и «однокоренные слова»;
- формировать умение выделять корень в родственных словах;
- развивать умение анализировать, сравнивать, делать выводы;
- развивать навыки самоконтроля, познавательные процессы и орфографическую зоркость.
- воспитывать аккуратность, усидчивость чувство коллективизма.
ХОД УРОКА
1. Организационный момент
Организационный момент
Итак, друзья, внимание –
Ведь прозвенел звонок.
Садитесь поудобнее
Пора начать урок!
Тетрадь правильно кладу,
Ручку правильно держу,
За посадкою слежу
И красиво напишу!
(Запись числа, классной работы).
– Вспомните о каких признаках родственных слов рассказал Грамматик (аппликация на доске). Помня эти признаки, поиграем в игру «Где родственные слова?» (слова записаны на доске).
1 ряд:
1) Лист, лиса, лисёнок.
2) Лось, лоскут, лосёнок.
2 ряд:
3) Горный, город, городок.
4) Моряки, морской, моржи.
– Выпишите из каждой строки только родственные
слова. (Самостоятельно).
– Проверим и докажем правильность ответа (ребята
показывают сигнальные карточки, а учитель
стирает лишние слова).
2. Постановка цели урока
– Общей части родственных слов и будет посвящён наш урок: вы узнаете как она называется по-другому и обозначается в словах.
3. Изучение нового материала
Инсценировка «Два корня» (рисунок растения, шапочка с обозначением корня).
Встретились корень куста и корень слова.
– Здравствуй. Я корень, а ты кто?
– Я тоже корень.
– Я в земле живу, а ты где?
– А я живу в словах.
– Ну какой же корень может жить в словах?! Вот я
корень так корень! Посмотри: от меня в земле берут
начало ростки и вырастает цветок – одуванчик,
дерево – берёза, кустарник – шиповник
– Не хвастайся. От меня и других таких же корней,
как я, тоже вырастают целые кусты, только не
растений, а новых слов.
– Подумаешь! Зато на кустах и деревьях, которые
от меня вырастают, зреют вкусные фрукты и ягоды,
которые можно съесть. А твоих слов не съешь!
– А без слов, которые от меня вырастают, ни одного
твоего растения, ни фруктов, ни ягод и назвать-то
нельзя!
Услышали спор люди и говорят:
– Не спорьте, друзья, оба вы нам необходимы. Нужны
нам и такие корни, от которых хорошие растения
вырастают, очень нужны и такие, от которых новые
слова вырастить можно. Как у всех веток куста
есть общий корень, так и у родственных слов
есть общая для всех часть, от которой они
произошли, которая называется – КОРЕНЬ.
– Из уважения к родителю слов родственные слова соорудили домик для корня вот такой округлый и уютный (нарисовать на доске). Давайте найдём корень в наших словах и выделим его в игре «Где родственные слова?»
В корне заключён смысл слова; это главная
часть слова и от неё «растут» слова, они
называются ОДНОКОРЕННЫМИ.
4. Закрепление изученного материала
1) Работа с учебником. С. 46-47
– Прочитайте сообщение Грамматика.
Упр. 8. Прочитайте задание и назовите
первую(вторую) группу родственных слов.Докажите.
– Запишите родственные слова и выделите корень(2
учеников работают за доской).
– Проверим. Что такое корень?
5. Физминутка
2) Игра «Найди корень» (играют по рядам)
Так же, как и у кустов,
Корень слова есть у слов.
Будь внимателен к словам,
Отыщи в них корень сам!
– У каждого кустарника есть корень, но
ветви-слова с ним не соединены. Выделите корень в
слове и соедините с корнем куста. Прочитайте
внимательно слова прежде, чем начать игру.
– Чей ряд быстро и правильно выполнит задание?!
– Какой корень у слов 1 ряда? (-вод-); у
слов 2 ряда? (-мороз-)
– Как называются эти слова? Запишите три любых
однокоренных слова, выделите корень?
6. Итог урока
Итог урока
Корень слова – главная,
Значимая часть.
Родственные связи
Дарят корню власть.
– Что такое корень? Каким значком обозначается
в словах?
– Как называются слова с одним и тем же корнем?
– Чему учились на уроке?
7. Оценивание. Разбор домашнего задания
С.46-47 упр.9
– Прочитайте задание.
– Объясните, от какого слова образовано каждое
слово и что оно называет.
Домашняя – от какого слова образовано? Ставьте цифру 1 и ищите ему однокоренные слова. (Проследить за слабыми учениками).
Литература:
1. Игры и занимательные упражнения на уроках
русского языка 1-4. Н.М. Бетенькова, Д.С. Фонин.
2. Дидактические игры на уроках русского языка в
1-4 кл. вспомогательной школы А. К. Аксёнова, Э.В.
Якубовская.
К. Аксёнова, Э.В.
Якубовская.
3. Учебник русского языка во 2-м классе.
3. Выявление места и причины затруднения.
4. Построение проекта выхода из затруднения.
5. Реализация построенного проекта
6. Первичное закрепление во внешней речи.
7. Самостоятельная работа с самопроверкой по образцу
8. Включение в систему знаний и повторение
9. Рефлексия учебной деятельности.
| Регулятивные: волевая саморегуляция. Коммуникативные: планирование учебного сотрудничества с учителем и со сверстниками. Познавательные: общеучебные: умение структурировать знания, контроль и оценка процесса и результатов деятельности; логические: анализ, синтез, выбор оснований Регулятивные: контроль, коррекция, прогнозирование (при анализе пробного действия перед его выполнением). Познавательные: умение структурировать знания; постановка и формулирование проблемы; умение осознанно и произвольно строить речевое высказывание. Регулятивные: целеполагание как постановка учебной задачи, планирование, прогнозирование. Познавательные: общеучебныевыбор наиболее эффективных способов решение в зависимости от конкретных условий. Коммуникативные: планирование учебного сотрудничества со сверстниками, сотрудничество; управление поведением партнера; умение выражать свои мысли. Познавательные: общеучебные: поиск и выделение необходимой информации; смысловое чтение и логические: построение логической цепи рассуждений, анализ, синтез. УУД постановки и решения проблем: самостоятельное создание способов решения проблем поискового характера. Коммуникативные: умение выражать свои мысли. Регулятивные: контроль в форме сличения способа действия и его результата с заданным эталоном; коррекция; оценка – оценивание качества и уровня усвоения; коррекция. Познавательные: общеучебные: умение осознанно и произвольно строить речевое высказывание Умение доказывать свою точку зрения, пользуясь эталоном (Общий корень, связаны по смыслу) Регулятивные: прогнозирование Познавательные: общенаучные: умение структурировать знания; оценка процесса и результатов деятельности. Коммуникативные: умение выражать свои мысли. Регулятивные: волевая саморегуляция; оценка – выделение и осознание учащимися того, что уже усвоено и что еще подлежит усвоению, прогнозирование. | ||



 к. другие слова обозначают одно лицо, а это слово обозначает два или несколько лиц.
к. другие слова обозначают одно лицо, а это слово обозначает два или несколько лиц. 52 и прочитайте, что там об этом написано.
52 и прочитайте, что там об этом написано.
 А вы докажите(вторая группа), что ваши слова никак не связаны. Работа в группах. Обсуждение.
А вы докажите(вторая группа), что ваши слова никак не связаны. Работа в группах. Обсуждение. Работа водителя не связана с водой). Ребята, какой мы сделаем вывод?
Работа водителя не связана с водой). Ребята, какой мы сделаем вывод? Откройте учебник на странице, номер которой является двузначным числом и состоит из соседних чисел, в сумме составляющих 15.
Откройте учебник на странице, номер которой является двузначным числом и состоит из соседних чисел, в сумме составляющих 15. А сколько у нас в классе человек?
А сколько у нас в классе человек? Запишите эту группу слов в тетради и докажите, родственные это слова или нет.
Запишите эту группу слов в тетради и докажите, родственные это слова или нет.







«Состав слова. Значимые части слова»
Тема: «Состав слова. Значимые части слова»
Цели урока:
Уточнить представление учащихся о признаках однокоренных слов.
Развивать умение распознавать однокоренные слова в тексте и самостоятельно их записывать.
Воспроизвести знания об одинаковом написании корня в однокоренных словах.
Развивать речь учащихся, мышление, внимание.
Воспитывать интерес к урокам русского языка.
Задачи:
*содействовать формированию умения разбирать слова по составу;
*развивать умение выбирать из предложенного списка слова, соответствующие заданной схеме;
*развивать умение работать с текстом: определять тему и главную мысль, подбирать заголовок и части текста.
Ход урока
- Организационный момент. Запись числа в тетради.
– Урок наш будем измерять шагами. Каждый шаг – это проделанная работа.
2 . Постановка темы, цели урока.
1 шаг.
– Посмотрите на доску, что за значки вы видите? На доске нарисовано
– Расшифруйте значки: окончание, основа, корень, приставка, суффикс.
– Назовем эти понятия одним словом – морфемы.
Морфема – это наименьшая значимая часть слова
Отгадаем загадку:
Слово делится на части
Ах, какое это счастье!
Может каждый грамотей
Собрать слово из частей.
– О каких частях слова говорится?
– Какие части слова вы запомнили с уроков украинского языка?
– Сегодня мы вспомним конкретнее о составе слова и его составляющих.
3. Работа над материалом урока.
2 шаг.
Если знаешь части слова,
То напишешь их толково.
Помни при разборе слова
Окончанье и основу
Первым делом находи.
Окончание – значимая часть слова, которая образует формы слова. Окончание может указать на часть речи.
Окончание слова определяется с помощью изменения формы слова.
Задание самостоятельно : выделить окончания в словах.
Зима, зимний, снежинка, заморозит.
3 шаг
Основа – это часть слова без окончания. В основе слова может быть одна морфема (корень), но может быть и больше.
Задание в паре : выделить основы слов.
Перемена, пришкольный, звонок, учительница.
– Какие морфемы входят в состав основы?
4 шаг
Корень слова – значимая часть.
Над словами родственными держит власть.
Выясни умело линию родства –
Однокоренные подбери слова.
Корень – главная значимая часть слова, в которой заключено общее лексическое значение всех однокоренных слов. Для того чтобы выделить корень надо подобрать однокоренное слово.
Задание в группе по 4 человека
5 шаг
– После корня будет суффикс.
Суффикс – суффикс значимая часть слова, которая стоит после корня и служит для образования новых слов.![]() Чтобы правильно выделить суффикс надо подобрать ряд родственных слов.
Чтобы правильно выделить суффикс надо подобрать ряд родственных слов.
Задание в паре : в словах выделить суффиксы.
Гармонист, летчик, ежик, домишко.
6 шаг
Приставка ( префикс ) – значимая часть слова, которая стоит перед корнем и служит для образования новых слов. Чтобы выделить приставку, нужно подобрать ряд однокоренных слов и с помощью сравнения выделить данную часть слова.
Задание коллективно : выделить приставки в словах
Убежал, записать, распрекрасный, подберезовик.
Индивидуальная дифференцированная работа (работа по рядам).
Вставьте нужную приставку и запишите полученное слово.
4. Закрепление материала урока
Послушайте историю.
Собрались у Воды родственники. Подводник с Водицей беседуют. Водолаз с Водопадом на солнышке греются. Водитель на гармошке играет. Водомерка с Водорослями заигрались. Даже сам Водяной пожаловал. И все старуху Воду ждут. Вышла мудрая Вода на крылечко, глянула на гостей, сразу чужака приметила. Велела ему идти в свою семью.
Учитель: Кто оказался чужаком? ( Водитель).
Учитель: Назовите однокоренные слова к слову водитель. ……….водить,водила,заводить.
7 шаг
Творческое задание для групп. Учитель читает задания ,дети записывают на листочках, потом вслух говорят ответ.
Учитель читает задания ,дети записывают на листочках, потом вслух говорят ответ.
А теперь вы попробуете из указанных мною морфем собрать слова.
А) Возьмите тот же корень, что в слове «клонится».
Добавьте приставку, которая есть в слове «слезет».
Суффикс и окончание возьмите из слова «обращение». (склонение)
Б) Возьмите корень из слова «полагать», приставку- из слова «приложение», суффиксы- из слова восклицательный, а окончание- из слова «числительное». Подсказка: получится слово- часть речи, обозначающая признак предмета (прилагательное)
Подсказка: получится слово- часть речи, обозначающая признак предмета (прилагательное)
В) Возьмите приставку из слова «подруга», корень- из слова «игрушка», суффиксы- из слова «читал», а окончание в этом слове будет такое же, как в слове «стул» (поиграл)
Г) Это будет слово, однокоренное со словом «летчик». Добавьте к корню такую же приставку, как в слове «перепрыгнул». Оба суффикса этого слова такие, как в глаголе «слышать» (перелетать)
8 шаг
Закрепление материала
Ст. 108 упр. 262
108 упр. 262
Зимушка, зима, зимний
9 шаг
Самостоятельная работа
10 шаг
Итог урока:
Учитель: Ребята, все вы сегодня работали очень хорошо.
– Ребята, наш урок подходит к концу.
-Что вы сегодня нового узнали на уроке?
-Какие задания были самые интересные?
-В чём испытали затруднения?
-Сейчас, я раздам вам карточки-светофоры. Раскрасьте зелёный круг, если вам все понравилось, жёлтый, если понравилось, но что-то было непонятно. И красный, если не понятно, не понравилось.
– Спасибо за старание, все сегодня молодцы!
Конспект урока Однокоренные слова.
 Корень слова.
Корень слова.Тема: Однокоренные слова. Корень слова.
Цель: развитие умений анализировать однокоренные слова, выделять корень, определять лексическое значение корня и общее значение однокоренных слов.
Задачи:
Развить умение находить однокоренные слова;
Развивать умение выделять в однокоренных словах корень;
Понимать лексическое значение однокоренных слов и корня в них.
Планируемые результаты урока:
Сформируется базовый уровень умений:
– Анализировать однокоренные слова: называть общее лексическое значение корня.
– Выделять корень в слове, в группе однокоренных слов, подбирать однокоренные слова.
– Отличать однокоренные слова от слов с похожим корнем (горе, гора, гореть).
Универсальные учебные действия (УУД)
Познавательные:
Развивать умение извлекать информацию из схем, иллюстраций, текстов.
Выявлять сущность, особенности объектов.
На основе анализа объектов делать выводы.
Обобщать и классифицировать по признакам.
Ориентироваться на страницах учебника.
Регулятивные:
Самостоятельно организовывать свое рабочее место.
Развиваем умение высказывать своё предположение на основе работы с материалом учебника.
Оценивать учебные действия в соответствии с поставленной задачей.
Прогнозировать предстоящую работу (составлять план).
Осуществлять познавательную и личностную рефлексию.
Коммуникативные:
Развиваем умение слушать и понимать других.
Строить речевое высказывание в соответствии с поставленными задачами.
Оформлять свои мысли в устной форме.
Умение работать в паре.
Личностные:
Выполнять правила этикета.
Развиваем умения высказывать своё отношение, выражать свои эмоции.
Оценивать поступки в соответствии с определённой ситуацией.
Формируем мотивацию к обучению и целенаправленной познавательной деятельности.
1. Орг. момент.
– Здравствуйте, ребята! Повернитесь друг к другу, улыбнитесь и пожелайте удачи себе и своим соседям! Откройте тетради, запишите число, классная работа.
2.Чистописание.
-На минутке чистописания повторим соединения буквы ~ «дэ». (д)
Д Дд дд ди до од род
5.Словарная работа
(1 ученик на доске, остальные в тетрадях)
Доставляющий наслаждение взору, приятный внешним видом
Хозяйственные предметы, предназначенные для хранения продуктов, а также еды, питья.
Произведение живописи, выполненное в красках на бумаге, доске, ткани.
Специалист, который преподает какой-либо учебный предмет в школе
Тонкая палочка графита, вделанная в деревянную оболочку.
Последний осенний месяц.
II. Работа над темой урока.
-Прочитайте три группы слов, написанных на доске.
1 2 3
Сор, соринка, сорный
Снег, снега, снегом
Солдат, боец, воин
— Найдите группу родственных слов. (1-я)
— Почему вы так считаете? Докажите. (общая часть, которая общая по смыслу и написанию)
— Как называется эта общая часть родственных слов? (Корень)
— Это разные слова или одно и то же слово? (Разные однокоренные слова)
— Что вы можете сказать о второй группе слов? (Формы одного и того же слова).
— Почему вы так считаете? Докажите. (Одинаковые основы)
— Что вы можете сказать о третьей группе слов? Это родственные слова? (Нет. Только близкие по значению)
2.Определение темы урока :
— Как вы думаете, о чем пойдет речь на нашем уроке?
(Об однокоренных словах, о корне слова, их общей части)
Какая тема нашего урока?
— Правильно, ребята. На уроке мы будем продолжать работать над темой «Корень слова. Однокоренные слова».
3.Работа со словарём.
— Как вы думаете, все ли вы знаете о корне и однокоренных словах?
Слово корень – многозначное. Обратимся к словарю и посмотрим значения этого слова.
Чтение сообщений по карточкам.
-Итак, корень – это подземная часть растения, внутренняя находящаяся в теле часть зуба.
Что общего в значениях слова корень? (Важная часть чего-либо)
А ещё в словаре говорится, что корень – это основная значимая часть слова.
Как по-другому можно сказать о корне слова?
Вывод: корень – главная часть слова.
4. Физкультминутка
VI. Закрепление изученного материала.
1. Самостоятельная работа. Работа с учебником.
а) Упр. 85 с.61.
– Как правильно определить в слове корень? (Подобрать однокоренные слова и найти общую часть)
VIII. Проверка изученного
Задание «Верно ли, что …»
а) каждый ученик имеет карточку красного и зелёного цветов, которые использует при выполнении задания;
б) учитель читает фразы;
Верно ли, что в словах русского языка может быть два корня?
Верно ли, что слово зерносушилка образовалось от слов картофель сушить?
Верно ли, что в слове цветник два корня?
Верно ли, что от фразы кофе варить образовалось слово кофемолка?
Верно ли, что в слове жизнерадостный два корня?
в) после каждой фразы школьники осмысливают сказанное, и, если с высказыванием согласны, поднимают карточку зелёного цвета, если не согласны – красного.
VII. Итог урока.
– Что такое корень?
– Роль корня в слове.
– Как выделить корень в слове?
– Какие слова называются однокоренными?
VIII. Рефлексия.
-Я узнал…
Я научился…
Мне было легко…
Я доволен собой,…
Адрес публикации: https://www.prodlenka.org/metodicheskie-razrabotki/362047-konspekt-uroka-odnokorennye-slova-koren-slova
Урок русского языка во 2 классе на тему: “Корень слова. Однокоренные слова” с использованием электронных образовательных ресурсов
Библиографическое описание:Карапузова, Н. Н. Урок русского языка во 2 классе на тему: “Корень слова. Однокоренные слова” с использованием электронных образовательных ресурсов / Н. Н. Карапузова. — Текст : непосредственный // Педагогическое мастерство : материалы VII Междунар. науч. конф. (г. Москва, ноябрь 2015 г.). — Москва : Буки-Веди, 2015. — С. 133-136. — URL: https://moluch.ru/conf/ped/archive/184/9113/ (дата обращения: 01.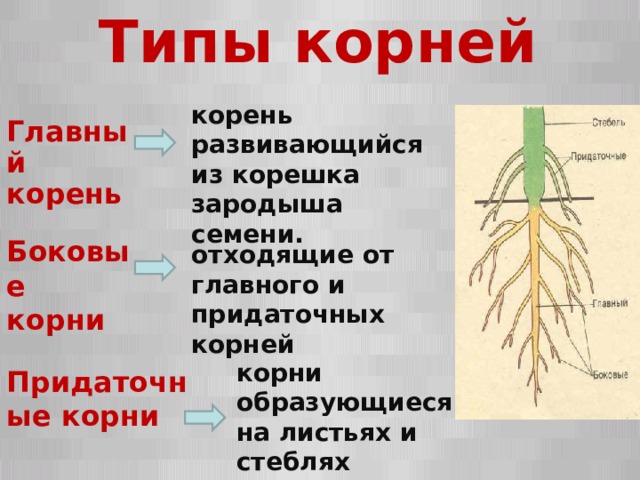 10.2022).
10.2022).
Тема: Корень слова. Однокоренные слова
Цель: познакомить с понятиями корень, однокоренные слова; развивать речь; пополнять словарный запас учащихся
Планируемые результаты:
—предметные: научить находить в словах корень и образовывать однокоренные слова, употреблять их в речи;
— метапредметные: аргументированно отвечать, доказывать свое мнение,анализировать, делать выводы, сравнивать;
— личностные: воспитывать интерес и любовь к родному языку, аккуратность, целеустремлённость.
Тип урока: овладения новыми знаниями
Формы работы учащихся: индивидуальная, фронтальная, работа в парах
Оборудование: интерактивная доска, мультимедийная презентация «Корень слова. Однокоренные слова», толковый словарь, сигнальные карточки, ЭОР.
Однокоренные слова», толковый словарь, сигнальные карточки, ЭОР.
Технологическая карта урока
|
Этап урока |
Формы, методы, методические приёмы |
Название используемых ЭОР (с указанием порядкого номера из таблицы) |
Содержание учебного материала Деятельность учителя с указанием действий ЭОР |
Деятельность учащихся |
Формирование УУД |
|
I этап. Организационный момент. Цель: мотивирование учащихся к учебной деятельности посредством создания эмоциональной обстановки |
Фронтальная. Словесный. Слово учителя |
|
Начинается урок, Он пойдёт ребятам впрок, Постарайтесь всё понять, Учитесь тайны открывать, Ответы полные давать, Чтоб за работу получать Только лишь отметку «пять»! (слай1) — Предлагаю, чтобы сегодняшний наш урок прошёл под девизом: (слайд2) «Ум да разум надоумят сразу» |
Проверяют готов-ность к уроку |
Регулятивные УУД:волевая саморегуляция. Личностные УУД: действие смыслообразова-ния. Коммуникатив-ные УУД: планирование учебного сотруд-ничества с учите-лем и со сверст-никами |
|
II этап. Минутка чистописания Цель: отработка каллиграфического написания определённых букв
Словарная работа Цель: повторить правописание непроверяемых гласных и согласных в словах |
Наглядный, практический, частично-поисковый
Практический, взаимопроверка |
Правописание заглавных и строчных букв ЭОР№ 1
http://files. |
Демонстрирует учащимся буквы КРНЬ. Уточняет, что данные буквы в пути потеряли гласные буквы. —Какие гласные убежали из слова? —Определите лексическое значение слову «корень»
—Спишите слова в тетради, вставляя пропущенные буквы. К..п..тан, с..одня ..ктябрь, лес.н…ца р…бята, ок…л… с…рень, вмест… а……уратный |
Прописывают каллиграфически в тетради буквы по образцу.
Записывают словарные слова в тетрадь, встав-ляя пропущен-ные буквы |
Познавательные УУД: общеучебные: умение структу-рировать знания, контроль и оценка процесса и результатов деятельности; логические: анализ, синтез, выбор оснований для сравнения. Регулятивные УДД: контроль, кор-рекция; прогнозирование (при анализе пробного действия перед его выполнением)
|
|
III этап. Цель: проверка уровня усвоения полученных знаний по теме «Родственные слова» |
Практический, наглядный |
Игра «Рассели жильцов» Интерактивное задание ЭОР№ 2
http://files.school-collection.edu.ru/dlrstore/2eea00cc-de75–4d72–9dcc-ca04b7853691/ %5BNS-RUS_2–09 %5D_ %5BID_004 %5D.swf |
Предлагает учащимся задание на развитие орфографической зоркости и умение видеть родственные слова |
Выполняют практические действия на нахождение и объяснение в словах изученных орфограмм. Анализируют записанные слова, доказывают: все ли они являются родственными словами, выделяют общую часть |
Познавательные УУД: общеучебные: умение структурировать знания, контроль и оценка процесса и результатов деятельности;
логические: анализ, синтез, выбор оснований для сравнения. Регулятивные УУД: контроль, коррекция; прогнозирование (при анализе пробного действия перед его выполнением) |
|
IV этап. Самоопределение к деятельности. Цель: постановка учебной проблемы, выделение и формулирование познавательной цели |
Словесный, фронтальный, проблемный |
|
Формулирует проблемный вопрос «Как иначе можно назвать общую часть родственных слов?» —Определите тему нашего урока. — Какую цель поста-вим на уроке? |
Высказывают предположения. Формулируют под руководством учителя цели урока. |
Познавательные УУД: умение структу-рировать знания; постановка и формулирование проблемы; умение осознанно и произвольно строить речевое высказывание |
|
V этап. Первичное усвоение новых знаний. Цель: познакомить с понятиями ко-рень, однокорен-ные слова; научить находить корень в словах. 1.Введение понятия «корень слова».
2. |
Наглядный, словесный |
Что такое корень? ЭОР№ 3 http://files.school-collection.edu.ru/dlrstore/4cdf6c5c-633a-4c3f-abcb-28a320286261/ %5BNS-RUS_2–09 %5D_ %5BTE_002 %5D.swf Как найти корень в слове? ЭОР№ 4
http://files.school-collection.edu.ru/dlrstore/56ed9f51–5629–480a-93b7–910f78a0fab0/ %5BNS-RUS_2–09 %5D_ %5BQN_003 %5D.swf |
Предлагает учащимся работу с интерактивным модулем. Проверку организует фронтально. |
Словарик. Выясняют, что такое «корень». Выполняют практическую работу (на интерактивной доске) |
Познавательные УУД: общеучебные: поиск и выделение необходимой ин-формации, приме-нение методов информационного поиска; смысловое чтение и выбор чтения в зависимости от цели; умение осознанно и произвольно строить речевое высказывание;
логические: построение логической цепи рассуждений, анализ, синтез. |
|
Физминутка Цель: профилак-тика утомления |
Коллективный, практический |
|
Утром на лесной опушке Собираются зверушки. Все приучены к порядку: Дружно делают зарядку |
Выполняют движения под руководством учителя |
Регулятивные УУД: осуществляют профилактику утомления |
|
VI этап. Цель: закрепить новые полученные знания |
Практический. Работа в парах. Взаимопровер- ка. Взаимооценка. |
|
Предлагает учащимся задания учебника на поиск однокоренных слов и выделение корня в найденных однокоренных словах. |
Выполняют задания в тетради (упр.№ 87 с. 62) |
Коммуникатив-ные УУД: управление пове-дением партнера; умение выражать свои мысли |
|
VII этап. Цель: самооценка деятельности |
Фронтальная, индивидуаль-ная. Словесный. Беседа |
|
— Что нового вы узнали на уроке? — Что особенно вам понравилось? Почему? — Что вызвало затрудне-ние? Почему? — Кто может сказать, что уже понял, что такое корень слова? —Какие знания, умения, навыки помогали нам сегодня на уроке? — Спасибо за урок |
Высказывают мнение. Форму-лируют, что ещё по данной теме они хотели бы узнать и чему научиться. |
Регулятивные УУД: оценка– выделе-ние и осознание того, что уже усвоено и что еще подлежит усвое-нию
|
|
VIII этап. Дом. задание |
Фронтальный, словесный. Слово учителя |
|
Выполнить упр.№ 88(с.62) дописать в каждую группу по два однокоренных слова |
|
Регулятивные УУД:прогно-зирование. |

 school-collection.edu.ru/dlrstore/6979e3af-097b-445c-92b4-a1ca1a73d81d/ResFile.SWF
school-collection.edu.ru/dlrstore/6979e3af-097b-445c-92b4-a1ca1a73d81d/ResFile.SWF
 Дополняют буквы гласными, получают и записывают в тетрадь слово «корень»
Дополняют буквы гласными, получают и записывают в тетрадь слово «корень»
 Актуализация опорных знаний.
Актуализация опорных знаний.

 Как найти корень слова?
Как найти корень слова?

 Первичное закрепление изученного (работа в тетради по заданиям учебника)
Первичное закрепление изученного (работа в тетради по заданиям учебника)
 Рефлексия.
Рефлексия.

Основные термины (генерируются автоматически): слово, корень слова, знание, умение, урок, этап, интерактивная доска, общая часть, пробное действие, речевое высказывание.
Интерактивный метод обучения на уроках русского языка как…Корень — главная значимая часть слова, в которой заключается общее значение всех однокоренных слов.
интерактивное обучение, учебный процесс, образовательный процесс, русский язык, урок, групповая работа, познавательная деятельность, умение, знание, учащийся.
Использование
интерактивных методов обучения на уроках…Продуктивный урок должен формировать не только глубокие и прочные знания, но и умения
Предлагаю рассмотреть структура интерактивного урока
Этот этап начинается обычно с мозгового штурма, например: «Какие понятия связаны со словом лексика?»
Современный
урок русского языка. Технологическая карта урока…Тема урока: «Корень слова». Тип урока: Урок формирования и первичного закрепления новых знаний.
План урока. Составление «кластера» о корне слова (главная часть слова, лексическое значение). Ответ на тонкие и толстые вопросы.
Формирование познавательных универсальных учебных
действий…часть речи, русский язык, какая часть речи, урок, слово, местоимение, группа, пробное действие, самостоятельная работа, место имени существительного. Дифференциация на уроках окружающего мира в начальной…
Внеклассное мероприятие «Открываем секреты словарей»
‒ формировать умение использовать алфавитный принцип при нахождении нужного слова в
Активно включает работу со словарём на всех предметах и на каждом этапе урока.
Оборудование: ‒ 5 словарей толковых, фразеологических словарей. ‒ Интерактивная доска.
Игровые
интерактивные технологии обучения на уроке русского…Ключевые слова: игровые интерактивные технологии, русский язык как иностранный, методика обучения РКИ
Преподаватель подбирает текст, в который могут быть вставлены слова одной тематической группы, однокоренные, либо синонимы, различных частей речи.
Развитие
речи учащихся на уроках в начальной школеСовершенствование речевого общения. Использование возможностей урока и внеурочной работы для развития речевой деятельности детей.
Перед корнем, раз и рас, из и ис, слова часть, только вместе, а приставки не бывает, в и вз, к предлог, а не приставка, а как же без неё.
Использование
интерактивного метода обучения на уроках…В этом плане ориентация на интерактивные методы обучения способствуют овладению знаниями, формированию
Такая работа возможна на уроке любого типа, на разных его этапах.
Ашукин Н. С., Ашукина М. Г. Крылатые слова. М.: Художественная литература, 1987.
Интерактивные методы обучения на уроках русского языка…Скачать Часть 11 (pdf). Библиографическое описание: Казыгулова А. Т. Интерактивные методы обучения на уроках русского языка и литературы
В энциклопедическом словаре даётся такое толкование к слову технология — (от греч. techne — искусство, мастерство, умение и …логия). ..
..
“Корень слова и его варианты.Однокоренные слова” урок 5 класс
Тема: Корень слова и его варианты. Однокоренные слова.
Цели урока:
Познакомить учащихся с понятиями «корень и его варианты», «однокоренные слова»; учить
учащихся находить корень в словах, распознать и подбирать однокоренные
слова, обогащать словарный запас учащихся.
Развивать наблюдательность, память, совершенствовать навыки
аргументированного и лаконичного изложения мыслей и умения вести
учебный диалог, умение делать выводы, развивать творческие способности, речь.
Воспитывать любовь к родному языку, потребность пользоваться его
языковым богатством, умение работать совместно, привить патриотические чувства у учащихся, любовь к Родине, к окружающей среде, чувства бережного отношения к природе.
Тип урока: объяснение нового материала, используя элементы сказки.
Метод: словесный, кластерность, метод направленного чтения.
Межпредметная связь: чтение, биология.
Наглядность: учебник, интерактивная доска, опорная схема, буклеты.
Ход урока:
І. Организационның момент. Слово учителя.
1. Эмоциональный настрой.
Учитель просит детей встать в круг и по очереди пожелать друг другу хорошее, например; добра, счастья, удачного дня и т.д.
2.Деление класса на 2 группы.
Раздаются стикеры с разными рисунками, равными по количеству.
ІІ. Проверка домашнего задания.
Фронтальный опрос: учащиеся задают вопросы друг другу
/предполагаемые вопросы учащихся/
– Какую тему изучали на прошлом уроке? ( Состав слова и окончание)
– Что входит в основу слова? ( корень, приставка,суффикс)
– Что такое основа слова? ( Часть слова без окончания)
-Как переводятся на казахский язык; корень суффикс, окончание, основа слова и как они условно обозначаются? Показать на примере слова -загадка
– Как определяем твёрдую и мягкую основу? ( после тв. основы
основы
пишутся буквы а,о,у,ы; после мягкой ь,и,е,ё,ю,я)
(таблица уч. стр. 63)-слайд 5
2.Определите в словах основу, выделите основу и окончание Слайд 3
Слово | Мягкая основа | Твердая основа |
поле | ||
река | ||
дом | ||
речь | ||
родная | ||
синяя | ||
белая | ||
смелая |
Слайд 4- ответы
ІІІ.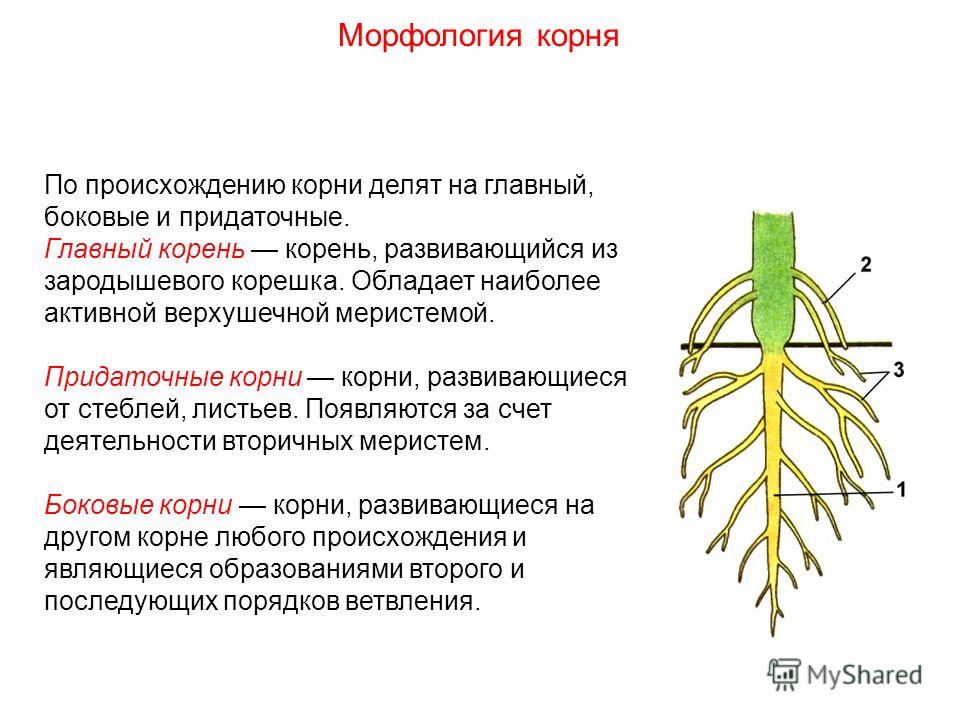 Объяснение нового материала
Объяснение нового материала
Ребята , я загадаю вам загадку. Загадывание загадки Слайд 6
Я у дуба.
Я у зуба.
Я у слов и у цветов.
Я упрятан в темноту.
Я не в верх, а вниз расту
Я главная часть слова
Я значение слова создаю (корень)
Ответ- слайд 7
И, действительно, ребята Корень главная значимая часть слова.
Сегодняшний урок мы посвятим этой теме.
2. Запись темы и задачи урока.
-Запишите тему урока в тетрадь. Корень слова и его варианты, однокоренные слова.
Наша задача сегодня: познакомиться с понятиями «корень слова», слайд 2
«однокоренные слова». Научиться определять в словах
корень, находить однокоренные слова. Запишите тему
урока в тетрадь.
Запись на доске : Зри в корень! Слайд 8
– Как вы понимаете это выражение? ( Это значит, что надо смотреть на
самое важное,синоним этого слова смотри)
Правильно.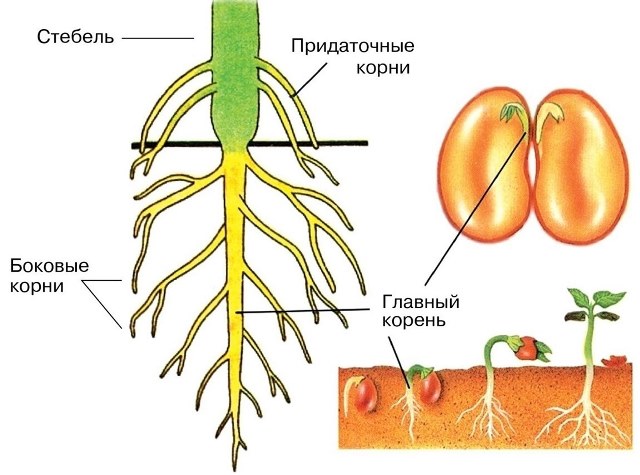 Потому что в корне заключён главный смысл всех родственных слов.
Потому что в корне заключён главный смысл всех родственных слов.
3. Определение значения слова корень.
Корень –это Слайд 9
1. Подземная часть растения.
2. Начало, источник чего-нибудь.
3. В грамматике – часть основы слова, в которой заключено его значение.(буклет с рисунками; зуб с корнем, цветок с корнем, разобранное по составу слово, а также словесный пример родословной одного из учащихся- жеты ата)
4. Сказка «Чудесное дерево» Упр.142
– Словарно – лексическая работа (рисунок дерева) Слайд 10
– выразительное чтение учителем сказки.
– назовите в тексте слова, которые имеют один и тот же корень.
– (лететь,прилететь, улететь, вылететь, полететь, долететь, перелететь, перелётная). Корень этих слов – лет-, от глагола летать.
– подумайте, как в русском языке называются слова, образованные от одного корня?
Вывод: Слова, образованные от одного корня называются Слайд 11
однокоренными.
-А что такое корень слова?
Вывод: Корень слова – общая часть родственных слов. Слайд 12
Однокоренные слова надо отличать от разных форм одного и того
же слова.
Входить, вход, входной – это однокоренные слова.
Вход, у входа – это разные формы одного и того же слова.
Окончание образует формы слова.
Понятие о варианте корня. Ребята, посмотрите на эти пары слов и скажите; однокоренные ли они и какой процесс вы в них прослеживаете:
Книга-книжка
Писать-пишу
Ответить-отвечу
(Ответ: процесс чередования согласных звуков г/ж, с/ш, т/ч в корне слова)
Ребята, поэтому в результате чередования звуков в корне слова образуются варианты корня
ІҮ. Закрепление
Выполните упражнение № 145 стр. 64.
1) В каждой строчке найдите однокоренные слова.
Лист, листик, листва, цветок
Золото, золотой, золотистый, серебрянный
Дождь, дождливый, дождевой, снег
Жёлтый, желтеть, пожелтеть, зеленый
Вода, водный, водяной, водитель
Ответы- слайд 13
-Почему слова каждой группы ты считаешь однокоренными?
(Потому что у них общий корень)
2)Выборочный диктант. (выполненная работа взаимопроверяется группами) Слайд 14
(выполненная работа взаимопроверяется группами) Слайд 14
Выпишите однокоренные слова. 1 группа- к слову лететь, земля, цвет. 2 группа к словам цветы, лето, хлеб.
Лететь, прилететь, улететь, цвет, цветной, цветы, долететь, летний, лето цветочный, земли, зимовать, земной, подземный, хлебушко,хлебный, хлеб.
Ответы- слайд 15
Ү. Самостоятельная индивидуальная работа учащихся.
(выполненная работа взаимопроверяется соседним учащимся)
Слайд 16
1-ое задание: Определите, где однокоренные слова, а где формы слова? Найдите слово где произошло чередование согласного в корне?
? | ? | |
сад | садовник | сады |
садовый | в саду | |
сажать | о саде | |
лес | лесник | леса |
лесной | по лесу | |
лесистый | лесом | |
Ответы- слайд 17
2-ое задание: Образуйте группу слов, связанных со словами школа, дом.
Слайд 18, ответы-слайд 19
школа, дом, домовой, школьник, домашний, школьница, пришкольный,домик
ҮІ. Игра «Хлопаем в ладоши» Слайд 20
Сейчас я буду называть пары слов. В одних парах однокоренные, в других- не однокоренные. Если я назову однокоренные слова вы хлопаете в ладоши, если я назову не однокоренные, то вы поднимаете руки.
Лес – лесник
Вода – водопад
Сад- садовник
Дом – дым
Поле – море
Родина- родной
Природа- край
Обед- стол
Зима- Зимний
Друг- дружба
ҮІІ. Работа с текстом. Проблемная ситуация. Выборочное списывание. Слайд 21
Чужак
Собрались у Воды родственники. Подводник с Водицей беседуют. Водолаз с Водопадом на солнышке греются. Водитель на гормошке наигрывает. Водомерка с Водорослями разыгралась. Водичка по камушкам на одной ножке скачет. Даже сам Водяной пожаловал. Все старуху Воду ждут.
Водичка по камушкам на одной ножке скачет. Даже сам Водяной пожаловал. Все старуху Воду ждут.
Вышла мудрая Вода на крыльцо, глянула на гостей, сразу чужака приметила. Велела ему прочь идти, в свою семью. Пошел чужак, пригорюнился. Где ему родственников искать?
Задание: Выпишите в тетрадь однокоренные слова. Выделите в них корень.
– Какие однокоренные слова вы выписали? Обоснуйте свой ответ.
( Вода, подводник, водицей, водолаз, водопад, водомерка, водорослями, водичка,водяной).
– Найдите формы одного и того же слова (вода, воды, воду– изменены окончания).
Проблемный вопрос:
– А какое слово оказалось чужаком среди однокоренных к слову Вода?
(Водитель)
-Обоснуйте свой ответ.
Ответ- слайд 22
Вывод:
Признаки однокоренных
слов
имеют общую Близки по смыслу
часть
Итак, ребята корень этих слов вод от слова вода. А вода часть природы. А что такое природа? Это то, что нас окружает. Воздух, небо, леса и горы, птицы и животные. И природу мы должны беречь и охранять.
А вода часть природы. А что такое природа? Это то, что нас окружает. Воздух, небо, леса и горы, птицы и животные. И природу мы должны беречь и охранять.
VIII. Подведение итогов урока. Рефлексно-оценочный этап. Слайд 23. Ответы – слайд 24
Тест /да-нет/:
Вопрос | ответ | да | нет |
Что такое корень слова? | Корень слова – общая часть родственных слов | ||
Какие слова называются однокоренными? | Слова, образованные от одного корня называются однокоренными | ||
Однокоренные слова имеют общую часть? | |||
Однокоренные слова не близки по смыслу | |||
Как выделить корень слова? | Нужно подобрать однокоренные слова |
IХ. Задание на дом. Слайд 25
Задание на дом. Слайд 25
Упражнение 150 стр. 65
Правило стр. 63-64.
Написать мини-сочинение на тему:«Охранять природу- значить охранять Родину »
4. Дополнительное задание для тех, кто хочет знать больше. Карточки: правило и слова для разбора по составу
Х. Мнение учащихся об оценках за урок. Комментирование оценок.
XI. Отзыв учащихся об уроке в письменном виде на стикерах и прикрепление стикеров на рисунки с изображением лиц с положительной и отрицательной мимикой.
Корневая разработка — PMC
- Список журналов
- Книга арабидопсиса
- т.1; 2002 г.
- PMC3243376
Книга арабидопсиса. 2002 г.; 1: e0101.
2002 г.; 1: e0101.
Опубликовано в Интернете 30 сентября 2002 г. doi: 10.1199/tab.0101
, , и
Информация об авторе Информация об авторских правах и лицензии Отказ от ответственности
С точки зрения развития, корень арабидопсиса является образцом простоты. Небольшое количество стволовых клеток на кончике корня генерирует все типы клеток посредством стереотипных делений с последующей клеточной дифференцировкой и регулируемой клеточной экспансией. Поскольку рост корня не определен, эти процессы непрерывны, в результате чего все стадии развития присутствуют постоянно. Радиальная симметрия корня в сочетании с отсутствием движения клеток означает, что клонально родственные клетки часто обнаруживаются в клеточных файлах. Эти клеточные файлы можно проследить до их происхождения, которое представляет собой четыре типа стволовых клеток (или исходных клеток) на кончике корня (Dolan et al., 19).93). Эпидермальные/боковые зачатки корневого чехлика дают начало эпидермису и внешней части корневого чехлика, известной как латеральный корневой чехлик (4). Центральная часть корневого чехлика, колумелла, имеет собственный набор инициалов. Клетки основной ткани, кора и энтодерма, образуются путем деления коры/энтодермальных инициалей. Наконец, сосудистая ткань и перицикл имеют свои инициалы. Внутри и в контакте со всеми инициалями находится небольшое количество центральных клеток, которые митотически неактивны и известны как центр покоя (QC) ().
Центральная часть корневого чехлика, колумелла, имеет собственный набор инициалов. Клетки основной ткани, кора и энтодерма, образуются путем деления коры/энтодермальных инициалей. Наконец, сосудистая ткань и перицикл имеют свои инициалы. Внутри и в контакте со всеми инициалями находится небольшое количество центральных клеток, которые митотически неактивны и известны как центр покоя (QC) ().
Открыть в отдельном окне
а) Продольный срез кончика корня арабидопсиса с цветовой кодировкой для демонстрации различных типов клеток. b) Схема, показывающая два асимметричных клеточных деления, которые претерпевают начальная и дочерняя кора/энтодерма.
Деление инициалей может быть либо исключительно антиклинальным (ортогональным к оси роста) с образованием одного ряда клеток, либо сначала антиклинальным, а затем периклинальным (параллельным оси роста) с образованием двух или более слоев клеток. Инициалы колумеллы обычно делятся только антиклинально, и их потомство подвергается быстрому расширению клеток, а затем дифференцируется, продуцируя содержащие крахмал амилопласты, которые играют роль в восприятии гравитации. Три других типа инициалей обычно претерпевают как антиклинальные, так и периклинальные деления, в результате чего клеточные линии приобретают разные идентичности (Dolan et al., 19).93). Например, инициали коры/энтодермы сначала делятся антиклинально, чтобы регенерировать начальную клетку и базальную дочернюю клетку. Затем базальная дочерняя клетка подвергается периклинальному делению с образованием первых клеток коры и энтодермальных клонов. Эти клетки претерпят небольшое количество антиклинальных делений, в течение которых они приобретут свои специфические судьбы. Впоследствии они будут подвергаться быстрому полярному расширению.
Три других типа инициалей обычно претерпевают как антиклинальные, так и периклинальные деления, в результате чего клеточные линии приобретают разные идентичности (Dolan et al., 19).93). Например, инициали коры/энтодермы сначала делятся антиклинально, чтобы регенерировать начальную клетку и базальную дочернюю клетку. Затем базальная дочерняя клетка подвергается периклинальному делению с образованием первых клеток коры и энтодермальных клонов. Эти клетки претерпят небольшое количество антиклинальных делений, в течение которых они приобретут свои специфические судьбы. Впоследствии они будут подвергаться быстрому полярному расширению.
Во время развития эмбриона арабидопсиса клеточное деление происходит по стереотипному образцу. Это облегчает идентификацию клеток-основателей первичного корня. Происхождение покоящегося центра и корневого чехлика колумеллы можно проследить до одной клетки, гипофиза (Scheres et al. 19).94). Эта клетка, в свою очередь, происходит от базальной дочерней клетки первого зиготического деления (1) и является единственным вкладом базальной клетки в собственно зародыш.
Открыть в отдельном окне
Эмбриональное происхождение корня Arabidopsis . Слева направо: первое деление зиготы; зародыш стадии октанта; эмбрион на стадии сердца; проросток с увеличенным участком корневой меристемы.
Остальные клетки, которые образуют корень в зрелом эмбрионе, происходят из апикальной дочерней клетки зиготы. Их радиальная организация в слоях протодермы, основной ткани и прокамбиального слоя очень похожа на организацию всей оси зародыша, за исключением основной ткани (см. ниже). Действительно, мутации в нескольких генах, которые влияют на радиальную организацию корня, обнаруживают идентичные фенотипы корня и гипокотиля (Scheres et al. 19).95).
Граница между корнем и гипокотилем не очевидна из анатомии зародыша, хотя она четко обозначена постэмбрионально по характеристикам дифференцировки отдельных типов клеток, таким как образование корневых волосков и по содержанию хлорофилла в клетках основной ткани. Гипокотиль содержит три слоя основной ткани, тогда как большая часть корня содержит два. У зародыша видно исчезновение одного слоя основной ткани, но оно не совпадает с границей корня и гипокотиля, определяемой ориентирами дифференцировки клеток.
У зародыша видно исчезновение одного слоя основной ткани, но оно не совпадает с границей корня и гипокотиля, определяемой ориентирами дифференцировки клеток.
Клональный анализ для изучения развития зародышевых корней
Клональный анализ — это метод, который можно использовать для отслеживания судеб потомков отдельных клеток в многоклеточном организме. Он использовался для определения судеб клеток в зародыше, дающих начало первичному корню, и судеб клеток в корневой меристеме проростка. Клональный анализ зависит от возможности пометить клетку видимым генетическим маркером, который впоследствии можно будет подсчитать. Генетический маркер наследуется всеми потомками исходной меченой клетки, и эта группа меченых клеток составляет клон. Размеры клона отражают вклад клетки-предшественника в зрелый организм. Такие параметры, как размер и распределение клонов, позволяют определить такие параметры развития, как скорость деления клеток, количество клеток-основателей и т. д. Генетические маркеры, которые широко использовались в клональном анализе побегов, включают мутации, придающие фенотипы дефицита хлорофилла (альбиносные мутанты), или мутации антоцианов, которые придают либо интенсивный цвет пигмента, либо потерю цвета пигмента. В корнях таких маркеров немного. Вместо этого клоны были отмечены цифрой 9.0036 uid Ген (ген глюкуронидазы), который катализирует образование голубого осадка в клетках, экспрессирующих ген, при инкубации с соответствующим субстратом. В описанных исследованиях были сконструированы растения, трансформированные геном uid A , в который был вставлен транспозон Ac или Ds. Удаление транспозона из гена uidA приводит к восстановлению активности uidA , которая маркирует клетки.
В корнях таких маркеров немного. Вместо этого клоны были отмечены цифрой 9.0036 uid Ген (ген глюкуронидазы), который катализирует образование голубого осадка в клетках, экспрессирующих ген, при инкубации с соответствующим субстратом. В описанных исследованиях были сконструированы растения, трансформированные геном uid A , в который был вставлен транспозон Ac или Ds. Удаление транспозона из гена uidA приводит к восстановлению активности uidA , которая маркирует клетки.
Положение границы корня и гипокотиля в развивающемся зародыше было нанесено на карту с использованием клонального анализа (Scheres et al. 1994). Из трансгенных растений, несущих такую конструкцию, отбирали большие меченые клоны и анализировали границы этих клонов. Например, вырезание клетки зародыша, которое охватывает весь будущий гипокотиль, приведет к образованию гипокотильного сектора (1). Основная масса границ секторов либо совпадала, либо была тесно связана с делениями эмбриональных клеток. Это позволило построить карту, изображающую вероятную судьбу эмбриональных клеток, «карту судеб» ().
Это позволило построить карту, изображающую вероятную судьбу эмбриональных клеток, «карту судеб» ().
Открыть в отдельном окне
Рисунок 3А. Большой клон клеток, экспрессирующих β-глюкуронидазу, происходящий от одного предшественника, охватывает весь гипокотиль. Рисунок 3B. Клеточный клон, охватывающий зародышевую область корня, маркируется экспрессией β-глюкуронидазы.
Разделение между корневым и гипокотильным доменами совпало с делениями на ранней сердечной стадии эмбриогенеза, но секторы, отмечающие эти деления, иногда пересекали границу корень/гипокотиль, указывая на то, что они не разделяют судьбу корня и гипокотиля. Т.о., хотя и возможно картировать корневой домен у эмбрионов сердечной стадии, отдельные клетки могут реагировать на еще не идентифицированные позиционные сигналы и принимать судьбу либо корня, либо гипокотиля до последнего деления эмбриональной клетки.
Анализ также показал, что радиальное расположение клеток в корне устанавливается на стадии сердца и сохраняется после этого. Расположение инициальных клеток вокруг покоящегося центра, сохраняющееся в зрелом корне, получило название промеристемы (Clowes, 1954). Происхождение гипофиза правильное, без обнаруживаемых вариаций. Отмеченные производные гипофиза всегда дают начало центральным клеткам и колумелле, и секторов, включающих колумеллу/центральные клетки и другие типы клеток корня, не наблюдалось.
Расположение инициальных клеток вокруг покоящегося центра, сохраняющееся в зрелом корне, получило название промеристемы (Clowes, 1954). Происхождение гипофиза правильное, без обнаруживаемых вариаций. Отмеченные производные гипофиза всегда дают начало центральным клеткам и колумелле, и секторов, включающих колумеллу/центральные клетки и другие типы клеток корня, не наблюдалось.
Иссечение части зародышевого корня, находящейся вне меристемы, приводит к образованию сектора «зародышевого корня» (). Корневые клетки в этой области не окружены латеральным слоем корневого чехлика и не повторно инициируют клеточное деление постэмбрионально. Они образуют небольшую область соединения между корнем и гипокотилем с типичной эпидермальной морфологией и рано появляющимися корневыми волосками, «цангой».
На поздней сердечной стадии эмбриогенеза самая базальная эпидермальная клетка образовала слой латерального корневого чехлика путем периклинальных делений. Анатомический и клональный анализы показывают, что на этой стадии возникает типичный образец деления корневой меристемы, подобный стволовым клеткам. Клетки внутри развивающейся корневой меристемы, окруженные этим первым боковым слоем корневого чехлика, повторно инициируют клеточные деления после покоя семян и, таким образом, формируют постэмбриональную корневую меристему.
Клетки внутри развивающейся корневой меристемы, окруженные этим первым боковым слоем корневого чехлика, повторно инициируют клеточные деления после покоя семян и, таким образом, формируют постэмбриональную корневую меристему.
Открыть в отдельном окне
Боковой корневой чехлик (фиолетовый), корневой чехлик колумеллы (розовый) и покоящийся центр (серый) в зрелом зародыше. Латеральный слой корневого чехлика окружает клетки, которые митотически реактивируются при прорастании, давая начало корневой меристеме.
В совокупности клональный анализ эмбриона показывает, что регуляция точных моделей клеточных делений не требуется для развития проростка с его высокоорганизованной структурой. Это предполагает, что решения о клеточной судьбе принимаются независимо от наследственной истории вовлеченных клеток. Отсутствие роли клеточного происхождения в спецификации клеточной идентичности предполагает, что позиционная информация является важным фактором в спецификации клеточной идентичности.
Зачатки боковых корней возникают из клеток перицикла напротив полюсов ксилемы на некотором расстоянии от меристемы первичного корня (Dolan et al. 1993). Паттерны начальных клеточных делений, которые дают начало новым зачаткам, сильно отличаются от тех, которые происходят во время формирования первичного корня (Casimiro et al. 2001). На более поздних стадиях формирования боковых корней клеточная организация становится очень похожей на первичный корень, хотя боковые корни демонстрируют большую изменчивость в количестве клеток и точной клеточной организации. Анализ паттернов клеточного деления с помощью ловушек-энхансеров, которые маркируют различные ткани корня, использовался для определения стадии развития боковых корней (Malamy and Benfey, 19).97). Восемь стадий были определены на основе конкретных анатомических характеристик и клеточных делений. Экспрессию из маркерных линий, специфичных для типа клеток, использовали для идентификации типов клеток в развивающемся зачатке. Анализ паттернов экспрессии и паттернов клеточного деления показал, что значительная степень организации и дифференцировки клеток происходит на очень ранних стадиях развития зачатка бокового корня, при этом дифференциальная экспрессия генов проявляется после первой серии делений перицикла. Напротив, меристема бокового корня не становится активной до тех пор, пока зачаток не выйдет из родительского корня, и поэтому, по-видимому, не играет роли в формировании и организации раннего паттерна (Malamy and Benfey, 19).97). Это похоже на то, что наблюдается во время эмбриогенеза.
Напротив, меристема бокового корня не становится активной до тех пор, пока зачаток не выйдет из родительского корня, и поэтому, по-видимому, не играет роли в формировании и организации раннего паттерна (Malamy and Benfey, 19).97). Это похоже на то, что наблюдается во время эмбриогенеза.
Клональный анализ
Промеристема формируется во время эмбриогенеза. При прорастании клетки меристемы начинают делиться, и корень расширяется в осевом направлении в результате расширения клеток. По мере того, как корень продолжает расти, количество клеток в меристеме увеличивается, а скорость образования клеток увеличивается. Это увеличение количества клеток может объяснить удвоение скорости удлинения корней между 6 и 10 днями после прорастания. Клональный анализ использовался для изучения судеб клеток в различных частях меристемы во время роста корней молодых проростков (Kidner et al., 2000). Судьбы центральных («покоящийся центр») клеток и инициалей исследовали путем индукции клонов в 3-дневных меристемах с использованием индуцируемого нагреванием маркера на основе транспозона. Подвергая корни короткому тепловому шоку в течение 3 дней, можно было индуцировать вырезание транспозона из маркерного гена в небольшом количестве клеток и впоследствии исследовать их судьбу. Поскольку центральные клетки редко делятся, было идентифицировано несколько секторов, возникающих в результате событий вырезания в этих клетках. Те, что были замечены, указывали на то, что деление центральной клетки могло дать начало клеткам практически любого типа клеток корня — сектора, которые включали центральные клетки, также содержали клетки в различных тканях. Это указывает на то, что направление деления центральной клетки изменчиво. Кроме того, были обнаружены несколько коротких секторов, которые включали клетки в несколько слоев в радиальном измерении, что указывает на то, что деление некоторых клеток может привести к возникновению клеток в разных слоях. Это указывает на то, что, хотя схема клеточного деления в корне является стереотипной, вариации в общей схеме все же имеют место.
Подвергая корни короткому тепловому шоку в течение 3 дней, можно было индуцировать вырезание транспозона из маркерного гена в небольшом количестве клеток и впоследствии исследовать их судьбу. Поскольку центральные клетки редко делятся, было идентифицировано несколько секторов, возникающих в результате событий вырезания в этих клетках. Те, что были замечены, указывали на то, что деление центральной клетки могло дать начало клеткам практически любого типа клеток корня — сектора, которые включали центральные клетки, также содержали клетки в различных тканях. Это указывает на то, что направление деления центральной клетки изменчиво. Кроме того, были обнаружены несколько коротких секторов, которые включали клетки в несколько слоев в радиальном измерении, что указывает на то, что деление некоторых клеток может привести к возникновению клеток в разных слоях. Это указывает на то, что, хотя схема клеточного деления в корне является стереотипной, вариации в общей схеме все же имеют место.
Визуальное исследование немаркированных клонов также использовалось для демонстрации того, что позиционная информация регулирует спецификацию клеточной идентичности. Это основано на способности отслеживать характер клеточных делений в «пакетах» клеток, происходящих из одной клетки в эпидермисе. В этом случае «генетическая маркировка» не нужна. Редкие продольные деления клеток трихобластов приводят к образованию двойных клеточных массивов. Как правило, один из этих близнецов находится в положении трихобласта над двумя клетками коры и развивается как трихобласт (Berger et al., 19).98б). Другой будет находиться в положении атрихобласта, перекрывая единственную корковую клетку при просмотре в поперечном сечении, и предполагает идентичность атрихобласта. Если бы положение определяло судьбу клетки, то можно было бы ожидать, что каждый из близнецов примет судьбу, характерную для их положения – трихобласты будут формироваться в месте расположения трихобластов. С другой стороны, если бы родословная была важна, то оба близнеца развивались бы как трихобласты, поскольку они произошли от трихобласта. Поскольку близнецовые клетки дифференцируются в трихобласты и атрихобласты в зависимости от положения, это предполагает, что позиционные сигналы регулируют паттерн дифференцировки в эпидермисе.
Это основано на способности отслеживать характер клеточных делений в «пакетах» клеток, происходящих из одной клетки в эпидермисе. В этом случае «генетическая маркировка» не нужна. Редкие продольные деления клеток трихобластов приводят к образованию двойных клеточных массивов. Как правило, один из этих близнецов находится в положении трихобласта над двумя клетками коры и развивается как трихобласт (Berger et al., 19).98б). Другой будет находиться в положении атрихобласта, перекрывая единственную корковую клетку при просмотре в поперечном сечении, и предполагает идентичность атрихобласта. Если бы положение определяло судьбу клетки, то можно было бы ожидать, что каждый из близнецов примет судьбу, характерную для их положения – трихобласты будут формироваться в месте расположения трихобластов. С другой стороны, если бы родословная была важна, то оба близнеца развивались бы как трихобласты, поскольку они произошли от трихобласта. Поскольку близнецовые клетки дифференцируются в трихобласты и атрихобласты в зависимости от положения, это предполагает, что позиционные сигналы регулируют паттерн дифференцировки в эпидермисе.
Лазерная абляция
Клональный анализ впервые показал, что стереотипные клеточные деления внутри зародыша и корня арабидопсиса не являются существенными для поддержания ранних решений клеточной судьбы. Скорее, клетки остаются способными адаптировать свою судьбу в соответствии со своим положением. В клональном анализе, особенно при сосредоточении внимания на относительно редких делениях, причина изменений плоскости деления, которые приводят к необычным клонам, неизвестна и может формально отражать более раннюю нестабильность судьбы, что затрудняет распутывание причины и следствия. Вот почему вывод о том, что позиционная передача сигналов имеет первостепенное значение в меристеме постэмбрионального корня, был расширен другим методом: использованием лазерной абляции, которая приводит к расширению и последующему делению клеток на соседние слои (Van den Berg et al. 1995; Бергер и др. 1998). В абляционных анализах причина смещения клеток известна — она индуцируется экспериментально — и, следовательно, нет отбора «нестабильных» клеток, но соседние клеточные трупы могли бы формально изменить ответные свойства клеток, меняющих положение. Независимо от этого возможного предостережения, абляция и последующее перераспределение меристематических клеток вдоль радиальной оси и вдоль апикально-базальной оси приводит к соответствующим изменениям клеточных судеб. Это снова указывает на то, что клеточные судьбы зависят как от радиальной, так и от апикально-базальной позиционной передачи сигналов. Таким образом, все доступные данные с использованием подходов с различными преимуществами и недостатками подтверждают один и тот же вывод о том, что позиционная информация постоянно интерпретируется клетками меристемы корня.
Независимо от этого возможного предостережения, абляция и последующее перераспределение меристематических клеток вдоль радиальной оси и вдоль апикально-базальной оси приводит к соответствующим изменениям клеточных судеб. Это снова указывает на то, что клеточные судьбы зависят как от радиальной, так и от апикально-базальной позиционной передачи сигналов. Таким образом, все доступные данные с использованием подходов с различными преимуществами и недостатками подтверждают один и тот же вывод о том, что позиционная информация постоянно интерпретируется клетками меристемы корня.
Какова природа позиционной информации, определяющей дистальную спецификацию верхушки корня? При абляции клеток покоящегося центра проваскулярные клетки в апикально-базальной оси (мы используем апикальные и базальные, как у эмбриона, и, следовательно, апикальные находятся на проксимальной стороне кончика корня, а базальные — на дистальном конце) переопределяются на формируют корневой чехлик колумеллы и покоящиеся центральные клетки (Van den Berg et al. 1997) (и ). Кончики корней накапливают ауксины, а накопление синтетического чувствительного к ауксину промоторного элемента показывает максимум сразу под QC (панель A). Дистальная респецификация, наблюдаемая после лазерной абляции QC, предвещается новым максимумом экспрессии этого промоторного элемента. Респецификация не происходит в корнях, обработанных низкими уровнями полярных ингибиторов транспорта ауксина, или в доминантных мутантов axr3 (Sabatini et al. 1999), которые стабилизируют регулятор транскрипции AXR3/IAA17 ауксиновых ответов (Rouse et al. 1998; Ouellet et al. 2001). Т.о., накопление ауксина на кончике корня необходимо для спецификации, зависящей от дистального положения. Изменение локализации паттерна ответа ауксина при обработке высокими дозами ингибиторов полярного транспорта приводит к спецификации QC, колумеллы и латерального слоя корневого чехлика во всей меристеме, предполагая, что восприятие высоких уровней ауксина не только необходимо, но и достаточно.
1997) (и ). Кончики корней накапливают ауксины, а накопление синтетического чувствительного к ауксину промоторного элемента показывает максимум сразу под QC (панель A). Дистальная респецификация, наблюдаемая после лазерной абляции QC, предвещается новым максимумом экспрессии этого промоторного элемента. Респецификация не происходит в корнях, обработанных низкими уровнями полярных ингибиторов транспорта ауксина, или в доминантных мутантов axr3 (Sabatini et al. 1999), которые стабилизируют регулятор транскрипции AXR3/IAA17 ауксиновых ответов (Rouse et al. 1998; Ouellet et al. 2001). Т.о., накопление ауксина на кончике корня необходимо для спецификации, зависящей от дистального положения. Изменение локализации паттерна ответа ауксина при обработке высокими дозами ингибиторов полярного транспорта приводит к спецификации QC, колумеллы и латерального слоя корневого чехлика во всей меристеме, предполагая, что восприятие высоких уровней ауксина не только необходимо, но и достаточно. дистализация корневой меристемы ().
дистализация корневой меристемы ().
Открыть в отдельном окне
Лазерная абляция центра покоя в постэмбриональном корне. Изображения конфокального сканирования из одного корня. Клетки очерчены окрашиванием йодидом пропидия, выполненным сразу после абляции (день 0) и перед визуализацией в последующие дни. Клетки колумеллы выделяются экспрессией GFP. В день 0 успешная абляция отмечается поступлением йода пропидия в клетки QC. Эти клетки сжимаются и в последующие дни располагаются более дистально из-за дифференцировки и отслоения корневого чехлика. Маркер колумеллы смещается и вновь экспрессируется в бывших проваскулярных клетках. Через 5 дней эти клетки образуют новую колумеллу, и вскоре их клеточная организация будет неотличима от необработанных корней.
Открыть в отдельном окне
Лазерная абляция как в . Слияние промотора SCARECROW с GFP используется для визуализации повторного появления клеток покоящегося центра в проваскулярном домене через 3 дня после абляции и далее.
Открыть в отдельном окне
Рисунок 7А. Синтетический реагирующий на ауксин элемент DR5::GUS маркирует самые дистальные клетки кончика корня, и его расположение резко меняется при добавлении полярных ингибиторов транспорта ауксина, таких как нафтилфталаминовая кислота (NPA). Обработка NPA также изменяет размер и форму домена корневого чехлика колумеллы, что визуализируется здесь по их характерным гранулам крахмала. Рисунок 7В. Модель поддержания стволовых клеток, окружающих покоящийся центр, с помощью сигналов ближнего действия, которые подавляют прогрессирование клеточной дифференцировки.
Важным аспектом дистального кончика корня является то, что он содержит все зачатки меристем, которые действуют как стволовые клетки для различных клеточных линий. Абляция одиночных клеток QC приводит к потере статуса стволовых клеток и прогрессированию дифференцировки контактирующих исходных клеток (Van den Berg et al. 1997). Наиболее ярко это проявляется в прекращении деления клеток и приобретении зерен крахмала инициалями корневого чехлика колумеллы. Воздействуют только инициали, контактирующие с аблированными клетками QC, что свидетельствует о том, что сигналы ближнего действия, исходящие от QC, поддерживают активность стволовых клеток окружающих инициалей ()). Интересно отметить, что аналогичная функция поддержания стволовых клеток была предложена для клеток под слоями стволовых клеток в апикальной меристеме побега (Mayer et al, 2000).
Воздействуют только инициали, контактирующие с аблированными клетками QC, что свидетельствует о том, что сигналы ближнего действия, исходящие от QC, поддерживают активность стволовых клеток окружающих инициалей ()). Интересно отметить, что аналогичная функция поддержания стволовых клеток была предложена для клеток под слоями стволовых клеток в апикальной меристеме побега (Mayer et al, 2000).
Изоляция исходных клеток основной ткани от более зрелых клеток основной ткани путем удаления их апикально расположенных дочерних клеток препятствует изменению ориентации плоскости клеточного деления, необходимому для формирования новых энтодермальных и кортикальных клеточных массивов. Таким образом, для асимметричного клеточного деления необходима связность клеточного слоя. Это можно интерпретировать как отражение потока позиционной информации от более зрелых клеток к стволовым клеткам как механизма поддержания информации о формировании радиального паттерна (Van den Berg et al. 19).95). Однако, поскольку QC задерживает дифференцировку или способствует судьбе стволовых клеток, эти наблюдения могут также отражать накопление факторов, стимулирующих стволовые клетки (Van den Berg et al. 1997). Абляции основной ткани и QC необходимо будет повторить с новыми доступными маркерами клеточной судьбы и статуса клеточной дифференцировки, чтобы различать эти возможности. Лазерная абляция также использовалась для анализа природы передачи сигналов между клетками, участвующей в спецификации трихобластов (Berger et al. 19).98). Интересно, что клеточная судьба не изменялась, когда связь с живыми соседними клетками была нарушена, что позволяет предположить, что позиционные сигналы имеют внеклеточную природу.
1997). Абляции основной ткани и QC необходимо будет повторить с новыми доступными маркерами клеточной судьбы и статуса клеточной дифференцировки, чтобы различать эти возможности. Лазерная абляция также использовалась для анализа природы передачи сигналов между клетками, участвующей в спецификации трихобластов (Berger et al. 19).98). Интересно, что клеточная судьба не изменялась, когда связь с живыми соседними клетками была нарушена, что позволяет предположить, что позиционные сигналы имеют внеклеточную природу.
Было показано, что многие гены участвуют в различных аспектах развития корня (). Ниже их роль будет обсуждаться в пяти основных контекстах: формирование дистального паттерна, формирование радиального паттерна, формирование эпидермального паттерна, клеточное деление и экспансия клеток.
Таблица 1.
Гены, участвующие в развитии корня арабидопсиса
Открыть в отдельном окне
Многочисленные физиологические эксперименты продемонстрировали, что формирование целых корневых систем может стимулироваться ауксинами, тогда как вышеупомянутые эксперименты на Arabidopsis сосредоточены на потребности в ауксине для дистальной спецификации. Анализ мутантов с нарушением корнеобразования дает дополнительные доказательства роли ауксинов в формировании всего корня, а также в спецификации его дистальных элементов.
Анализ мутантов с нарушением корнеобразования дает дополнительные доказательства роли ауксинов в формировании всего корня, а также в спецификации его дистальных элементов.
Мутации в нескольких генах препятствуют формированию зародышевых корней. 9У мутантов 0036 monopteros (mp) полностью отсутствует зародышевый корень (Berleth, Jurgens, 1993). Ген MP кодирует член белков AUXIN RESPONSE FACTOR, которые могут обеспечивать быстрые транскрипционные ответы на ауксин (Hardtke et al. 1998). Мутанты bodenlos (bdl) и ауксин-резистентные6 (axr6) обнаруживают сходные ранние эмбриональные фенотипы и тяжелые дефекты первичных корней (Hamann et al. 1999; Hobbie et al. 2000). Оба этих мутанта проявляют повышенную устойчивость к внешне добавляемому ауксину, а 9Мутант 0036 bdl генетически взаимодействует с мутантами mp (Hamann et al. 1999). Таким образом, мутации по крайней мере в трех генах указывают на участие ауксинов в формировании корней зародыша. Кроме того, более тонкие дефекты постэмбриональной корневой структуры и/или дифференцировки клеток наблюдались у ауксин-резистентных1 (axr1) и транспортных . устойчивых к ингибиторам1 (tir1) мутантов , что согласуется с их предполагаемой функцией в модуляции ауксиновых ответов посредством направленного протеолиза (Leyser et al. 1993; Рюггер и др. 1998).
Кроме того, более тонкие дефекты постэмбриональной корневой структуры и/или дифференцировки клеток наблюдались у ауксин-резистентных1 (axr1) и транспортных . устойчивых к ингибиторам1 (tir1) мутантов , что согласуется с их предполагаемой функцией в модуляции ауксиновых ответов посредством направленного протеолиза (Leyser et al. 1993; Рюггер и др. 1998).
Мутации в гене HOBBIT (HBT) также мешают корнеобразованию. Постэмбриональная меристемная активность отсутствует, и дистальные типы клеток (QC, колумелла и латеральный корневой чехлик) не дифференцируются (Willemsen et al. 1998). Самый ранний дефект, обнаруженный у мутантов hbt , представляет собой нарушение плоскостей клеточного деления в гипофизе, клетке-предшественнике QC и колумелле. Этот дефект возникает в более ограниченном домене зародыша, а также на более поздней стадии, чем mp , bdl и axr6 дефекты. Мутанты hbt реагируют на IAA, но синтетический элемент ответа на ауксин DR5 может индуцироваться только при высоких концентрациях ауксина, указывая на то, что транскрипционные ответы на ауксин десенсибилизированы на мутантном фоне (Blilou et al. , in prep). Ген HBT кодирует гомолог субъединицы дрожжевого комплекса, стимулирующего анафазу (Blilou et al., в процессе подготовки), и может быть необходим в зачаточном зачатке корня для обеспечения ауксин-зависимого развития корня, а на более поздних стадиях – для обеспечения ауксин-зависимого развития корня. зависимая дифференцировка дистальных типов клеток.
, in prep). Ген HBT кодирует гомолог субъединицы дрожжевого комплекса, стимулирующего анафазу (Blilou et al., в процессе подготовки), и может быть необходим в зачаточном зачатке корня для обеспечения ауксин-зависимого развития корня, а на более поздних стадиях – для обеспечения ауксин-зависимого развития корня. зависимая дифференцировка дистальных типов клеток.
Компоненты предполагаемых переносчиков оттока, которые необходимы для полярного транспорта ауксина, могут также обнаруживать тонкие дефекты в полюсе зародышевого корня. Напр., мутанты pin4 обнаруживают аберрантные деления клеток гипофиза и изменения в распределении максимума репортерного ауксина в кончике корня (Friml et al. 2002). Мутации в гене GNOM , кодирующем белок ARF-GEF, участвующий в перемещении везикул, нарушают координатную полярную локализацию по крайней мере одного из носителей оттока (Steinmann et al. 19).99). Будет интересно выяснить, лежат ли дефекты транспорта полярных ауксинов в основе неспособности проростков gnom образовывать корневой зачаток.
Хотя было идентифицировано много генов, которые связывают развитие корней с ауксинами, еще не сообщалось о факторах транскрипции, которые определяют различные дистальные типы клеток. Несколько факторов класса MADS box (Alvarez-Buylla et al. 2000) и два транскрипционных фактора типа AP2 (Aida et al, в процессе подготовки) в настоящее время исследуются, и новые инструменты постгеномной эры могут помочь в идентификации других.
У мутантов tornado ( trn1 и trn2 ) формирующие отделы начального эпидермального/латерального корневого чехлика дефектны. Более того, клетки с атрибутами латерального корневого чехлика обнаруживаются в самом внешнем клеточном слое, который в норме содержит только эпидермальные клетки (Cnops et al., 2000). Это указывает на то, что TRN1 и TRN2 необходимы для правильной клеточной спецификации в самом внешнем слое клеток и что роль этих генных продуктов заключается в репрессии судьбы латерального корневого чехлика в клетках эпидермального расположения. Молекулярная природа продуктов гена TRN пока неизвестна. Эпидермальная судьба меняется в 9Мутанты 0036 trn могут указывать на то, что спецификация латерального корневого чехлика как элемента дистального паттерна включает взаимодействие с эпидермальными факторами, которые сами по себе зависят от событий формирования радиального паттерна.
Молекулярная природа продуктов гена TRN пока неизвестна. Эпидермальная судьба меняется в 9Мутанты 0036 trn могут указывать на то, что спецификация латерального корневого чехлика как элемента дистального паттерна включает взаимодействие с эпидермальными факторами, которые сами по себе зависят от событий формирования радиального паттерна.
Если развитие первичного корня сильно зависит от фитогормона ауксина, кажется разумным ожидать аналогичной важной роли ауксинов в инициации боковых корней. Действительно, генетический анализ образования боковых корней также выявил многочисленные связи с ауксинами. Добавление экзогенного ауксина приводит к образованию нештатных корней, и 9Мутанты 0036 superroot (также называемые аберрантным образованием боковых корней1 и без крючков3 ) демонстрируют сходный фенотип. (Буржан и др., 1995; Селенца и др., 1995). Мутанты, устойчивые к ауксину, имеют уменьшенное количество боковых корней (Hobbie and Estelle, 1995; Timpte et al, 1995). Индуцируемый ауксином транскрипционный фактор NAC1, чья активность определяет число зачатков боковых корней, действует ниже tir1 -зависимых ауксиновых ответов (Xie et al. 2000). Образование боковых корней не стимулируется ИУК в аберрантное формирование боковых корней4 (alf4) мутантов, указывая на то, что соответствующий ген также действует ниже начального IAA-связанного стимула для образования боковых корней (Celenza et al. 1995). Наконец, мутант с одиночным корнем со специфическим дефектом формирования боковых корней несет доминантную мутацию в гене, кодирующем член семейства белков AUX-IAA, что указывает на особую роль этого конкретного члена в индукции боковых корней (Fukaki et al. , 2002). Многие мутанты и соответствующие гены, обсуждаемые в настоящее время, обеспечивают возможность анализа механизмов, с помощью которых ауксины, по-видимому, способствуют развитию боковых корней. Будет интересно определить степень перекрытия и специфические различия между индуктивными процессами в боковых корнях и процессами, действующими в зародыше.
Индуцируемый ауксином транскрипционный фактор NAC1, чья активность определяет число зачатков боковых корней, действует ниже tir1 -зависимых ауксиновых ответов (Xie et al. 2000). Образование боковых корней не стимулируется ИУК в аберрантное формирование боковых корней4 (alf4) мутантов, указывая на то, что соответствующий ген также действует ниже начального IAA-связанного стимула для образования боковых корней (Celenza et al. 1995). Наконец, мутант с одиночным корнем со специфическим дефектом формирования боковых корней несет доминантную мутацию в гене, кодирующем член семейства белков AUX-IAA, что указывает на особую роль этого конкретного члена в индукции боковых корней (Fukaki et al. , 2002). Многие мутанты и соответствующие гены, обсуждаемые в настоящее время, обеспечивают возможность анализа механизмов, с помощью которых ауксины, по-видимому, способствуют развитию боковых корней. Будет интересно определить степень перекрытия и специфические различия между индуктивными процессами в боковых корнях и процессами, действующими в зародыше.
Радиальная организация корня создается стереотипным делением исходных клеток и последующим приобретением клеточной судьбы. На поперечном срезе корня выделяют четыре радиально-симметричных слоя (снаружи внутрь, эпидермис, кора, эндодерма, перицикл), которые окружают билатерально-симметричную сосудистую ткань (состоит из флоэмы, ксилемы и прокамбия) (Scheres et al., 1995) (Scheres et al., 1995). ). Сосудистая ткань и окружающий перицикл называются стелой. Мутации, нарушающие радиальный паттерн, оказались полезными для идентификации генов, играющих важную роль в установлении и поддержании паттерна. Многие из этих мутаций были выделены путем скрининга корней, которые больше не могли расти неопределенным образом. В результате этих скринингов были идентифицированы мутации, нарушающие формирование паттерна основной ткани и сосудистого цилиндра (Scheres et al., 19).95).
Открыть в отдельном окне
Радиальная организация корней арабидопсиса. У мутанта чучело отсутствует один клеточный слой, и полученный мутантный слой имеет дифференцированные признаки как коры, так и эндодермы.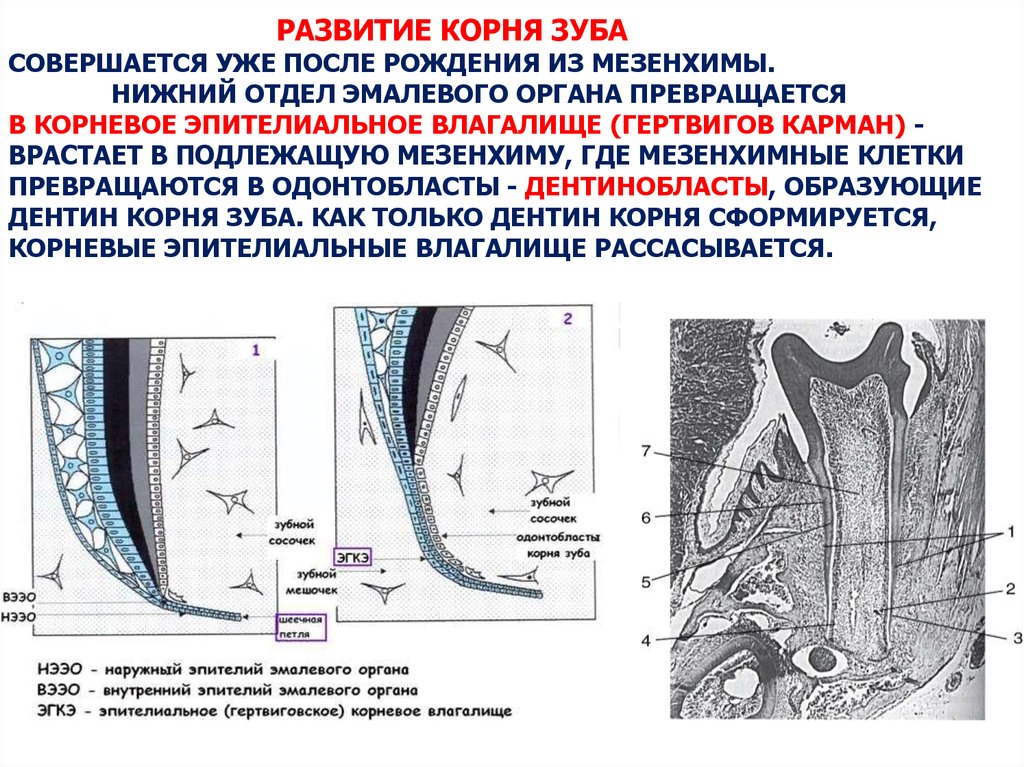 У мутанта с коротким корнем также отсутствует клеточный слой, но его мутантный слой имеет признаки только коры.
У мутанта с коротким корнем также отсутствует клеточный слой, но его мутантный слой имеет признаки только коры.
Как у чучела ( scr ), так и у короткокорневых мутантов вместо коры и эндодермы имеется один мутантный слой между эпидермисом и стелой (Scheres et al., 1995) (). Анализ тканеспецифических маркеров показал, что мутантный слой у двух мутантов специфицирован по-разному. В scr в мутантном слое присутствуют маркеры как коры, так и эндодермы, что указывает на то, что SCR необходим для периклинального деления исходной клетки, но не играет роли в спецификации клеток (Di Laurenzio et al., 1996). (). В shr обнаружены только маркеры коры, что указывает на то, что SHR необходим как для продольного клеточного деления начальной, так и для спецификации энтодермальных клеток (Di Laurenzio et al., 19).96) (). Оба гена кодируют членов семейства GRAS предполагаемых регуляторов транскрипции. SCR транскрибируется в кортексе/энтодермальных инициалях, его дочерних клетках и во всех энтодермальных клетках, а также в QC (Di Laurenzio et al. , 1996) (4). Поскольку экспрессия SCR подавляется на фоне мутантного shr , был сделан вывод, что SHR необходим для полной активации транскрипции SCR (Helariutta et al., 2000). Однако SHR РНК обнаруживается исключительно в клетках стел (1), что указывает на то, что она действует вне клетки автономно. Иммунолокализация белка SHR, а также слияния SHR с зеленым флуоресцентным белком (GFP) показали, что SHR может перемещаться из стелы в соседний клеточный слой. Белок SHR обнаруживается не только в стеле, но и во всех клетках соседнего слоя (), включая кору/инициальную энтодерму, ее дочернюю часть, энтодерму и QC (Nakajima et al., 2001). Эти результаты указывают на то, что белок SHR способен транслоцироваться из стелы в клетки соседнего слоя. Хотя точный механизм еще не определен, было высказано предположение, что SHR способен перемещаться через плазмодесмы. Движение, по-видимому, строго регулируется, так как нет никаких признаков присутствия белка SHR в клетках, находящихся вне соседнего слоя.
, 1996) (4). Поскольку экспрессия SCR подавляется на фоне мутантного shr , был сделан вывод, что SHR необходим для полной активации транскрипции SCR (Helariutta et al., 2000). Однако SHR РНК обнаруживается исключительно в клетках стел (1), что указывает на то, что она действует вне клетки автономно. Иммунолокализация белка SHR, а также слияния SHR с зеленым флуоресцентным белком (GFP) показали, что SHR может перемещаться из стелы в соседний клеточный слой. Белок SHR обнаруживается не только в стеле, но и во всех клетках соседнего слоя (), включая кору/инициальную энтодерму, ее дочернюю часть, энтодерму и QC (Nakajima et al., 2001). Эти результаты указывают на то, что белок SHR способен транслоцироваться из стелы в клетки соседнего слоя. Хотя точный механизм еще не определен, было высказано предположение, что SHR способен перемещаться через плазмодесмы. Движение, по-видимому, строго регулируется, так как нет никаких признаков присутствия белка SHR в клетках, находящихся вне соседнего слоя.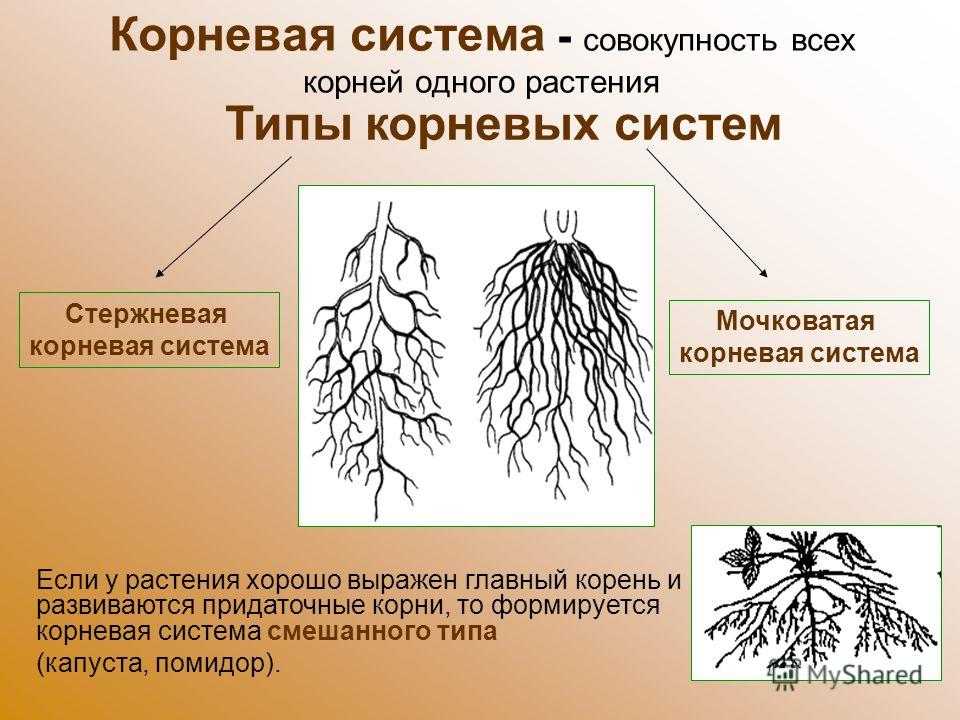 Более того, в стеле SHR локализуется как в ядре, так и в цитоплазме, но однажды перемещаясь в энтодерму, он обнаруживается исключительно в ядре (1) (Nakajima et al., 2001). Эти результаты показывают, что SHR действует как активатор клеточного деления и индуктор клеточной судьбы, а также как позиционный сигнал.
Более того, в стеле SHR локализуется как в ядре, так и в цитоплазме, но однажды перемещаясь в энтодерму, он обнаруживается исключительно в ядре (1) (Nakajima et al., 2001). Эти результаты показывают, что SHR действует как активатор клеточного деления и индуктор клеточной судьбы, а также как позиционный сигнал.
Открыть в отдельном окне
а) Характер экспрессии РНК SCARECROW , определенный с помощью гибридизации in situ (Di Laurenzio et al., 1996). b) Экспрессия, обеспечиваемая промотором SCR , слитым с GFP (Wysocka-Diller et al., 2000).
Открыть в отдельном окне
а) Экспрессия, обеспечиваемая промотором SHR , слитым с GFP. B) Экспрессия в результате трансляционного слияния SHR с GFP, управляемая промотором SHR , введенным в shr-2 мутантный фон (Nakajima et al., 2001).
Эктопическая экспрессия SHR, управляемая конститутивным промотором 35S, приводит к дополнительным клеточным слоям с измененными клеточными судьбами (Helariutta et al. , 2000). Когда экспрессия SHR управляется промотором SCR, наблюдается очень специфическое увеличение основных слоев ткани (Nakajima et al., 2001). В сильно экспрессирующих линиях эти эктопические клетки основной ткани экспрессируют энтодермальные маркеры. Существует тесная корреляция между присутствием белка SHR и судьбой энтодермальных клеток, указывая на то, что SHR может быть достаточно, чтобы индуцировать спецификацию энтодермальных клеток. Знакомство с 9Мутация 0036 scr в этих линиях продемонстрировала, что клеточные деления, которые продуцируют дополнительные слои, зависят от активности SCR (Nakajima et al., 2001).
, 2000). Когда экспрессия SHR управляется промотором SCR, наблюдается очень специфическое увеличение основных слоев ткани (Nakajima et al., 2001). В сильно экспрессирующих линиях эти эктопические клетки основной ткани экспрессируют энтодермальные маркеры. Существует тесная корреляция между присутствием белка SHR и судьбой энтодермальных клеток, указывая на то, что SHR может быть достаточно, чтобы индуцировать спецификацию энтодермальных клеток. Знакомство с 9Мутация 0036 scr в этих линиях продемонстрировала, что клеточные деления, которые продуцируют дополнительные слои, зависят от активности SCR (Nakajima et al., 2001).
Открыть в отдельном окне
а) Продольный оптический срез трансгенных растений, экспрессирующих SHR из промотора SCR. Обратите внимание на дополнительные слои наземной ткани. б) Поперечный срез корней тех же растений (Nakajima et al., 2001).
У мутанта деревянной ноги ( wol ) протоксилема является единственной тканью в сосудистом цилиндре (сравните с диким типом у ) (Scheres et al. , 1995) (Махонен и др., 2000). В норме флоэма и прокамбий образуются за счет набора асимметричных клеточных делений их инициалей. Для этих делений необходим ген WOODEN LEG ( WOL ), который экспрессируется в сосудистой ткани корня, начиная с ранних стадий эмбриогенеза (Mahonen et al., 2000). Ген WOL кодирует новую двухкомпонентную гистидинкиназу. Имеются данные о том, что продукт гена WOL способен связывать цитокинин, что повышает вероятность того, что этот фитогормон играет непосредственную роль в регуляции формирования паттерна сосудистой сети (Inoue et al., 2001).
, 1995) (Махонен и др., 2000). В норме флоэма и прокамбий образуются за счет набора асимметричных клеточных делений их инициалей. Для этих делений необходим ген WOODEN LEG ( WOL ), который экспрессируется в сосудистой ткани корня, начиная с ранних стадий эмбриогенеза (Mahonen et al., 2000). Ген WOL кодирует новую двухкомпонентную гистидинкиназу. Имеются данные о том, что продукт гена WOL способен связывать цитокинин, что повышает вероятность того, что этот фитогормон играет непосредственную роль в регуляции формирования паттерна сосудистой сети (Inoue et al., 2001).
Открыть в отдельном окне
a) Клетки в сосудистой ткани дикого типа b) В сосудистой ткани деревянной ноги обнаружены только протоксилемные клетки (Scheres et al. 1995?).
Спецификация клеточной идентичности
Эпидермис корня состоит из двух типов клеток, идентичность которых регулируется информацией о положении. Трихобласты развиваются в волосковые клетки и располагаются в расщелине между нижележащими клетками коры (если смотреть в поперечном сечении), тогда как атрихобласты остаются безволосыми и располагаются над отдельными клетками коры (Dolan et al. , 19).94; Голуэй и др., 1994). Эксперименты по лазерной абляции и клональный анализ указывают на то, что позиционные сигналы направляют клеточную идентификацию, и есть некоторые свидетельства того, что эти позиционные сигналы расположены в клеточной стенке (Berger et al., 1998b).
, 19).94; Голуэй и др., 1994). Эксперименты по лазерной абляции и клональный анализ указывают на то, что позиционные сигналы направляют клеточную идентификацию, и есть некоторые свидетельства того, что эти позиционные сигналы расположены в клеточной стенке (Berger et al., 1998b).
Идентичность клеток регулируется в эпидермисе каскадом регуляторов транскрипции. GLABRA 2 является положительным регулятором судьбы атрихобластов/неволосковых клеток и экспрессируется в атрихобластах/неволосковых клетках (Masucci and Schiefelbein). GLABRA 2 положительно регулируется WEREWOLF, предполагаемым связанным с MYB регулятором транскрипции, который экспрессируется в атрихобластах/не волосковых клетках, и TRANSPARENT TESTA GLABRA, белком с повторами WD, который, как предполагается, опосредует белок-белковые взаимодействия (Lee and Schiefelbein 19).99; Голуэй и др., 1994). Судьба трихобластов положительно регулируется Myb-родственным белком CAPRICE (Wada et al. 1997). Было высказано предположение, что баланс между двумя белками, родственными Myb, определяет судьбу клетки; высокие значения WER/CPC в атрихобластах/неволосяных эпидермальных клетках и низкие значения WER/CPC в трихобластах/волосяных клетках (Lee and Schiefelbein, 1999) (полное обсуждение см. в главе N).
в главе N).
После того, как волосковые клетки определены, инициируются волоски с внешней стороны волосковой клетки, ближайшей к меристеме. Полярная локализация волосковой клетки зависит от пути этилена и ауксина и совпадает с установлением высокого уровня Ca 9Градиент 0284 2+ на кончике волоса (Schiefelbein et al., 1992; Wymer et al. 1997). Устанавливается рост кончиков и волосы удлиняются. Хотя было идентифицировано большое количество мутантов с дефектами роста волос, на сегодняшний день подробно охарактеризовано лишь немногие из них (Schiefelbein and Somerville, 1990; Schneider et al., 1997; Parker et al., 2000). ROOT HAIRLESS1 — это ядерный белок, необходимый для образования волос (Schneider et al., 1998). ROOT HAIR DEFECTIVE3 представляет собой небольшой G-белок, необходимый для контроля направления роста кончиков (Wang et al., 19).96). KOJAK/AtCSLD3 необходим для синтеза полисахарида клеточной стенки (Favery et al., 2001; Wang et al., 2001). LEUCINE RICH EXPANSIN1 (LRX1) представляет собой белок клеточной поверхности, необходимый для поддержания формы клетки (Baumberger et al. , 2001). TINY ROOT HAIR3 является переносчиком калия, необходимым во время инициации (Rigas et al., 2001). Вместе эти молекулярные и генетические анализы способствуют построению модели роста волосковых клеток.
, 2001). TINY ROOT HAIR3 является переносчиком калия, необходимым во время инициации (Rigas et al., 2001). Вместе эти молекулярные и генетические анализы способствуют построению модели роста волосковых клеток.
В меристеме образуются новые клетки, в которых очевидны отчетливые зоны активности клеточного деления. Клетки центрального «покоящегося центра» делятся нечасто, и скорость клеточного деления постепенно увеличивается вверх по корню, пока не будет достигнут максимум в точке, которая зависит от возраста корня и условий роста (Beemster and Baskin, 19).98). Путем измерения включения нуклеотидного аналога бромдезоксиуридина (BrdU) в реплицирующуюся ДНК было показано, что 7% клеток включили предшественник в ядра центральных клеток за 24-часовой период (Fujie et al., 1993). Инициальные клетки, которые окружают центральные клетки, чаще включают предшественники – 28% таких клеток включают предшественников. 84% ядер в клетках, расположенных на расстоянии двух клеток от центральных клеток, включали предшественники ДНК. Это согласуется с тем, что центральные клетки имеют очень низкую скорость клеточного деления с увеличением скорости деления в клетках на большем расстоянии от центральных клеток. Скорость клеточного деления увеличивается от центральных клеток к точке позади кончика, которая меняется по мере старения корня. В 6-дневных меристемах пик клеточного деления наблюдается примерно в 600 мкм от центральных клеток, в то время как в более старых корнях пик находится дальше от кончика примерно на 1000 мкм.
Это согласуется с тем, что центральные клетки имеют очень низкую скорость клеточного деления с увеличением скорости деления в клетках на большем расстоянии от центральных клеток. Скорость клеточного деления увеличивается от центральных клеток к точке позади кончика, которая меняется по мере старения корня. В 6-дневных меристемах пик клеточного деления наблюдается примерно в 600 мкм от центральных клеток, в то время как в более старых корнях пик находится дальше от кончика примерно на 1000 мкм.
Деление клеток в эпидермисе регулируется подмножеством генов, которые регулируют идентичность клеток
Трихобласты короче атрихобластов на протяжении всего своего развития за счет меристемы, зон элонгации и дифференцировки Dolan et al., 1994; Голуэй и др., 1994). Разница в длине клеток видна уже вблизи промеристемы и в дальнейшем сохраняется за счет дифференцировки (Beemster and Baskin, 1998). Поскольку два типа клеток различаются по длине от раннего развития до зрелых стадий, скорость клеточного деления в меристематической зоне одинакова для обоих типов клеток. Тем не менее, разница в размерах клеток между двумя типами клеток постоянно регулируется. В то время как большинство клеточных делений в эпидермисе являются поперечными, добавляя больше клеток к каждому файлу, иногда происходят продольные деления, которые приводят к образованию пары клеток рядом (близнецов), что приводит к дублированию файла (Berger et al., 1998). Обе дочерние клетки имеют одинаковую длину, но после нескольких циклов деления клетки в положении трихобласта, полученные от одного из близнецов, становятся короче, чем клетки, полученные из другого близнеца в положении атрихобласта. Чтобы клетки в положении трихобластов были короче клеток в положении атрихобластов, скорость клеточного деления должна быть выше в трихобластах, чем в атрихобластах. Поэтому при переключении позиций изменяются параметры клеточного деления, в результате чего образуются клетки соответствующих размеров. Это указывает на то, что относительный размер клеток и скорость деления клеток строго регулируются в эпидермисе.
Тем не менее, разница в размерах клеток между двумя типами клеток постоянно регулируется. В то время как большинство клеточных делений в эпидермисе являются поперечными, добавляя больше клеток к каждому файлу, иногда происходят продольные деления, которые приводят к образованию пары клеток рядом (близнецов), что приводит к дублированию файла (Berger et al., 1998). Обе дочерние клетки имеют одинаковую длину, но после нескольких циклов деления клетки в положении трихобласта, полученные от одного из близнецов, становятся короче, чем клетки, полученные из другого близнеца в положении атрихобласта. Чтобы клетки в положении трихобластов были короче клеток в положении атрихобластов, скорость клеточного деления должна быть выше в трихобластах, чем в атрихобластах. Поэтому при переключении позиций изменяются параметры клеточного деления, в результате чего образуются клетки соответствующих размеров. Это указывает на то, что относительный размер клеток и скорость деления клеток строго регулируются в эпидермисе.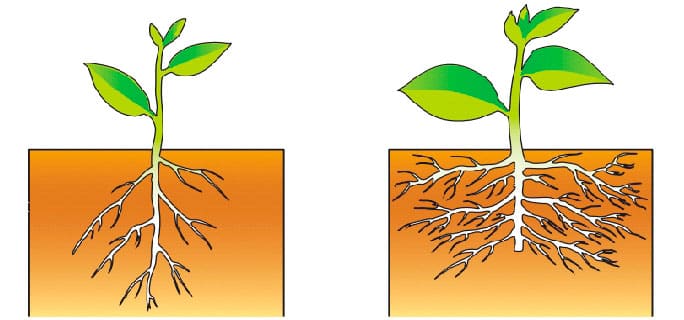
Продольные деления, в результате которых образуются клетки-близнецы, приводящие к дупликации файлов, происходят преимущественно в клетках в положении трихобластов. Ограничение этих продольных делений трихобластами контролируется подмножеством генов, которые регулируют спецификацию типов клеток в эпидермисе (глава Schiefelbein and Grierson). У мутантов Transparent testa glabra ( ttg ) на каждой клетке эпидермиса появляются волоски, но также происходит продольное деление как в атрихобластах, так и в трихобластах, что указывает на то, что ТТГ также необходим для подавления продольных делений в атрихобластах (Berger et al. ., 1998). С другой стороны, в каждой эпидермальной клетке мутантов glabra2 ( gl2 ) развиваются волоски, но в атрихобластах не происходит продольных делений [если в каждой клетке развивается волос, тогда где атрихобласты?], что указывает на то, что спецификация клеточной судьбы и продольные деления могут быть разъединены. Вместе эти данные указывают на то, что подмножество генов, регулирующих спецификацию клеток, также регулируют плоскость клеточного деления в эпидермисе.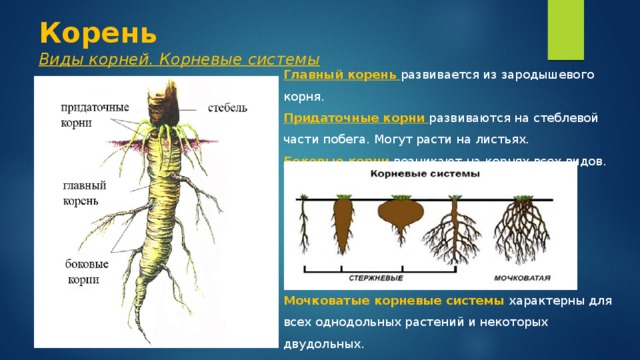
Расширение корневых клеток
Поскольку во время развития растений движение клеток отсутствует, расширение клеток является одним из ключевых параметров, определяющих окончательную форму органов растений. Насколько далеко расширяется клетка и в каком направлении находятся два основных фактора, определяющих окончательную форму клетки. Корень Arabidopsis представляет собой идеальную систему для анализа роста клеток. В частности, можно легко идентифицировать мутации, влияющие на степень или ориентацию клеточной экспансии. Различные типы клеточной экспансии происходят в разных областях корня. Для создания клеточных файлов исходные клетки и их непосредственное потомство проходят непрерывный процесс расширения и деления. Сначала расширение этих делящихся клеток корня происходит относительно медленно и неполярно, так что их размер остается довольно постоянным (4). В это время продолжающиеся клеточные деления в меристематической зоне смещают эти клетки вверх по клеточному ряду.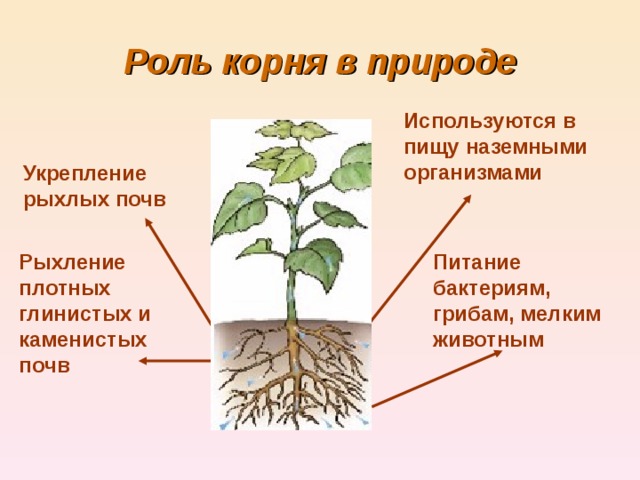 Затем происходит переход к анизотропному расширению, которое в сочетании с клеточным делением приводит к образованию клеток с более длинными радиальными размерами, чем продольные. Это особенно заметно в клетках эпидермиса (4). На этом этапе устанавливается окончательный радиус корня. Второй переход к высокополяризованному продольному расширению происходит в середине зоны растяжения. К этому времени деление клеток почти прекратилось. Это расширение происходит быстро и сильно ориентировано по продольной оси.
Затем происходит переход к анизотропному расширению, которое в сочетании с клеточным делением приводит к образованию клеток с более длинными радиальными размерами, чем продольные. Это особенно заметно в клетках эпидермиса (4). На этом этапе устанавливается окончательный радиус корня. Второй переход к высокополяризованному продольному расширению происходит в середине зоны растяжения. К этому времени деление клеток почти прекратилось. Это расширение происходит быстро и сильно ориентировано по продольной оси.
Открыть в отдельном окне
Расширение ячейки в корне. В меристематической зоне клетки обычно изодиаметричны. Когда они пересекают зону удлинения, радиальное расширение заканчивается и происходит быстрое продольное расширение.
Все данные свидетельствуют о том, что клеточная стенка растений регулирует как степень, так и ориентацию клеточной экспансии. Считается, что основной несущей молекулой в клеточной стенке являются микрофибриллы целлюлозы, которые расположены как кольца вокруг бочки. Таким образом, неудивительно, что многие из генов, идентифицированных в мутантных скринингах на измененную экспансию клеток корня, по-видимому, играют роль в синтезе целлюлозы. Например, rsw1 представляет собой чувствительную к температуре мутацию, которая при рестриктивной температуре характеризуется набуханием клеток, уменьшением количества кристаллической целлюлозы и отсутствием розеток, из которых продуцируется целлюлоза (Arioli et al., 1998). Ген RSW1 кодирует дальнего родственника каталитической субъединицы бактериальных целлюлозсинтаз с предсказанными восемью трансмембранными доменами (Arioli et al., 1998).
Таким образом, неудивительно, что многие из генов, идентифицированных в мутантных скринингах на измененную экспансию клеток корня, по-видимому, играют роль в синтезе целлюлозы. Например, rsw1 представляет собой чувствительную к температуре мутацию, которая при рестриктивной температуре характеризуется набуханием клеток, уменьшением количества кристаллической целлюлозы и отсутствием розеток, из которых продуцируется целлюлоза (Arioli et al., 1998). Ген RSW1 кодирует дальнего родственника каталитической субъединицы бактериальных целлюлозсинтаз с предсказанными восемью трансмембранными доменами (Arioli et al., 1998).
Скрининг аномальной экспансии клеток корня также выявил несколько мутаций, которые иным образом влияют на форму клеток корня. Для класса мутантов CORE (Conditional Root Expansion) фенотип клеточной экспансии зависит от скорости роста — в условиях, которые снижают скорость роста (более низкий уровень сахарозы или пониженная температура), корневые клетки напоминают клетки дикого типа (Hauser et al.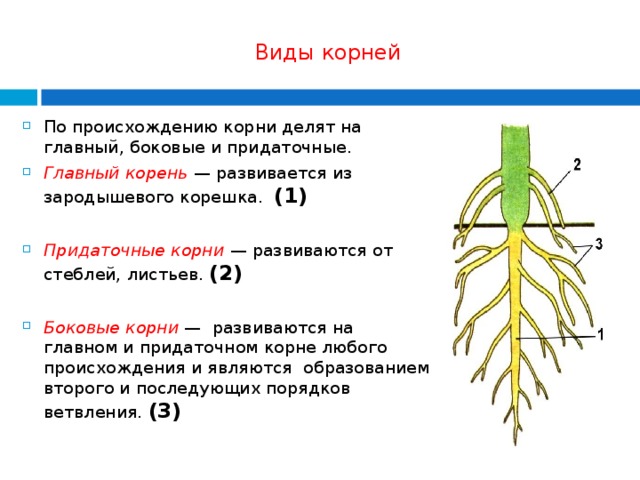 , 19).95). У одного из мутантов CORE, cobra ( cob ), первичный дефект, по-видимому, заключается в регуляции ориентации экспансии. Напротив, мутация lion’s tail вызывает уменьшение удлинения с небольшим увеличением радиального расширения, так что объем клеток значительно меньше, чем у клеток дикого типа. Другие четыре мутации CORE, quill , cudgel , pom-pom1 и pom-pom2 , имеют объемы клеток, которые больше, чем у дикого типа, что согласуется со структурной ролью или ролью в регулировании степени расширения (Hauser et al., 1995).
, 19).95). У одного из мутантов CORE, cobra ( cob ), первичный дефект, по-видимому, заключается в регуляции ориентации экспансии. Напротив, мутация lion’s tail вызывает уменьшение удлинения с небольшим увеличением радиального расширения, так что объем клеток значительно меньше, чем у клеток дикого типа. Другие четыре мутации CORE, quill , cudgel , pom-pom1 и pom-pom2 , имеют объемы клеток, которые больше, чем у дикого типа, что согласуется со структурной ролью или ролью в регулировании степени расширения (Hauser et al., 1995).
Два исходных мутанта CORE были обнаружены в ходе других генетических исследований. Мутация procuste была идентифицирована при скрининге аномального удлинения гипокотилей и является аллельной quill (Desnos et al., 1996). PROCUSTE/QUILL кодирует другую каталитическую субъединицу ферментативного комплекса синтеза целлюлозы (Fagard et al., 2000). Мутация korrigan была выделена при скрининге инсерционных мутантов на короткие и радиально расширенные клетки гипокотиля и является аллельной львиный хвост . KORRIGAN кодирует член семейства эндо-1,4-b-D-глюканаз (EGase) (Nicol et al., 1998; Sato et al., 2001). Выведенная аминокислотная последовательность KOR указывает на то, что это интегральный мембранный белок с каталитическим доменом, внешним по отношению к мембране.
KORRIGAN кодирует член семейства эндо-1,4-b-D-глюканаз (EGase) (Nicol et al., 1998; Sato et al., 2001). Выведенная аминокислотная последовательность KOR указывает на то, что это интегральный мембранный белок с каталитическим доменом, внешним по отношению к мембране.
Анализ синтеза целлюлозы у кобры выявил дефицит целлюлозы в корне, что позволяет предположить, что регуляция COB ориентированного расширения клеток связана с отложением целлюлозы. Мотивы последовательности указывают на то, что С-конец COBRA процессирован и замещен гликозилфосфатидилинозитоловым (GPI) якорем (Schindelman et al., 2001). Известно, что в клетках животных эта липидная связь обеспечивает полярную локализацию белков. Антитела локализуют белок COB преимущественно на продольных сторонах клеток корня в зоне быстрого удлинения, что согласуется с его ролью в регуляции ориентации клеточной экспансии ()). Уровни экспрессии мРНК COBRA резко возрастают в клетках, попадающих в зону быстрой продольной экспансии (Schindelman et al.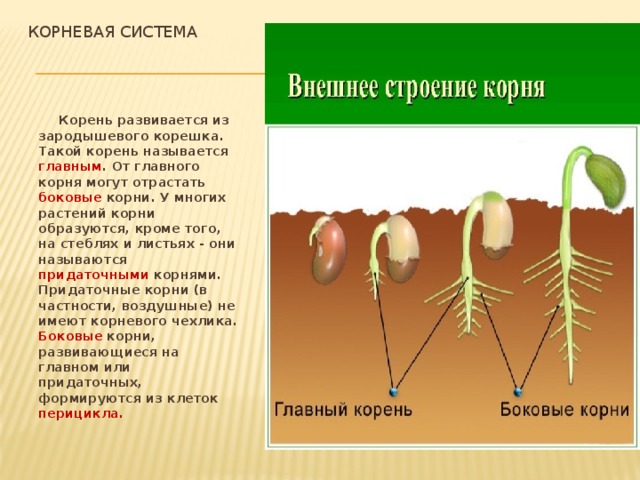 , 2001) (4).
, 2001) (4).
Открыть в отдельном окне
Окрашивание антителами белка COBRA в клетках корня. Белок, по-видимому, преимущественно локализован на продольных сторонах клеток (Schindelman et al., 2001).
Открыть в отдельном окне
Экспрессия COBRA РНК, определенная гибридизацией in situ . Экспрессия резко усиливается, когда клетки начинают фазу сильно поляризованной продольной элонгации (Schindelman et al., 2001).
Образец цитирования: Шерес Б., Бенфи П. и Долан Л. (2002) Развитие корней. Книга об арабидопсисе 1:e0101. doi:10.1199/tab.0101
elocation-id: e0101
Опубликовано: 30 сентября 2002 г.
- Ариоли Т., Пэн Л., Бетцнер А.С., Берн Дж., Виттке В., Херт В., Камиллери C., Hofte H., Plazinski J., Birch R. Молекулярный анализ биосинтеза целлюлозы у арабидопсиса. Наука. 1998;2791(1):717–20. [PubMed] [Google Scholar]
- Cnops G., Wang X., Linstead P., Van Montagu M., Van Lijsebettens M., Dolan L.
 Tornado1 и tornado2 необходимы для определения радиального и периферического узора арабидопсиса. корень. Разработка. 2000; 1271(1):3385–9.4. [PubMed] [Google Scholar]
Tornado1 и tornado2 необходимы для определения радиального и периферического узора арабидопсиса. корень. Разработка. 2000; 1271(1):3385–9.4. [PubMed] [Google Scholar] - Desnos T., Orbovic V., Bellini C., Kronenberger J., Caboche M., Traas J., Hofte H. Мутанты Procuste1 идентифицируют два различных генетических пути, контролирующих удлинение клеток гипокотиля, соответственно у проростков арабидопсиса, выращенных в темноте и на свету. Разработка. 1996;1221(1):683–93. [PubMed] [Google Scholar]
- Di Laurenzio L., Wysocka-Diller J., Malamy J.E., Pysh L., Helariutta Y., Freshour G., Hahn M.G., Feldmann K.A., Benfey P.N. Ген SCARECROW регулирует асимметричную клетку деление, необходимое для создания радиальной организации корня арабидопсиса. Клетка. 1996;861(1):423–33. [PubMed] [Google Scholar]
- Долан Л., Джанмаат К., Виллемсен В., Линстед П., Поэтиг С., Робертс К., Шерес Б. Клеточная организация корня Arabidopsis thaliana. Разработка. 1993;1191(1):71–84. [PubMed] [Google Scholar]
- Fagard M.
 , Desnos T., Desprez T., Goubet F., Refregier G., Mouille G., McCann M., Rayon C., Vernhettes S., Hofte H. PROCUSTE1 кодирует Синтаза целлюлозы, необходимая для нормального удлинения клеток, особенно в корнях и выросших в темноте гипокотилях арабидопсиса. Растительная клетка. 2000;121(1):2409–2424. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Desnos T., Desprez T., Goubet F., Refregier G., Mouille G., McCann M., Rayon C., Vernhettes S., Hofte H. PROCUSTE1 кодирует Синтаза целлюлозы, необходимая для нормального удлинения клеток, особенно в корнях и выросших в темноте гипокотилях арабидопсиса. Растительная клетка. 2000;121(1):2409–2424. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Хаузер М. Т., Мориками А., Бенфей П. Н. Мутанты арабидопсиса с условной корневой экспансией. Разработка. 1995;1211(1):1237–52. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Helariutta Y., Fukaki H., Wysocka-Diller J., Nakajima K., Jung J., Sena G., Hauser MT., Benfey P.N. Ген контролирует формирование радиального паттерна корня арабидопсиса посредством радиальной передачи сигналов. Клетка. 2000;1011(1):555–567. [PubMed] [Академия Google]
- Иноуэ Т., Хигучи М., Хашимото Ю., Секи М., Кобаяши М., Като Т., Табата С., Шинозаки К., Какимото Т. Идентификация CRE1 как рецептора цитокинина из арабидопсиса. Природа. 2001;4091(1):1060–3.
 [PubMed] [Google Scholar]
[PubMed] [Google Scholar] - Махонен А. П., Бонке М., Кауппинен Л., Рииконен М., Бенфей П. Н., Хелариутта Ю. Новая двухкомпонентная гибридная молекула регулирует сосудистый морфогенез корня арабидопсиса. Гены Дев. 2000;141(1):2938–43. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Накадзима К., Сена Г., Нави Т., Бенфей П. Н. Межклеточное перемещение предполагаемого фактора транскрипции SHR при формировании корневого паттерна. Природа. 2001;4131(1):307–11. [PubMed] [Google Scholar]
- Nicol F., His I., Jauneau A., Vernhettes S., Canut H., Hofte H. Предполагаемая эндо-1,4-бета-D-глюканаза, связанная с плазматической мембраной, представляет собой необходим для нормальной сборки стенок и удлинения клеток у арабидопсиса. Эмбо Дж. 1998;171(1):5563–76. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Сато С., Като Т., Какегава К., Исии Т., Лю Ю. Г., Авано Т., Такабэ К., Нишияма Ю., Куга С., Накамура Y. Роль предполагаемой связанной с мембраной эндо-1,4-бета-глюканазы KORRIGAN в удлинении клеток и синтезе целлюлозы у Arabidopsis thaliana.
 Физиология клеток растений. 2001;421(1):251–63. [PubMed] [Академия Google]
Физиология клеток растений. 2001;421(1):251–63. [PubMed] [Академия Google] - Scheres B., Di Laurenzio L., Willemsen V., Hauser M-T., Janmaat K., Weisbeek P., Benfey P.N. Мутации, влияющие на радиальную организацию корня Arabidopsis, обнаруживают специфические дефекты по всей оси зародыша. Разработка. 1995;1211(1):53–62. [Google Scholar]
- Schindelman G., Morikami A., Jung J., Baskin T.I., Carpita N.C., Derbyshire P., McCann M.C., Benfey P.N. Расширение клеток арабидопсиса. Гены Дев. 2001;151(1):1115–27. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Высоцкая-Диллер Дж. В., Хелариутта Й., Фукаки Х., Малами Дж. Э., Бенфей П. Н. Молекулярный анализ функции ПУЧЕЛА выявляет радиальный механизм формирования паттерна, общий для корня и побега. Разработка. 2000;1271(1):595–603. [PubMed] [Google Scholar]
- Alvarez-Buylla E.R., Liljegren S.J., Pelaz S., Gold S.E., Burgeff C., Ditta G.S., Vergara-Silva F., Yanofsky M.F. Эволюция гена MADS-box за пределами цветков: экспрессия в пыльце , эндосперм, замыкающие клетки, корни и трихомы.
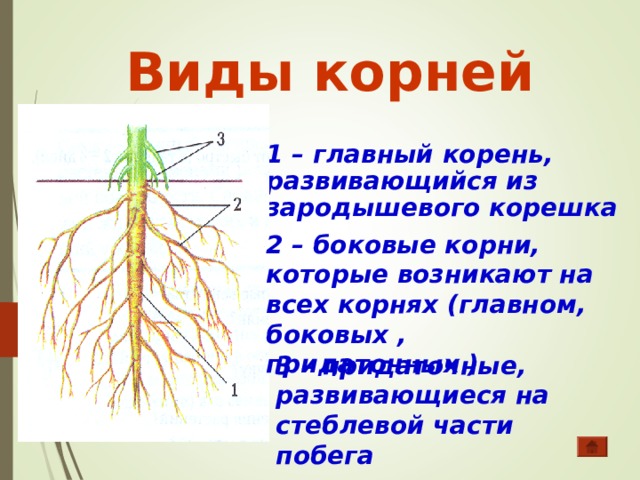 Плант Дж. 2000;241(4)(1):457–66. [PubMed] [Академия Google]
Плант Дж. 2000;241(4)(1):457–66. [PubMed] [Академия Google] - Baumberger N., Ringli C., Keller B. Химерный богатый лейцином повтор/экстенсиновый белок клеточной стенки LRX1 необходим для морфогенеза корневых волосков у Arabidopsis thaliana. Гены Дев. 2001;151(1):1128–39. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beemster GTS, Baskin TI. Анализ деления и удлинения клеток, лежащих в основе ускоренного развития роста корней у Arabidopsis thaliana . Завод Физиол. 1998;1161(1):1515–1526. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Berger F., Hung C.Y., Dolan L., Schiefelbein J. Контроль деления клеток в корневом эпидермисе Arabidopsis thaliana. Дев биол. 1998;1941(1):235–45. [PubMed] [Google Scholar]
- Berger F., Haseloff J., Schiefelbein J., Dolan L. Позиционная информация в корневом эпидермисе определяется во время эмбриогенеза и действует в доменах со строгими границами. Карр Биол. 1998;81(1):421–30. [PubMed] [Google Scholar]
- Берлет Т.
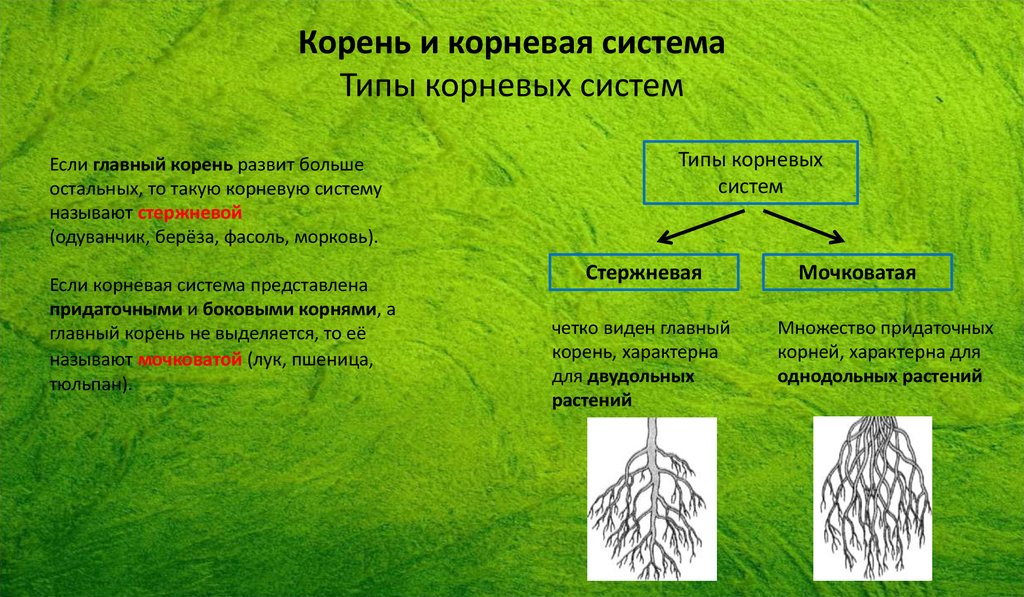 , Юргенс Г. Роль гена monopteros в организации базальной области тела Эмбрион арабидопсиса . Разработка. 1993;1181(1):575–587. [Google Scholar]
, Юргенс Г. Роль гена monopteros в организации базальной области тела Эмбрион арабидопсиса . Разработка. 1993;1181(1):575–587. [Google Scholar] - Boerjan W., Cervera M. T., Delarue M., Beeckman T., Dewitte W., Bellini C., Caboche M., Van Onckelen H., Van Montagu M., Inze D. Superroot, рецессивный мутация у арабидопсиса вызывает перепроизводство ауксина. Растительная клетка. 1995; 71(9)(1):1405–19. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Casimiro I., Marchant A., Bhalerao R.P., Beeckman T., Dhooge S., Swarup R., Graham N., Inze D., Sandberg G., Casero PJ, Bennett M. Транспорт ауксина способствует инициации боковых корней арабидопсиса. Растительная клетка. 2001; 131(4)(1):843–52. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Целенца Дж. Л., младший, Гризафи П. Л., Финк Г. Р. Путь образования боковых корней у Arabidopsis thaliana. Гены Дев. 1995; 91(17)(1):2131–42. [PubMed] [Google Scholar]
- Clowes F. A. L. Промеристем и минимальный строительный центр на низовом уровне.
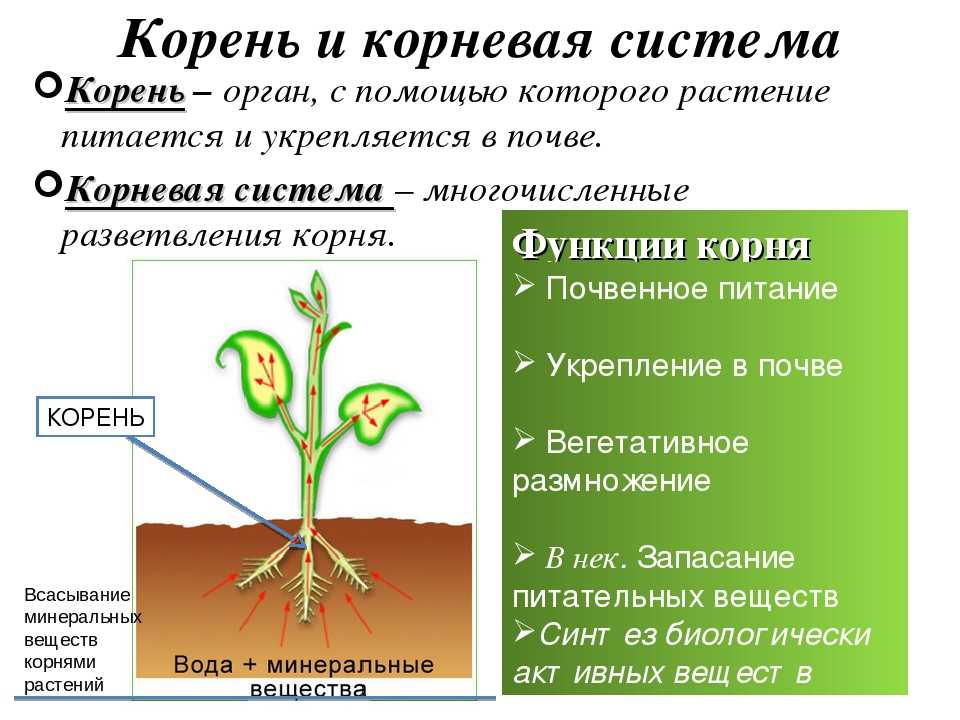 Новый Фитол. 1954; 521 (1): 48–57. [Google Scholar]
Новый Фитол. 1954; 521 (1): 48–57. [Google Scholar] - Долан Л., Дакетт С. М., Грирсон С., Линстед П., Шнайдер К., Лоусон Э., Дин С., Поэтиг Р. С., Робертс К. Клональные взаимоотношения и формирование клеточного паттерна в корневом эпидермисе Арабидопсис . Разработка . 1994;1201(1):2465–2475. [Google Scholar]
- Долан Л., Джанмаат К., Виллемсен В., Линстед П., Поэтиг С., Робертс Р., Шерес Б. Клеточная организация корня Arabidopsis thaliana . Разработка. 1993;1191(1):71–84. [PubMed] [Google Scholar]
- Favery B., Ryan E., Foreman J., Linstead P., Boudonck K., Steer M., Shaw P., Dolan L. KOJAK кодирует белок, подобный синтазе целлюлозы. необходим для морфогенеза клеток корневых волосков у арабидопсиса. Гены Дев. 2001;151(1):79–89. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Friml J., Benkova E., Blilou I., Wisniewska J., Hamann T., Ljung K., Woody S., Sandberg G., Scheres B., JFCrgens G., Palme K. AtPIN4 опосредует управляемые стоком градиенты ауксина и формирование корневого паттерна у Arabidopsis .
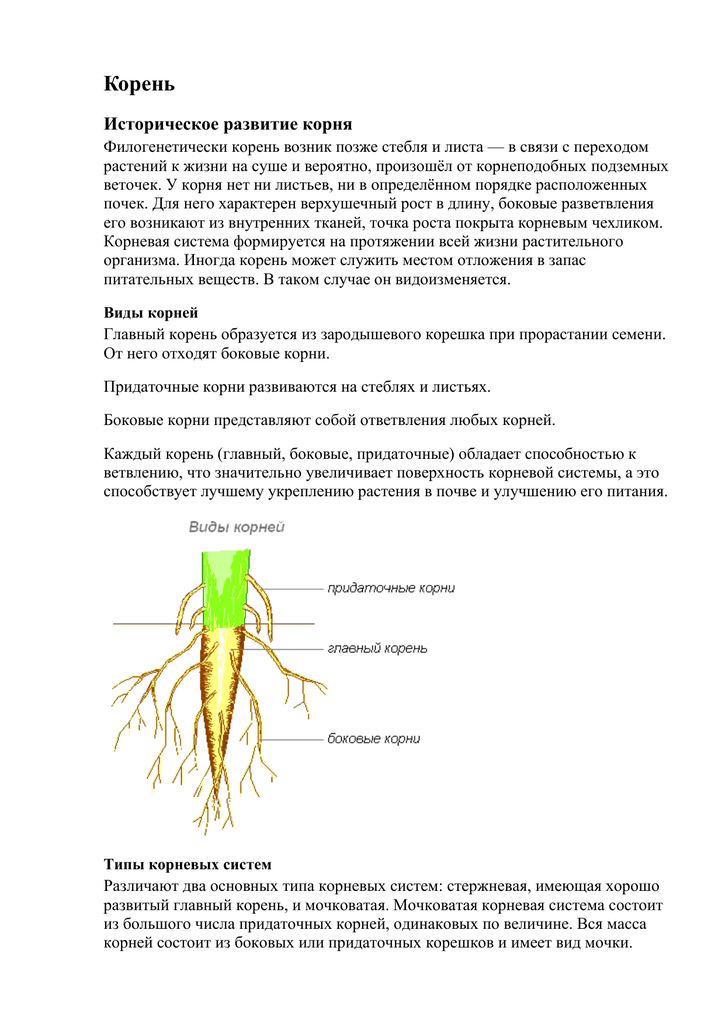 Клетка. 2002. в печати. [PubMed]
Клетка. 2002. в печати. [PubMed] - Fujie M., Kuroiwa H., Suzuki T., Kawano S., Kuroiwa T. Синтез ДНК органелл в покоящемся центре Arabidopsis thaliana (Col.). жур. Опыт Ботаника. 1993;441(1):69–693. [Google Scholar]
- Фукаки Х., Тамеда С., Масуда Х., Тасака М. Образование боковых корней блокируется мутацией усиления функции в гене SOLITARY-ROOT/IAA14 арабидопсиса. Плант Дж. 2002; 291 (1): 153–68. [PubMed] [Google Scholar]
- Голуэй М. Э., Масуччи Дж. Д., Ллойд А. М., Уолбот В., Дэвис Р. В., Шифельбейн Дж. В. Ген TTG необходим для определения судьбы эпидермальных клеток и формирования клеточного паттерна в корне арабидопсиса. Дев биол. 1994;1661(1):740–54. [PubMed] [Академия Google]
- Hamann T., Mayer U., Jurgens G. Нечувствительная к ауксину мутация bodenlos влияет на формирование первичного корня и формирование апикально-базального паттерна у зародышей Arabidopsis. Разработка. 1999;1261(7)(1):1387–95. [PubMed] [Google Scholar]
- Hardtke C.
 S., Berleth T. Ген Arabidopsis MONOPTEROS кодирует фактор транскрипции, опосредующий формирование оси эмбриона и развитие сосудов. EMBO J. 1998;171(1):1405–1411. [Бесплатная статья PMC] [PubMed] [Google Scholar]
S., Berleth T. Ген Arabidopsis MONOPTEROS кодирует фактор транскрипции, опосредующий формирование оси эмбриона и развитие сосудов. EMBO J. 1998;171(1):1405–1411. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Hobbie L., McGovern M., Hurwitz L.R., Pierro A., Liu N.Y., Bandyopadhyay A., Estelle M. Мутанты axr6 Arabidopsis thaliana определяют ген, участвующий в реакции на ауксин и раннем развитии. Разработка. 2000;1271(1)(1):23–32. [PubMed] [Google Scholar]
- Киднер К., Сундаресан В., Робертс К., Долан Л. Клональный анализ корня арабидопсиса подтверждает, что судьбу клетки определяет положение, а не происхождение. Планта. 2000;2111(1):191–199. [PubMed] [Google Scholar]
- Ли М. М., Шифельбейн Дж. WEREWOLF, белок, родственный MYB у арабидопсиса, является позиционно-зависимым регулятором формирования паттерна эпидермальных клеток. Сотовый . 1999;991(1):473–483. [PubMed] [Google Scholar]
- Leyser O., Lincoln C. A., Timpte C.
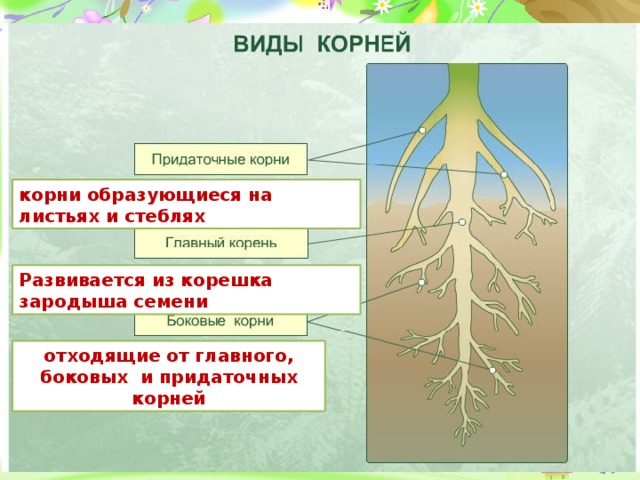 , Lammer D., Turner J., Estelle M. Arabidopsis ген устойчивости к ауксину AXR1 кодирует белок, связанный с убиквитин-активирующим ферментом Е1. Природа. 1993;3641(1):161–164. [PubMed] [Google Scholar]
, Lammer D., Turner J., Estelle M. Arabidopsis ген устойчивости к ауксину AXR1 кодирует белок, связанный с убиквитин-активирующим ферментом Е1. Природа. 1993;3641(1):161–164. [PubMed] [Google Scholar] - Малами Дж. Э., Бенфей П. Н. Организация и дифференцировка клеток в боковых корнях Arabidopsis thaliana. Разработка. 1997;1241(1)(1):33–44. [PubMed] [Академия Google]
- Masucci J.D., Rerie W.G., Foreman D.R., Zhang M., Galway M.E., Marks MD, Schiefelbein J.W. Ген гомеобокса GLABRA2 необходим для позиционно-зависимой дифференцировки клеток в корневом эпидермисе Arabidopsis thaliana . Разработка. 1996;1221(1):1253–60. [PubMed] [Google Scholar]
- Mayer K. F., Schoof H., Haecker A., Lenhard M., Jurgens G., Laux T. Роль WUSCHEL в регуляции судьбы стволовых клеток в меристеме побегов арабидопсиса. Клетка. 1998;951(6)(1):805–15. [PubMed] [Академия Google]
- Уэлле Ф., Овервурде П. Дж., Теологис А. IAA17/AXR3. Биохимический взгляд на ауксиновый мутантный фенотип.
 Растительная клетка. 2001;131(4)(1):829–42. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Растительная клетка. 2001;131(4)(1):829–42. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Паркер Дж. С., Кавелл А. С., Долан Л., Робертс К., Грирсон К. С. Генетические взаимодействия во время морфогенеза корневых волосков у арабидопсиса. Растительная клетка. 2000; 121(1):1961–74. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ригас С., Дебросес Г., Харалампидис К., Висенте-Агулло Ф., Фельдманн К., Грабов А., Долан Л., Хацопулос П. TRh2 кодирует переносчик калия, необходимый для роста кончиков корневых волосков арабидопсиса. Растительная клетка. 2001;131(1):139–51. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Роуз Д., Маккей П., Штирнберг П., Эстель М., Лейзер О. Изменения ауксинового ответа в результате мутаций в гене AUX/IAA. Наука. 1998;2791(5355)(1):1371–3. [PubMed] [Google Scholar]
- Рюггер М., Дьюи Э., Грей В. М., Хобби Л., Тернер Дж., Эстель М. Белок TIR1 Arabidopsis функционирует в ответ на ауксин и связан с SKP2 человека и дрожжей.
 гр1п. Гены Дев. 1998;121(1):198–207. [Бесплатная статья PMC] [PubMed] [Google Scholar]
гр1п. Гены Дев. 1998;121(1):198–207. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Sabatini S., Beis D., Wolkenfelt H., Murfett J., Guilfoyle T., Malamy J., Benfey P., Leyser O., Bechtold N., Вайсбек П., Шерес Б. Ауксин-зависимый дистальный организатор паттерна и полярности в корне арабидопсиса . Клетка. 1999;991(1):463–472. [PubMed] [Google Scholar]
- Scheres B., Wolkenfelt H., Willemsenm V., Terlouw M.P., Lawson E., Dean C., Weisbeek P. Эмбриональное происхождение первичного корня и корневой меристемы арабидопсиса. Разработка. 1994;1201(1):2475–2487. [Google Scholar]
- Scheres B., DiLaurenzio L., Willemsen V., Hauser M-T., Janmaat K., Weisbeek P., Benfey P. Мутации, влияющие на радиальную организацию корня Arabidopsis , обнаруживают специфические дефекты на протяжении эмбрионального развития. ось. Разработка. 1995;1211(1):53–62. [Google Scholar]
- Schiefelbein J. W., Somerville C. Генетический контроль развития корневых волосков у Arabidopsis thaliana .
 Завод. Клетка. 1990;21(1):235–243. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Завод. Клетка. 1990;21(1):235–243. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Schneider K., Mathur J., Boudonck K., Wells B., Dolan L., Roberts K. Ген ROOT HAIRLESS 1 кодирует ядерный белок, необходимый для инициации корневых волосков у Arabidopsis. Гены Дев. 1998;121(1):2013–21. [Статья бесплатно PMC] [PubMed] [Google Scholar]
- Шнайдер К., Уэллс Б., Долан Л., Робертс К. Структурный и генетический анализ дифференцировки клеток эпидермиса в первичных корнях арабидопсиса . Разработка. 1997; 1241(1):1789–1798. [PubMed] [Академия Google]
- Steinmann T., Geldner N., Grebe M., Mangold S., Jackson C.L., Paris S., Galweiler L., Palme K., Jurgens G. Координированная полярная локализация переносчика оттока ауксина PIN1 с помощью GNOM ARF GEF. Наука. 1999;2861(5438)(1):316–8. [PubMed] [Google Scholar]
- Тимпте С., Уилсон А.К., Эстель М. Мутация axr2-1 у Arabidopsis thaliana представляет собой мутацию с приобретением функции, которая нарушает ранний этап реакции ауксина. Генетика. 1994;1381(4)(1):1239–49. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Van den Berg C., Willemsen V., Hendriks C., Weisbeek P., Scheres B. Ближний контроль дифференцировки клеток в корневой меристеме Arabidopsis . Природа. 1997;3901(1):287–289. [PubMed] [Google Scholar]
- Van den Berg C., Willemsen V., Hage W., Weisbeek P., Scheres B. Судьба клеток в корне Arabidopsis определяется направленной передачей сигналов. Природа. 1995;3781(1):62–65. [PubMed] [Google Scholar]
- Вада Т., Татибана Т., Шимура Ю., Окада К. Дифференцировка эпидермальных клеток арабидопсиса, определяемая гомологом Myb, CPC. Наука. 1997;2771(1):1113–1116. [PubMed] [Google Scholar]
- Wang H., Lockwood S. K., Hoeltzel M. F., Schiefelbein J. W. Ген ROOT HAIR DEFECTIVE3 кодирует эволюционно консервативный белок с GTP-связывающими мотивами и необходим для регулируемого увеличения клеток у арабидопсиса. Гены Дев. 1997;111(1):799–811. [PubMed] [Google Scholar]
- Willemsen V. , Wolkenfelt H., de Vrieze G., Weisbeek P., Scheres B. Ген HOBBIT необходим для образования первичной меристемы корня в Эмбрион арабидопсиса . Разработка. 1998;1251(1):521–531. [PubMed] [Google Scholar]
- Ваймер С. Л., Бибикова Т. Н., Гилрой С. Распределение свободного кальция в цитоплазме при развитии корневых волосков Arabidopsis thaliana. Плант Дж. 1997;121(1):427–39. [PubMed] [Google Scholar]
- Xie Q., Frugis G., Colgan D., Chua N.H. Arabidopsis NAC1 передает сигнал ауксина ниже по течению от TIR1, чтобы способствовать развитию боковых корней. Гены Дев. 2000;141(23)(1):3024–36. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Статьи из книги Arabidopsis предоставлены здесь Американским обществом биологов растений
корней | Информационная система для кормов
Корни служат для прикрепления растений к почве, как органы хранения, а также для поглощения и отвода воды и основных минералов из почвы. Эти процессы варьируются в зависимости от воды и минералов, имеющихся в растении и почве. Корни травянистых растений глубоко погружаются в почву, прежде чем большая часть их выйдет из-под поверхности почвы. Это дает основу для роста. Например, до появления побега развивается 2-3 дюйма корня. Корень может достигать 6 дюймов в глубину в течение 2 недель.
У злаков существуют две корневые системы: 1) первичные корни, развивающиеся из зародыша при прорастании семян, и 2) придаточные корни, возникающие из узлов кроны и боковых стеблей.
Первичные или семенные (семенные) корни обычно не живут дольше первого сезона после посадки. Вторичные или придаточные корни начинают формироваться вскоре после появления первого листа. Придаточные корни могут функционировать на протяжении всей жизни побега, однако продолжительность жизни корней частично определяется управлением дефолиацией, а также производством семян.
Как и у теплолюбивых трав, у теплолюбивых трав рост корней возобновляется весной с наступлением благоприятных для них климатических условий. При разумном управлении виды теплого сезона пускают корни на несколько большую глубину в профиле почвы, чем виды холодного сезона, из-за большей устойчивости к жаре и засухе.
Рост корня происходит за счет расширения недавно развитых клеток в результате деления клеток меристемы, расположенных сразу за корневым чехликом. Созревание и дифференцировка удлиненных клеток приводят к развитию специализированных тканей для транспортировки воды и питательных веществ к другим частям растения.
Вода поглощается корневыми волосками и отводится в энтодермальные клетки по осмотическому градиенту в проводящие клетки ксилемы. Поглощение питательных веществ зависит от энергии дыхания. Травы, растущие на сильно уплотненных или заболоченных почвах, могут иметь недостаточное количество кислорода для дыхания, что приводит к ограничению транспорта внутри корня.
Виды травы могут быть обозначены как многолетние укореняющиеся или однолетние укореняющиеся, имея в виду, как часто заменяется корневая система многолетних растений. Корни мятлика Кентукки сохраняются более года (многолетнее укоренение), в то время как полевица, бермуда, райграс пастбищный и другие образуют новые корни каждый год (ежегодное укоренение).
Ссылка: Освальт, Д.Л., А.Р. Бертран и М. Р. Тил. Влияние азотных удобрений и обрезки на корни травы. Почвовед. соц. амер. проц. 23: № 3, 1959.
Последствия для управления
Когда побеги травы, способные дать цветоносную стеблю, дефолиируют во время раннего соединения или позже таким образом, что удаляется вся надземная меристема, рост корней серьезно затрудняется. Продолжительность этого эффекта зависит не только от последствий дефолиации цветущих побегов, но и от побегов, не индуцируемых к цветению. «Материнская» корневая система побега стареет вместе с развитием семенной головки. Таким образом, отростки должны развить новую корневую систему. Своевременный выпас или обрезка пастбищ для предотвращения развития семенных головок вызывает рост отростков и последующее развитие корней до наступления засушливых почвенных условий. Вышеупомянутая агротехника снижает выраженность летнего покоя костра гладкого.
Суть хорошего управления пастбищами заключается в сохранении меристем до тех пор, пока корни не укоренятся. Вовлеченные меристемы – это апикальная меристема (находится в зачатке побега) и интеркалярная меристема (находится в воротнике в основании листовой пластинки). Когда побеги травы вступают в переходную стадию, когда удлинение междоузлий стебля поднимает точку роста побега (зачаток) до уязвимой высоты, побеги травы могут подвергаться дефолиации тремя различными способами, каждый из которых по-разному влияет на корни.
1) Интенсивная дефолиация уничтожает «приподнятый» зачаток побегов на большом проценте цветущих побегов. Этот эффект обычно называют «обезглавливанием». При декапитации потенциальная головка семени и критическая меристема интеркалярной пластинки разрушаются. Скорость отрастания новых побегов, возникающих из адвентивных почек в ткани кроны, будет варьироваться в зависимости от степени развития зачатков новых побегов и уровня доступных запасов пищи. Корни будут лишены энергии фотосинтеза до тех пор, пока новые побеги не дадут больше энергии, чем используется для вегетативного обновления.
2) Менее интенсивная дефолиация сохранит точку роста побегов, однако из-за удлинения влагалища более развитые листья могут быть отделены ниже воротниковой зоны, т. Таким образом, по мере того, как стебель продолжает развиваться, может остаться только флаговый лист. Этот эффект упоминается как «обнажение», поскольку стебель остается практически голым. Оголенные побеги обладают минимальной фотосинтетической способностью. Запасы энергии также минимальны, поскольку для раннего вегетативного роста требуются практически все углеводы, произведенные в результате фотосинтеза. Имеющиеся источники энергии будут использоваться для развития стеблей и семенных головок. Корни будут лишены источника энергии до тех пор, пока новые побеги не произведут избыток сахара.
3) Дефолиация в начале удлинения междоузлий, обычно называемая в управлении выпасом скота «ранним укусом», создает минимальный риск быстрого восстановления при условии сохранения точки роста побега и достаточного количества ткани листовой пластинки в составе стерни. При соблюдении вышеуказанных предосторожностей восстановительный рост в значительной степени не зависит от запасов пищи в тканях кроны из-за быстрого возобновления развития листовых пластинок.
Меристематическая точка роста вместе с интеркалярной меристемой, расположенной в воротниковой зоне развивающихся лопастей, представляют собой активные в настоящее время механизмы отрастания. При сохранении эти системы вызывают конкурентное возобновление роста с минимальным нарушением развития корней.
Практическое применение
Подкормочные пастбища
В конце пятидесятых годов один австралийский зоотехник подвел итоги многолетнего исследования пастбищ с участием трех животноводческих ферм. Вес телят при отъеме на одном ранчо неизменно превышал вес телят с соседних ранчо на 70–100 фунтов на голову. Различия в весе при отъеме нельзя было объяснить изменчивостью, связанной с породами, датами отела, типами почвы, видами корма и т. Д. Однако владелец ранчо с более тяжелыми телятами практиковал «удобрение» своих пастбищ. Эта практика была единственной выявленной переменной, которая могла бы объяснить различия в весе при отъеме. Обрезка была объяснена как стрижка пастбища роторной косилкой в начале колошения, чтобы предотвратить дальнейшее развитие цветоносов. Если цветущие стебли не съедать до появления семенных головок, они становятся деревянистыми и неприятными на вкус. При своевременной стрижке, пока трава еще вкусная, скот охотно съест остатки.
Как докормка связана с изменением веса при отъеме? Физиологи трав показали, что нарушение развития семенных головок вызывает образование новых побегов (отходов) из базальных почек в ткани кроны. Своевременная засыпка пастбищ приводит к более раннему получению откорма. Пышные последствия обеспечили рацион более высокого качества, чем на соседних ранчо.
Таким образом, домашний скот получил двойную выгоду от вышеуказанной практики; они потребляли цветущие стебли, которые в противном случае были бы отвергнуты, и наслаждались более качественным рационом в середине лета, состоящим из пышных побегов.
Развитие корней молодой кукурузы (Университет Пердью)
Обновлено в апреле 2020 г.
URL: http://www.kingcorn.org/news/timeless/Roots.html
R.L. ун-т
West Lafayette, IN 47907-2054
Адрес электронной почты: rnielsen at purdue.edu
Twitter: @PurdueCornGuy
Успешные всходы (быстрые и равномерные), хотя и важны, не гарантируют успешного образования насаждений кукурузы. Следующей важной фазой в жизни молодых растений кукурузы является начальное формирование мощной узловой корневой системы. Успешное формирование насаждения в значительной степени зависит от начального развития узловых корней примерно от V2 (два листа с видимыми воротничками листьев) до V6.
Кукуруза представляет собой траву и имеет корневую систему мочковатого типа, в отличие от соевых бобов или люцерны, которые имеют стержневую корневую систему. Задержка роста или ограничение узловых корней во время их начального развития (например, из-за чрезмерно сухой почвы, чрезмерно влажной почвы, очень холодной почвы, повреждения насекомыми, гербицидами, уплотнения боковых стенок, уплотнения при обработке почвы) может легко остановить развитие всего растения. На самом деле, когда вы пытаетесь диагностировать причину низкорослости кукурузы в начале сезона, первое место, с которого нужно начать поиск виновника, находится под землей.
Чтобы лучше понять развитие укоренения и проблемы, связанные с ограничением корней, важно понимать, что развитие корней у кукурузы происходит в две фазы. Первая фаза – развитие семенной или семенной корневой системы. Вторая фаза – развитие узловой или коронковой корневой системы.
Корни Общая информация: Иногда можно услышать, что семенную корневую систему называют первичной корневой системой, а узловую корневую систему — вторичной корневой системой. Эта классификация была описана Кэнноном (1949) и, безусловно, имеет хронологический смысл. Однако с точки зрения важности для кукурузы узловая корневая система явно является первичной корневой системой.
Семенная (семенная) корневая система
Семенные (семенные) корни происходят из щиткового узла, расположенного внутри семенного зародыша. Семенные корешки состоят из корешков и боковых семенных корешков. Несмотря на то, что семенные корни технически являются узловыми корнями, они традиционно обсуждаются отдельно от узловых корней, которые развиваются позже из области макушки сеянца.
Корень корешка выходит первым, разрывая семенную оболочку возле кончика ядра (рис. 1) и ненадолго удлиняясь в этом направлении, независимо от того, направлен ли кончик ядра вверх, вниз или вбок. Боковые семенные корешки появляются позже из-за колеоптиля (рис. 2) и первоначально удлиняются к вдавленному концу ядра. Однако вскоре обе группы семенных корешков меняют направление своего удлинения и начинают удлиняться вниз под действием силы тяжести (рис. 3).
Семенная корневая система помогает поддерживать развитие проростков благодаря поглощению воды из почвы, но молодые проростки кукурузы в первую очередь зависят от энергетических запасов крахмалистого эндосперма ядра для питания, пока позже не разовьется узловая корневая система. Примерно к стадии роста V1 скорость нового роста семенной корневой системы резко замедляется, поскольку первичная узловая корневая система начинает развиваться из узлов над мезокотилем.
Несмотря на то, что семенная корневая система мало способствует сохранению растений кукурузы в течение всего сезона, раннее повреждение корешков или боковых семенных корней может остановить начальное развитие проростков и задержать появление всходов. Такое повреждение не обязательно приведет к немедленной гибели проростка, пока само ядро и мезокотиль остаются здоровыми, но может привести к задержке появления всходов или распусканию листьев под землей. По мере того, как со временем образуется все больше и больше узловых корней, повреждение семенной корневой системы будет все меньше и меньше влиять на выживаемость проростков.
Примеры повреждения семенных корней включают имбибиционное повреждение от переохлаждения (Nielsen, 2020a), повреждение после прорастания от летальных или сублетальных низких температур (Nielsen, 2020b) и «солевое» повреждение из-за чрезмерного количества стартового удобрения, размещенного слишком близко к ядро. Симптомы такого повреждения корня включают замедленное удлинение корня, обесцвечивание ткани коричневого цвета, обильное ветвление корня и полное отмирание ткани корня. Если корень зародыша сильно поврежден во время его выхода из ядра, весь корень зародыша может погибнуть. Как только корешок удлинится примерно на полдюйма или около того, повреждение кончика корня не обязательно приведет к гибели всего корня, скорее пазушные меристемы корня могут инициировать обширное ветвление корня в ответ на повреждение апикальной меристемы.
Изображения, показанные на рис. 4-6 показывают пример отсроченного появления в поле, где “нормальные” всплывающие растения уже находились в позднем V1 – раннем V2. Корешковый корень был полностью разрушен, хотя боковые семенные корешки были целы и здоровы. Колеоптиль на этом сеянце был расщеплен по всей длине его стороны и, вероятно, привел к тому, что листья остались под землей. Расщепленный колеоптиль, вероятно, возник из-за продолжающегося естественного расширения закрытых листьев, которые в противном случае обычно вырастали бы над землей.
Изображения, показанные на рис. 7 и 8 показывают другой пример позднего появления всходов на том же поле, где другие сеянцы были от позднего V1 до раннего V2. Единственным видимым повреждением этого отсроченного прорастания был его корешковый корень, апикальная меристема которого была повреждена. Повреждения были менее серьезными, чем в предыдущем примере, поэтому сеянец не так сильно отставал в росте, и ему удалось прорасти над землей.
Узловая корневая система
Узловые корни развиваются последовательно из отдельных узлов над мезокотилем, начиная с самого нижнего узла в области молодого проростка, известного как «корона». Когда воротник первого листа впервые становится видимым, первый набор узловых корней можно определить по небольшому вздутию в самом нижнем узле. К концу V1 первая группа узловых корней заметно начала удлиняться (рис. 9).и 10). На стадии листа V2 отчетливо виден первый набор узловых корней, а второй набор узловых корней может начать удлиняться от второго узла проростка. Каждый набор или «мутовка» узловых корней начинает удлиняться от соответствующих узлов примерно в то же время, когда каждая листовая шейка выходит из настоящей мутовки сеянца.
Информация о глубине посева и глубине укоренения: Некоторые считают, что более глубокая посадка кукурузы способствует более глубокому укоренению, и наоборот. Это убеждение в основном является мифом с небольшим намеком на правду. Это, безусловно, правда, что глубина корневой системы СЕМИНА зависит от глубины посева. Однако на корневую систему NODAL, которая развивается из макушки растения, глубина посева совсем не влияет. Это связано с тем, что глубина кроны довольно постоянна независимо от глубины посева. Во время появления всходов мезокотиль удлиняется и поднимает колеоптиль и коронку к поверхности почвы. Когда колеоптиль приближается к поверхности почвы, изменения в соотношении красных и дальнекрасных длин волн света вызывают изменение поступления одного или нескольких гормонов роста от колеоптиля к ткани мезокотиля, и удлинение мезокотиля, следовательно, останавливается (Vanderhoef & Бриггс, 19 лет78). Поскольку глубина, на которой появляющиеся проростки воспринимают переход от красного к дальнему красному свету, довольно постоянна, результирующая глубина кроны (основания) колеоптиля почти одинакова (от 1/2 до 3/4 дюйма) для глубины посева. одного дюйма или больше.
Удлинение ткани стебля начинается между стадиями листа V4 и V5. Удлинение междоузлия над пятым узлом обычно приподнимает шестой узел над землей. Последующее удлинение междоузлий стебля с более высокими номерами приведет к более высокому размещению остальных узлов стебля. Наборы узловых корней, которые формируются в надземных узлах стебля, обычно называют «распорными» корнями, но они функционируют идентично тем узловым корням, которые формируются под землей. Если условия на поверхности почвы благоприятны (влажная и не слишком жаркая), опорные корни будут успешно проникать в почву, размножаться и эффективно очищать верхние слои почвы от воды и питательных веществ.
Корни Общая информация: Корневые волоски представляют собой боковые отростки корневых эпидермальных клеток, вырастают до нескольких миллиметров в длину, и их число составляет около 200 на квадратный миллиметр (Gardner et al. , 1985). Их типичная продолжительность жизни составляет всего около 2 дней при умеренных температурах и меньше при более высоких температурах (Gardner et al., 1985). Корневые волоски видны даже на корневом отростке молодого сеянца (рис. 13). В совокупности площадь поверхности, представленная корневыми волосками, очень велика и может составлять большую долю поглощения питательных веществ и влаги растением.
Корни Общая информация: Первичная меристема корня располагается вблизи кончика корня (рис. 14). Удлинение клеток позади меристемы приводит к удлинению корня.
Расколотый стебель старого растения обнажит «деревянистый» или «содержательный» треугольник ткани стебля в нижней части стебля кукурузы. Этот треугольник обычно состоит из четырех стеблевых узлов, расположенных последовательно с № 1 внизу, чьи связанные междоузлия не удлиняются (рис. 15). Первым удлиняющимся междоузлием ОБЫЧНО является тот, который находится над четвертым узлом, который удлиняется примерно на 1/4–1/2 дюйма, над которым находится пятый узел, который обычно все еще находится ниже или прямо на поверхности почвы. Следовательно, пять наборов или мутовок узловых корней обычно обнаруживаются под землей, по одному набору на каждый из подземных узлов стебля (рис. 16).
Проростки кукурузы переходят от питательной зависимости от запасов зерна к питательной зависимости от узловых корней на стадии листа V3. Повреждение или стресс первых нескольких наборов развивающихся узловых корней в период времени с V1 по V5 может серьезно остановить или задержать развитие растения кукурузы. Повреждение первых нескольких групп узловых корней вынуждает молодой сеянец продолжать зависимость от запасов ядра дольше, чем это необходимо. Если запасы ядра почти исчерпаны, дальнейшее развитие сеянцев легко останавливается, и сеянцы нередко погибают. Типичные стрессы, которые могут задержать начальное узловое развитие, включают повреждение солями удобрений, болезни рассады, повреждение гербицидами, повреждение кормления насекомыми, чрезмерно влажную или сухую почву, уплотнение почвы (обработка почвы или сеялка).
Стартовое удобрение Примечание: Успех или неудача этого переходного периода, который происходит вокруг стадии развития V3, сильно влияет на то, будет ли культура продолжать развиваться сильно и равномерно. Нередки случаи, когда месторождения разрабатываются довольно равномерно примерно до V3, в то время как они все еще полагаются на основные запасы. Однако иногда эти до сих пор однородные насаждения кукурузы «разваливаются» после стадии V3, если развитие узловых корней было нарушено «плохими» условиями выращивания, а переход от запасов ядра к поддержке узловых корней терпит неудачу или менее чем успешен. Именно на этом этапе стартовое удобрение играет роль в обеспечении успешного прохождения переходного периода. Приблизительно на V3 один или несколько узловых корней соединятся с лентой стартового удобрения, размещенной примерно на 2 дюйма в сторону и на 2 дюйма ниже семени (пресловутое размещение 2×2). Стартовое удобрение, размещенное в этом положении, имеет преимущества перед внесением семян, потому что а) его положение относительно развития узловых корней является более выгодным и б) можно использовать более высокие дозы азота и/или калия без риска повреждения семян во время выращивания. прорастание и появление.
Несколько необычный, но драматический симптом низкорослости корня называется синдромом «гибкой кукурузы» или «мозоли без корней» (Nielsen, 2019a). Эта проблема чаще всего возникает в результате пагубного воздействия чрезмерно сухой поверхности почвы во время начального удлинения узловых корней у молодых (от V2 до V4) растений кукурузы. Молодые узловые корни, которые выходят из области кроны растения, погибнут, если кончики их корней (и связанные с ними области меристемы) высохнут до успешного укоренения во влажной почве. Корона молодого растения кукурузы обычно располагается всего на 3/4 дюйма или около того ниже поверхности почвы и поэтому особенно уязвима для условий сухого верхнего слоя почвы.
Щелкните изображение, чтобы просмотреть увеличенную версию во всплывающем окне. Стрелка влево или вправо на клавиатуре будет циклически перемещаться по всплывающим изображениям.
Рис. 1. Корешковый корень и колеоптиль проростка до ВЭ.
Рис. 2. Боковые семенные корни, колеоптиль и прикорневой корень проростка до ВЭ.
Рис. 3. Семенная корневая система проростка ВЭ, но признаков узловой корневой системы пока нет.
Рис. 4. Отсроченная эмерджентность со здоровыми боковыми семенными корешками, но поврежденным корешковым корнем.
Рис. 5. Расщепленный колеоптиль; предшественник выхода из подполья.
Рис. 6. Отмерший корень проростка с отсроченным появлением всходов.
Рис. 7. Отсроченный эмерджент со здоровыми боковыми семенными корешками и поврежденным, но живым корешковым корнем.
Рис. 8. Вид вблизи поврежденной верхушки корневого корня с многочисленными придаточными корнями.
Рис. 9. Проросток кукурузы V1.
Рис. 10. Первый набор узловых корней, отходящих от самого нижнего узла коронки проростка кукурузы V1.
Рис. 11. Проростки кукурузы от позднего V1 до раннего V2.
Рис. 12. Семенные и узловые корни сеянца V2.
Рис. 13. Корневые волоски проростка кукурузы V2.
Рис. 14. Апикальная меристема корня.
Рис. 15. Удлинение междоузлий между четвертым и пятым узлами растения кукурузы.
Рис. 16. Пять идентифицируемых наборов или «завитков» узловых корней на разрезе стебля.
Похожие ссылки
Кэннон, Уильям Остин. 1949. Предварительная классификация корневых систем. Экология 30[4], 542-548.
Гарднер, Франклин П., Р. Брент Пирс и Роджер Л. Митчелл. 1985. Физиология сельскохозяйственных растений. Университет штата Айова. Пресс, Эймс, ИА.
Нильсен, Р.Л. (Боб). 2019а. Синдром бескорневой или «гибкой» кукурузы. Сеть новостей Corny, расширение Purdue. http://www.kingcorn.org/news/timeless/FloppyCorn.html. [URL по состоянию на апрель 2020 г.].
Нильсен, Р.Л. (Боб). 2019б. Визуальные индикаторы прорастания кукурузы. Сеть новостей Corny, расширение Purdue. http://www.kingcorn.org/news/timeless/GerminationEvents.html. [URL по состоянию на апрель 2020 г.].
Нильсен, Р.Л. (Боб). 2020а. Холодные почвы и риск имбибиционного холода в кукурузе. Сеть новостей Corny, расширение Purdue. http://www.kingcorn.org/news/timeless/ImbibitionalChilling.html. [URL по состоянию на апрель 2020 г.].
Нильсен, Р.Л. (Боб). 2020б. Всхожесть кукурузы. Сеть новостей Corny, расширение Purdue. http://www.kingcorn.org/news/timeless/EmergenceFailure.html. [URL по состоянию на апрель 2020 г.].
Нильсен, Р.Л. (Боб). 2020г. Процесс появления в кукурузе. Сеть новостей Corny, расширение Purdue. http://www.kingcorn.org/news/timeless/Emergence.html. [URL по состоянию на апрель 2020 г.].
Вандерхоф, Ларри Н. и Уинслоу Р. Бриггс. 1978. Ингибированное красным светом удлинение мезокотиля у проростков кукурузы. I. Гипотеза ауксина. Физика растений 61:534-537.
Carvana и Root, Inc. являются эксклюзивными партнерами по разработке первых в отрасли интегрированных решений по автострахованию для клиентов Carvana
Carvana и Root, Inc. являются эксклюзивными партнерами по разработке первых в отрасли интегрированных решений по автострахованию для клиентов Carvana
Carvana инвестирует около 126 миллионов долларов в ведущую компанию Insurtech Root, Inc.
ФЕНИКС И КОЛУМБУС, Огайо–(БИЗНЕС-ПРОВОД)– Carvana (NYSE: CVNA), ведущая платформа электронной коммерции для покупки и продажи подержанных автомобилей, и Root, Inc. (NASDAQ: ROOT), материнская компания Root Insurance Company («Root»), ведущего страхового оператора, сегодня объявила об эксклюзивном партнерстве для разработки интегрированных решений автострахования для онлайн-платформы Carvana для покупки автомобилей. В связи с партнерством Carvana инвестирует около 126 миллионов долларов в Root, Inc. при условии получения одобрения регулирующих органов.
«В лице Root мы нашли партнера, который разделяет нашу клиентоориентированность и ориентированный на технологии подход к обеспечению исключительного качества обслуживания клиентов. Мы рады продолжить успех нашего существующего партнерства, создав уникальное интегрированное решение», — сказал генеральный директор Carvana Эрни Гарсия. «Мы с нетерпением ждем возможности познакомить наших клиентов с бесшовным процессом страхования Root и верим, что это интегрированное предложение углубит и расширит наши отношения с клиентами между транзакциями».
«Carvana произвела революцию в сфере покупки подержанных автомобилей для потребителей. Это партнерство еще больше расширяет возможности Carvana за счет интеграции страхования с решениями о покупке автомобилей и финансировании», — сказал генеральный директор Root Алекс Тимм. «Это интегрированное решение предоставит Root эксклюзивный доступ к масштабируемому и растущему каналу потенциальных клиентов в момент принятия важного страхового решения о покупке автомобиля, что позволит нам удовлетворить насущные потребности клиентов и обеспечить устойчивый рост держателей полисов».
В соответствии с условиями инвестиционного соглашения, Carvana инвестирует около 126 миллионов долларов основного капитала в Root, Inc. в конвертируемые привилегированные ценные бумаги, конвертируемые по цене 9,00 долларов за акцию, примерно в 14 миллионов акций класса А в Root, Inc. или в собственность. процентная ставка примерно 5% на полностью разводненной основе на сегодняшний день. Инвестиционное соглашение также предоставляет Carvana варранты на акции класса A в Root, Inc. , которые связаны с результатами коммерческого партнерства.
Ardea Partners LP выступает в качестве финансового консультанта, а Skadden, Arps, Slate, Meagher & Flom LLP выступает в качестве юридического консультанта Root в связи с этой сделкой.
Root, Inc. Результаты за второй квартал 2021 года
Root, Inc. отдельно объявила результаты за второй квартал 2021 года и проведет телеконференцию в четверг, 12 августа 2021 года, в 8:00 по восточному времени.
О компании Carvana
Компания Carvana (NYSE: CVNA), основанная в 2012 году и базирующаяся в Фениксе, ставит перед собой задачу изменить то, как люди покупают автомобили. Удалив традиционную дилерскую инфраструктуру и заменив ее технологиями и исключительным обслуживанием клиентов, Carvana предлагает потребителям интуитивно понятную и удобную онлайн-платформу для покупки и финансирования автомобилей. Carvana.com позволяет потребителям быстро и легко покупать более 45 000 автомобилей, финансировать, обменивать или продавать свой текущий автомобиль Carvana, подписывать контракты и планировать доставку или получение на следующий день в одном из собственных автомобилей Carvana. автоматические торговые автоматы для автомобилей.
О Root, Inc.
Root, Inc. является материнской компанией Root Insurance Company. Root — технологическая компания, революционизирующая личное страхование с моделью ценообразования, основанной на честности и современном клиентском опыте. Мобильный клиентский опыт Root призван упростить страхование.
Надежная гавань для заявлений прогнозного характера Настоящий пресс-релиз и любые связанные с ним устные заявления, сделанные представителями Root или Carvana, могут содержать заявления прогнозного характера по смыслу Раздела 27A Закона о ценных бумагах от 1933 и раздел 21E Закона о фондовых биржах 1934 года. Прогнозные заявления могут относиться к стратегическим альтернативам, будущим операциям, стратегиям, планам, партнерствам, инвестициям, выкупу акций, финансовым результатам или другим событиям, и на них распространяются предположения, риски и неопределенности. Такие утверждения, как «руководство», «ожидание», «предвидение», «сильный», «верю», «намерение», «цель», «задача», «цель», «позиция», «потенциал», «будет» , «может», «будет», «должен», «может», «доставлять», «ускорять», «давать возможность», «оценивать», «проекты», «перспективы», «возможности», «расширение», « создание» или аналогичные слова, а также конкретные прогнозы будущих событий или результатов квалифицируются как прогнозные заявления. Прогнозные заявления по своему характеру подвержены целому ряду неотъемлемых рисков и неопределенностей, которые могут привести к тому, что фактические результаты будут существенно отличаться от прогнозируемых результатов. Многие из этих рисков и неопределенностей не могут контролироваться Root или Carvana и включают возможность того, что условия закрытия предлагаемой сделки могут быть не выполнены. Факторы, которые могут привести к существенному отличию наших фактических решений или результатов от предполагаемых в этих прогнозных заявлениях, можно найти в форме 10-K и форме 10-Q компании Root, поданных в Комиссию по ценным бумагам и биржам США, а также в форме 10-K компании Carvana и Форма 10-Q подается в Комиссию по ценным бумагам и биржам США соответственно. Прогнозные заявления действительны только на дату, когда заявление было сделано, и ни Root, ни Carvana не берут на себя никаких обязательств по обновлению таких прогнозных заявлений. Нет никакой гарантии, что другие факторы, которые в настоящее время не раскрываются или не ожидаются Root или Carvana, не окажут существенного неблагоприятного влияния на результаты деятельности или планы каждой компании. Инвесторов предупреждают, чтобы они не слишком полагались на какие-либо прогнозные заявления, сделанные любой компанией или от ее имени.
Посмотреть исходную версию на сайте businesswire.com: https://www.businesswire.com/news/home/20210811005876/en/
Отношения с инвесторами:
Карвана
Майк Левин
[email protected]
Корень, Inc.
Кристин Патрик
Вице-президент по связям с инвесторами
[email protected]
или
Контакты для СМИ:
Карвана
Эми О’Хара
[email protected]
Корень, Inc.
Том Кун
Директор по коммуникациям
[email protected]
Источник: Carvana
Укоренение черенков в воде
Дом Поддержите сад УчастникиМагазинВолонтерыВакансииСМИКонтакты
Укоренение черенков в воде
Выберите завод Выберите легко укореняющееся комнатное растение, например бегонию, которая стала высокой и длинноногой. Многие комнатные растения, такие как бегонии, колеусы, горошек, плющ и филодендроны, легко укореняются в воде. Другие растения, в том числе многие древесные растения, такие как гибискус и цитрусовые, плохо укореняются в воде. Они обычно гниют до укоренения. | |
| Взять черенки с растения Взять черенки с растения. Для большинства растений черенки должны быть от 4 до 6 дюймов в длину. Не делайте черенки слишком большими; они плохо укоренятся или, если укоренятся, вместо компактного станут высоким долговязым растением. | |
Срезанные стебли чуть ниже почки С помощью острого ножа (или секатора) срежьте чуть ниже места прикрепления листа к стеблю (узел). | |
Снятие нижних створок Удалите нижние листья, но оставьте два или три верхних. На любой части черенка, которая будет находиться ниже поверхности воды, не должно быть листьев. | |
Удалите все имеющиеся цветы Цветы не помогают процессу укоренения. Если оставить их на черенке, цветы попытаются развиться в семена и использовать пищу, оставшуюся в черенке, которую лучше использовать для укоренения. Отмирающие цветы также будут плесневеть и гнить во влажной среде укоренения. Итак, как бы трудно это ни было, удалите все цветы или бутоны с черенков. | |
Готов к рутированию После обрезки до узла и удаления нижних листьев и цветков черенок готов к укоренению в воде. | |
Резка в воде Несколько черенков можно поместить вместе в один контейнер. Обязательно добавляйте свежую воду по мере необходимости, пока черенки полностью не укоренятся. | |
Проверить наличие рута Укоренение обычно происходит через 3-4 недели, но некоторым растениям требуется больше времени. Когда корни достигают длины 1-2 дюйма или больше, черенок готов к пересадке в горшки. | |
Удаление из воды при укоренении Это растение имеет сильные корни и готово к перемещению в горшок с горшечной почвой. В большинстве случаев, когда корни имеют длину 1-2 дюйма, их можно переместить в отдельные горшки, но многие укоренившиеся черенки выживают в воде в течение продолжительных периодов времени. | |
Предварительно увлажните почву для горшков Аккуратно добавляйте воду в почву, тщательно перемешивая, пока почва не станет влажной, но не мокрой. | |
Выберите горшок подходящего размера и поместите почву на дно Выберите горшок, в котором достаточно места для корневой системы. Поместите достаточное количество предварительно увлажненной почвы на дно горшка, чтобы верхняя часть корневого кома поднялась примерно на дюйм ниже края горшка. | |
Наполнить горшок почвой Заполните пространство вокруг корней предварительно увлажненной почвой. Аккуратно прижмите среду вокруг растения, чтобы обеспечить хороший контакт между корнями и почвой. | |
Обрезать лишнюю листву Этот черенок значительно вырос в воде после того, как был укоренен и до того, как был пересажен в горшок. Высокий рост лучше всего обрезать, чтобы стимулировать новый рост у основания растения и получить более густое молодое растение. Удаленный участок может стать новым черенком, который можно поместить в воду для укоренения. | |
Вода Хорошо полейте почву, пока вся почва не станет влажной и вода не начнет вытекать из дна горшка. Это поможет осесть почве вокруг корней и удалить воздушные карманы. | |
Место в хорошо освещенном месте Как только лишняя вода вытечет из кастрюли, поставьте кастрюлю на блюдце. Не позволяйте растению стоять в воде. Поместите его в подходящее место для выращивания. Растения предъявляют разные требования к свету, влажности и температуре. Знай свое растение. Для получения информации о том, как выращивать бегонии с мочковатыми корнями, нажмите здесь. | |
Счастливое материнское растение Даже материнское растение, давшее черенки, после «стрижки» становится счастливее. Через 3 недели молодые побеги у основания материнского растения вырастают, образуя кустистое привлекательное растение, которое вскоре может дать еще больше черенков для укоренения. |
Сад не был бы Садом без наших членов, доноров и волонтеров.
- Члены
- посетителей
- Садовники
- Садоводы
- Педагоги
- Исследователи
- Поддержка сада
- Участников
- Магазин
- Волонтер
- Вакансии
- СМИ
- Контакт
4344 Shaw Blvd.
(314) 577-5100 часов и входной билет
15050 Faust Park
Chesterfield, MO 63017
(314) 577-0888 часов и входной билет
307 Pinetum Loop Rd,
Grey Summit, MO 914039 3 (75-55 914039 3 (75-59) 914039 часы и прием
Политика конфиденциальности | Условия | Авторское право | Принадлежности | Карта сайта
КОРНЕВОЕ РАЗВИТИЕ ОВОЩНЫХ КУЛЬТУР
КОРНЕВОЕ РАЗВИТИЕ ОВОЩНЫХ КУЛЬТУРДОМАШНЯЯ БИБЛИОТЕКА
Профессор экологии растений, Университет штата Небраска
Преподаватель ботаники, Университет Небраска
НЬЮ-ЙОРК: 370 SEVENTH AVENUE
ЛОНДОН: 6 & 8 BOUVERIE, ST.; ЕС 4
РОБЕРТ ДЖОН УИВЕР
Эта книга в значительной степени основана на исследованиях
авторами. Отсутствие точных знаний и часто
совершенно неточные утверждения о характере и масштабах
корневых систем овощных культур показали необходимость
интенсивное изучение. При работе с различными овощными культурами
обычно добавлялись очень скудные данные других исследований
представить, насколько это возможно, общее представление о развитии корня
овощных культур в США. Не было предпринято ни одной попытки
включить все коренные расследования в зарубежных странах
но было дано достаточно, чтобы адекватно дополнить американские
и, как считается, включает в себя большинство важных выводов. Приведенная литература направит студента, который, возможно, пожелает посмотреть
далее в зарубежные расследования.
Настоящая работа является дополнением к книге Уивера. «Корневое развитие полевых культур». Он предназначен для использование исследователей и производителей, а также для удовлетворения потребности студентов-огородников.
При изучении корневых систем по отношению к культурным практики, «Овощные культуры» Томпсона, «The Vegetable Crops» Бэйли. Принципы огородничества» и подобные работы оказался очень полезным. Но во всех случаях первоисточники свободно консультировались в попытке соотнести корневые отношения с садовой практикой.
Все чертежи с сопутствующим корнем
описания оригинальные. Из-за большого труда и затрат
участие в этой работе, она не могла бы быть выполнена иначе, как
за поощрение и финансовую поддержку, оказанную Фондом Карнеги
Институт Вашингтона. Для этого учреждения авторы являются
глубоко обязан за разрешение опубликовать эту книгу в качестве
сопутствующий том к «Развитию корней полевых культур». Авторы также признательны доктору Герберту С. Хэнсону за
ценную помощь в раскопках корней в Линкольне.
Старший писатель также многим обязан своим ученикам.
и особенно мистеру Т. Л. Стайгеру, миссис Л. Б. Мэтьюз и мисс
Энни Могенсен за большую часть иллюстраций и другие ценные материалы.
помощь. Авторы выражают благодарность профессору Т.
Дж. Фицпатрику за полезную критику при чтении рукописи.
и доказательство.
АВТОРЫ.
ЛИНКОЛЬН, НЕБРАСКА,
июнь 1927 г.
Изменение корневой среды — Адаптация корней к среде
— Корневая адаптация и растениеводство — Взаимоотношения растений,
почва и климат. Деятельность корней в недрах. Метод.
изучения корней — Условия для роста в Линкольне, штат Небраска — Почвы
— Количество и размер площадок — Обработка почвы — Осадки — SoiI
влажность — Температура — Влажность — Испарение — Условия
для выращивания в Нормане, штат Оклахома — Почва — Метод посадки и
обработка почвы — Осадки — Влажность почвы — Другие факторы —
Условия выращивания осенью и зимой.
Раннее развитие. Отношение площади поглощения к влажности почвы — Рост в середине лета — Созревающие растения — Резюме — Сравнение с корнями полевой кукурузы — Связь корневых привычек с производством сельскохозяйственных культур — Выращивание — Удобрения — Проращивание.
Лук Southport White Globe — Раннее развитие — Эффект структуры почвы на развитие корней — Половина взрослых растений — Зрелые растения — Отмирание старых корней — Желтый бермудский лук — Раннее развитие — Полусросшие растения — Рост в течение зима и вторая весна и лето — Резюме — Другие исследования на луке — Связь корневой системы с культурной практикой.
Раннее развитие — Наполовину взрослые растения — Зрелые растения — Резюме.
Раннее развитие — Рост зимой — Взрослые растения
— Резюме.
Раннее развитие — Зрелые растения — Резюме — Другие исследования на спарже — Привычка подземных частей по отношению к культурным практика — Выбор почвы — Пересадка — Выращивание рассады — Постоянная грядка — Хранение продуктов в связи со сбором урожая — Отношение к мульче и перегною — Резюме.
Зрелая корневая система — Резюме — Отношение корневой системы к культурная практика.
Раннее развитие — Рост в середине лета — Зрелые растения — Развитие корнеплодов семенной свеклы — Резюме — Другие исследования на свекле — Привычка укоренения по сравнению с сахарной свеклой — Привычка укоренения по отношению к культурной практике.
Раннее развитие — Половина — взрослые растения — Развитие в середине лета
— Развитие корней семенных растений — Сравнение развития корней
со свеклой — Резюме — Другие исследования мангольда —
Корневое развитие в связи с культурной практикой. Влияние на
структура почвы.
Раннее развитие — Половина — взрослые растения — Взрослые растения — Резюме — Другие исследования шпината — Развитие корня по отношению к культурной практике.
Капуста на Копенгагенском рынке — Раннее развитие — Середина лета рост — Взрослые растения — Резюме — Ранняя плоская голландская капуста — Зрелая корневая система — Рассада капусты — Развитие корней и ботва, затронутые культивацией — Другие исследования по капуста — Связь корневой системы с культурной практикой — Выращивание — Пересадка — Связь корней с конкуренцией и болезнями.
Раннее развитие — Наполовину взрослые растения — Зрелые растения — Резюме — Другие исследования цветной капусты — Корневые привычки по отношению к культурной практике.
Раннее развитие — Позднее развитие — Взрослые растения —
Резюме — Другие исследования коль-раби.
Раннее развитие — Рост в середине лета — Взрослые растения — Корень развитие второго сезона — Краткое содержание — Другие расследования на репе — Корневое развитие по отношению к культурной практике.
Раннее развитие — Позднее развитие — Зрелые растения — Резюме.
Взрослые растения — Резюме — Корневой габитус по отношению к культурным упражняться.
Ранняя алая репа белая — редька с кончиками — Раннее развитие
— Влияние структуры почвы на развитие корней — Двухмесячный возраст
Растения — Взрослые растения — Резюме — Ранняя длинная алая редька
— Раннее развитие — Позднее развитие — Взрослые растения —
Резюме — Другие исследования редьки — Развитие корневой системы
по отношению к культурной практике.
Взрослые растения — Резюме — Другие исследования земляники — Коренные привычки по отношению к культурной практике.
Раннее развитие — Позднее развитие — Взрослые растения — Резюме — Другие исследования гороха — Развитие и Роль корневых бугорков — Развитие корня по отношению к культурная практика.
Обыкновенная или фасоль — Раннее развитие — Рост в середине лета — Взрослые растения — Резюме — Лимская фасоль — Ранний рост — Середина лета рост — Зрелые растения — Резюме — Другие исследования по фасоль — коренная привычка по отношению к культурной практике.
Раннее развитие — Позднее развитие — Зрелые растения — Резюме — Другие исследования бамии.
Раннее развитие — Рост в середине лета — Зрелые растения —
Рост моркови зимой и весной — Взрослые растения —
Резюме — Другие исследования моркови — Корневая привычка в отношении
к культурной практике.
Champion Moss Curled петрушка — Раннее развитие — Середина лета рост — Резюме — Петрушка Extra Double Curled — Другие исследования на петрушку — Корневая привычка по отношению к культурной практике.
Раннее развитие — Рост в середине лета — Растения осенью — Рост корней зимой и весной — Взрослые растения — Резюме — Другие исследования пастернака — Корневая привычка в отношении к культурной практике.
Методы размножения — Ранний рост — Рост в середине лета — Взрослые растения — Резюме — Корневые привычки некоторых сладких растений — родственники картофеля — коренная привычка в связи с культурной практикой — Почвы и удобрения — Водные отношения — Культивация, укоренение виноградников — Обрезка и размещение — Сбор урожая — Резюме.
Раннее развитие корневой системы и его связь с пересадкой —
Более поздний рост — Созревающие растения — Резюме — Другие исследования
на томатах — Опыты по корнеобразованию, выращиванию,
и водосбережение — Коренные привычки по отношению к культурным
практика — Подготовка почвы — Пересадка — Расстояние между растениями
— Культивация — Удобрения — Обрезка — Влияние корней на
почвы.
Раннее развитие — Зрелые растения — Резюме — Другие исследования на баклажанах.
Раннее развитие — Половина — взрослые растения — — Зрелые растения — Резюме — Коренные привычки по отношению к культурной практике.
Тыквенные или виноградные культуры. Раннее развитие огурцов — Наполовину выросшие растения — Созревающие растения — Резюме — Другие исследования на огурце — Корневая система в связи с культурной практикой.
Раннее развитие — Наполовину взрослые растения — Зрелые растения — Резюме — Другие исследования дыни — Корневая привычка в отношение к культурной практике.
Раннее развитие — Наполовину взрослые растения — Взрослые растения —
Резюме — Другие исследования арбузов — Корневые привычки
по отношению к культурной практике.