Таблица рода: Род имён существительных (мужской, женский, средний, общий)
Род имён существительных (мужской, женский, средний, общий)
Род имён существительных — это постоянный грамматический признак слов этой части речи.
Узнаем, что такое род имён существительных и как его определить.
Что такое род имён существительных?
В русском языке все существительные, имеющие форму единственного числа, обладают категорией рода. У слов этой части речи, обозначающих предмет, обязательно указывают один из трёх родов — мужской, женский или средний:
- старшина, богатырь (мужской род)
- улица, речь (женский род)
- стекло, ущелье (средний род)
Важно!
Род имеют только существительные в форме единственного числа.
У слов, находящихся в форме множественного числа или имеющих форму только множественного числа, род не определяется:
- дома́
- скатерти
- каникулы
- вожжи
- хлопоты.

У одушевлённых существительных род находится в прямой зависимости от принадлежности лиц к мужскому или женскому полу:
- папа, дедушка, внук (мужской род)
- мама, тётя, женщина (женский род)
Категория рода существительных выявляется по грамматическим признакам слова в начальной форме именительного падежа единственного числа.
Как определить род существительного?
Род имён существительных определяется обычно с помощью слов-помощников:
- местоимений «он», «мой» для существительных мужского рода;
- «она», «моя» для существительных женского рода;
- «оно», «моё» для существительных среднего рода.
Понаблюдаем:
- он, мой юноша, вечер
- она, моя внучка, столица
- оно, моё создание, добро.
Род существительных определяется также по их характерному окончанию в начальной форме.
Мужской род
К мужскому роду принадлежат одушевлённые существительные I склонения с окончанием -а/-я:
- воевода
- старшина
- дядя
- судья
- слуга
- батюшка
- староста
и пр.
Категорию мужского рода выявим у существительных II склонения с нулевым окончанием:
- матрос
- градусник
- гвоздь
- малыш
В русском языке слово «подмастерье», обозначающее лицо мужского пола, является существительным мужского рода.
Запомним
толь, тюль, рельс — это существительные мужского рода.
Названия людей по профессии (директор, менеджер, врач, инженер) могут обозначать лицо мужского и женского пола, тем не менее они имеют нулевое окончание в форме именительного падежа и являются существительными мужского рода. Согласуемые с ними прилагательные употребляются в форме мужского рода, а род глагола-сказуемого зависит от имени собственного, являющегося подлежащим.
Согласуемые с ними прилагательные употребляются в форме мужского рода, а род глагола-сказуемого зависит от имени собственного, являющегося подлежащим.
Сравним:
По телефону ответил опытный менеджер Николай.
По телефону ответила опытный менеджер Екатерина.
Женский род
Существительные женского рода определим по характерным окончаниям -а/-я или нулевому окончанию в форме именительного падежа единственного числа:
- девушка
- сказка
- репетиция
- кухня
- фасоль
- ветошь
Существительные с нулевым окончанием, с твердой и мягкой основой, могут быть мужского и женского рода.
У существительных женского рода пишется опознавательный мягкий знак (рожь, вещь), а у слов мужского рода его нет (вояж, мякиш). Также их род различают по форме родительного падежа, в которой существительные мужского рода имеют окончание -а/-я, а женского рода — окончание -и:
Также их род различают по форме родительного падежа, в которой существительные мужского рода имеют окончание -а/-я, а женского рода — окончание -и:
- шалаш — постройка (чего?) шалаша
- фонарь — свет (чего?) фонаря
- боль — нет (чего?) боли
- пустошь — площадь (чего?) пустоши
Слово «туфля» имеет категорию женского рода. В форме родительного падежа множественного числа «туфель» (а не туфлей) оно имеет нулевое окончание.
Средний род
К среднему роду в основном принадлежат неодушевлённые существительные с окончаниями -о/-е:
- лукошко
- полено
- созвездие
- море
- счастье
Категорию среднего рода укажем у существительных с конечным буквосочетанием -мя:
Примеры
бремя, вымя, время, пламя, племя, стремя и пр.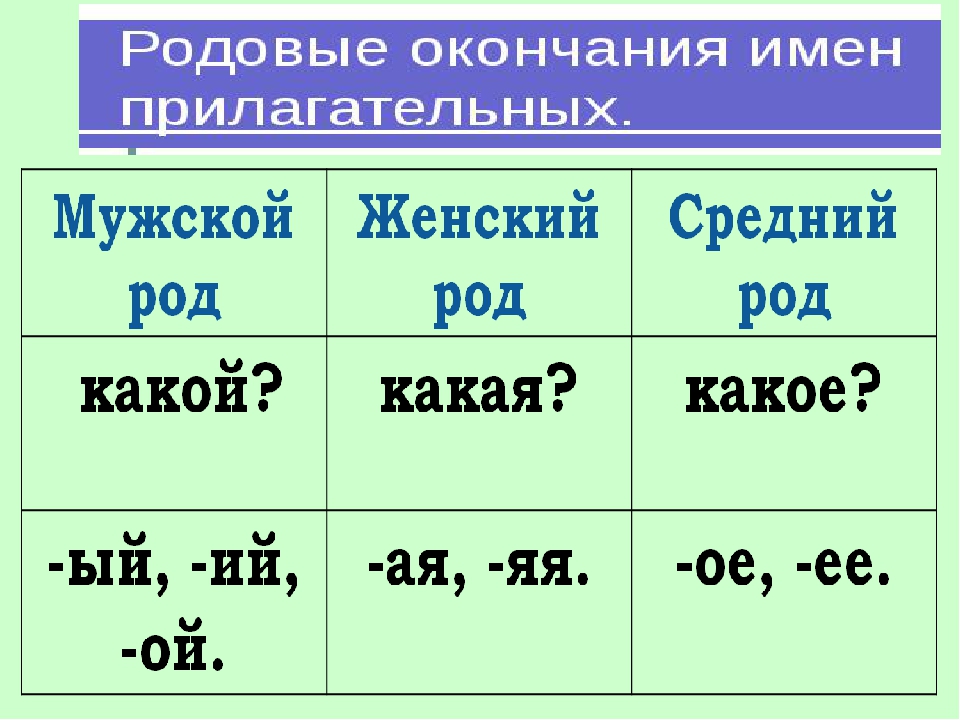
Только несколько одушевленных имён существительных имеют средний род:
- дитя
- насекомое
- животное
- существо
Общий род существительных
В русском языке имеются существительные с окончанием -а/-я, которые могут обозначать носителей определенных качеств:
плакса, разиня, невежа, хитрюга, задира, недотёпа, выскочка, неженка, растяпа и пр.
Среди этих слов укажем стилистически нейтральные по смыслу существительные:
сирота, коллега, умница, визави, протеже, инкогнито и пр.
В одной и той же грамматической форме такие универсальные существительные называют как лиц мужского пола, так и женского:
- маленький тихоня Саша — маленькая тихоня Таня
- круглый сирота Валера — круглая сирота Лена
- шумный забияка Вова — шумная забияка Света.

Эти слова являются существительными общего рода. Их род определяют в зависимости от смысла предложения.
Алёна — такая умница!
Наш Егор просто умница!
Род несклоняемых существительных
Иногда доставляет трудность определить род у несклоняемых имён существительных, которые являются заимствованными словами в русском языке. В определении их рода ориентируемся на значение слова и категорию одушевлённости/неодушевлённости имени существительного.
Одушевленные существительные, называющие человека по признаку пола, в соответствии с их значением имеют категорию мужского или женского рода:
- блестящий маэстро
- озабоченный портье
- остроумный конферансье
- известный кутюрье
- милая леди
- вежливая мадам
- юная мисс
- гордая пани
Иноязычные слова, называющие птиц и животных, имеют категорию мужского рода, если в контексте не указана самка животного:
- поющий колибри
- пёстрый какаду
- низкорослый зебу
- мохнатый шимпанзе
Сравним:
Шимпанзе уселась в уголке, чтобы покормить детёныша.
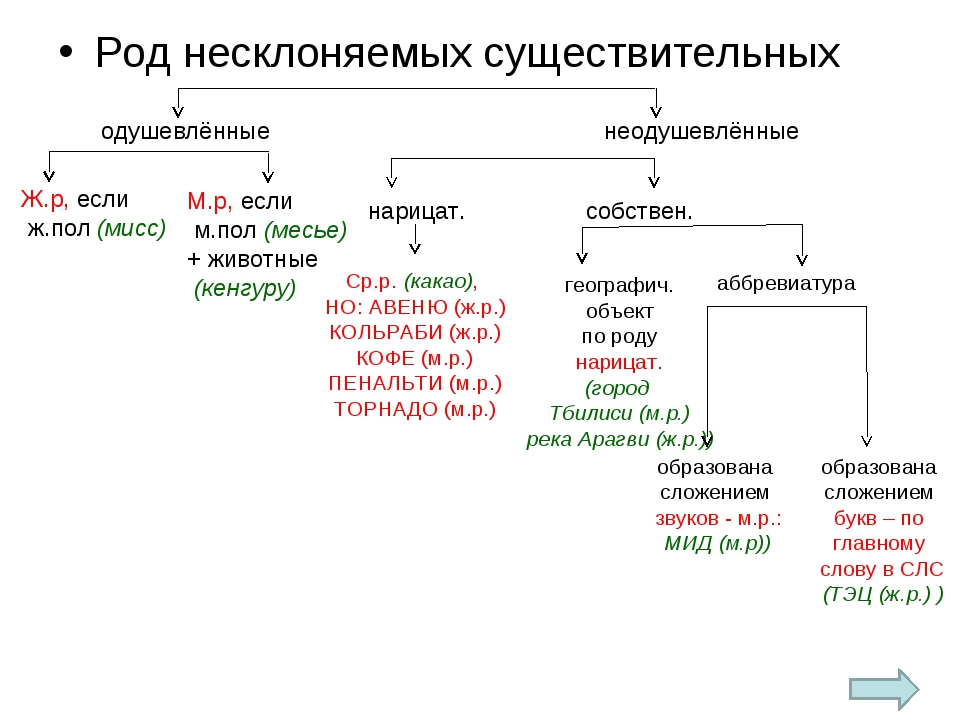
Неодушевленные существительные в основном имеют средний род:
- чистое метро
- весёлое шоу
- уютное кафе
- быстрое такси
Исключение!
кофе (мужской род)
Учтём, что у ряда неодушевлённых существительных род определяется по опорному слову:
- зелёная кольраби (капуста)
- вкусный сулугуни (сыр)
- стремительный торнадо (ураган)
- красивая авеню (улица)
- вкусная киви (ягода)
- соленая иваси (сельдь)
- результативный пенальти (штрафной удар)
Род географический названий и аббревиатур определяется по родовому слову:
- тихое Онтарио (озеро)
- бурная Миссисипи (река)
- уютный Бордо (город)
- ВДНХ (выставка) закрыта
- ГАИ (инспекция) сообщила
Видеоурок
Скачать статью: PDFСуществительные общего рода – примеры, список
Существительные общего рода — это одушевленные существительные первого склонения с окончанием -а/я, которые обозначают лицо женского или мужского пола в зависимости от смысла контекста.
Чтобы понять, что такое существительные общего рода, вспомним, что в русском языке все слова имеют постоянный грамматический признак — род.
Имена существительные, употребляющиеся в форме единственного числа, имеют один из трех родов:
- человек, город (он, мой) — мужской род;
- подруга, дочь, повесть (она, моя) — женский род;
- войско, стремление (оно, моё) — средний род.

Что такое существительные общего рода?
Среди слов этой самостоятельной части речи имеется немногочисленная группа существительных общего рода, объединяющая около 200 слов. Эти уникальные слова могут называть как лицо женского пола, так и лицо мужского пола:
Василий Петрович — это мой коллега.
Мария Васильевна — это моя коллега.
Слово «коллега» обозначает лицо и отвечает на вопрос кто? Оно грамматически оформлено как существительное первого склонения с окончанием -а и соответственно изменяется по падежам и числам:
- и. п. (кто?) коллега, коллеги
- р. п. совет (кого?) коллеги, коллег
- д. п. идем (к кому?) к коллеге, к коллегам
- в. п. спросим (кого?) коллегу, коллег
- т. п. восхищён (кем?) коллегой, коллегами
- п.
 п. сообщите (о ком?) о коллеге, о коллегах
п. сообщите (о ком?) о коллеге, о коллегах
В контексте в зависимости от того, о ком идет речь, оно может называть лицо как мужского, так и женского пола. Такие слова объединены в одну группу «существительные общего рода».
Определение
Существительные общего рода — это одушевленные существительные первого склонения с окончанием -а/-я, называющие лиц мужского или женского пола в зависимости от контекста.
Задиру Ваню, задиру Светку,
Обжору Катьку, обжору Генку
Найдешь в семье ты
И в каждом классе.
А соню Димку и соню Ленку
Опять обсуждали на классном часе.
| Существительные общего рода | |
|---|---|
| 1. нарицательные несклоняемые сущ. | визави, протеже |
| 2. собственные склоняемые имена на -а/-я | Валя, Женя, Саша |
3. нарицательные склоняемые сущ. на -а/-я, характеризующие лицо по склонности, пристрастию к чему-либо и стил. нейтральные нарицательные склоняемые сущ. на -а/-я, характеризующие лицо по склонности, пристрастию к чему-либо и стил. нейтральные | задира, кривляка, мямля, невежа, недотепа, плакса, пьяница, разиня, хитрюга, запевала, книгоноша, сирота, коллега и т.д. |
| Валя и Женя – протеже Нестеренко. | |
| Сущ. общего рода должны иметь определения в грамматических формах того рода (мужского или женского), к которому принадлежат названия характеризуемых ими предметов. | |
| Красавица Валя и бездельник Женя – протеже всё той же Нестеренко. | |
Существительные общего рода образуются от слов разных частей речи с помощью суффиксов -ак/-як-, -енк-, -ин-, -к-, -л-, -ук-, юл-, -яг- и пр.:
- зевать → зевака
- запевать → запевала
- нежный → неженка
- лакомиться → лакомка
- чистый → чистюля
- работать → работяга.

Костя — большой привереда (м. р.)
Вы еще не знаете, какая она привереда! (ж. р.)
Нам навстречу выбежал забияка (м. р.)
Ему повстречалась забияка. (ж. р.)
Употребление существительных общего рода
Среди этих слов только небольшая группа имеет нейтральную или положительную эмоциональную окраску:
- тёзка
- сирота
- левша
- коллега
- умница
- молодец
- молодчина
- почемучка
- самоучка
- всезнайка
Большинство существительных общего рода имеет отрицательный и даже уничижительный смысл. В разговорной речи они дают характеристику человеку, обозначая какой-то отрицательный обобщенный признак личности:
В разговорной речи они дают характеристику человеку, обозначая какой-то отрицательный обобщенный признак личности:
- неряха
- невежда
- разиня
- пустомеля
- выскочка
- зануда
В основном эти слова используются в разговорной речи. Их не применяют в научном или официально-деловом стиле речи. В художественной литературе с их помощью характеризуют персонажей произведения.
Я весь проныра и пролаза (Анатолий Рыбаков. Выстрел).
Ночной бродяга, которого мы видели мельком, шел в этом направлении (Виктор Гюго. Отверженные).
Слова «верзила», «воротила», «вышибала», «детина» имеют категорию мужского рода.
Также отличаем существительные общего рода от названий профессий, которые оформлены как существительные мужского рода, хотя и называют лиц женского пола.
К нам подошла детский врач Алена Федоровна.
Софья — хороший программист.
Нам посоветовали обратиться к Галине Васильевне, отличному технологу.
Приведем список наиболее употребительных существительных общего рода, расположив их по алфавиту.
Список существительных общего рода
- бедняга, бедняжка, бедолага, белоручка, бука, бродяга, брюзга;
- воображала, вояка, всезнайка, выскочка;
- горемыка, грязнуля, гулёна, гуляка;
- душенька, дылда;
- егоза, ехидна;
- жадина, жадюга;
- забияка, задира, зазнайка, заика, зануда, замарашка, заводила, запевала, зевака, злюка, злючка, зубрила;
- калека, каналья, коллега, коротышка, кривляка, кровопийца, кроха;
- лакомка, левша, лежебока;
- молодец, молодчина, мямля;
- невежа, невежда, невеличка, невидимка, недотёпа, недотрога, недоучка, неженка, незнайка, непоседа, неровня, неряха, неумека, неумеха;
- обжора, одиночка;
- писака, пискля, плакса, подлиза, подпевала, попрошайка, почемучка, привереда, придира, пройдоха, проныра, простофиля, пустомеля, пьяница;
- работяга, разиня, растеряха, растяпа, рёва, рёвушка, ровня;
- самоучка, сквалыга, скромняга, сладкоежка, сластёна, соня, сутяга;
- тараторка, тёзка, тихоня, трещотка, трудяга, тупица;
- умница
- ханжа, хапуга, хвастунишка, хитрюга, худышка;
- чистюля
- шельма
- юла
- ябеда
Видеоурок
Скачать статью: PDFСклонение существительных (таблица и примеры)
Склонение имен существительных — это изменение по падежам в соответствии с родовой принадлежностью.
В русской морфологии различают основные три типа склонения имен существительных. Но кроме них, выделяют еще несколько склонений.
Имена существительные в русском языке изменяются по падежам и числам. Это изменение формы слова называется склонением.
Определение
Изменение имен существительных по падежам и числам в русском языке называется склонением.
В соответствии с родовой принадлежностью и падежными окончаниями в форме единственного числа существительные делятся на три склонения.
Первое склонение
К первому склонению отнесем существительные мужского и женского рода с твердой и мягкой основой с окончанием -а/-я в форме именительного падежа, например:
- мужчина
- дядя
- юноша
- мальчишка
- берёста
- дорога
- няня
- деревня.
Сюда же добавим существительные общего рода, которые могут в зависимости от контекста называть как лиц мужского пола, так и женского, например:
Примеры
трудяга, забияка, плакса, молодчина, непоседа, выскочка, злюка, неряха
- Забияка Ваня
- плакса Валя
- непоседа Серёжка
- молодчина Стасик.
Склонение существительных на -ия
Отметим, что ряд существительных женского рода первого склонения заканчивается на -ия:
- история
- стадия
- лекция.
Такие слова в формах дательного и предложного падежа единственного числа имеют окончание -и в отличие от типичных существительных первого склонения, как например слово «трава».
Сравним:
- и. п. (что?) лекция, трава
- р. п. время (чего?) лекции, нет травы
- д. п. уделим внимание (чему?) лекции, траве
- в. п. вижу (что?) лекцию, траву
- т. п. восхищаемся (чем?) лекцией, травой
- п. п. сообщит (о чём?) о лекции, о траве
Второе склонение
Второе склонение объединяет существительные мужского рода с нулевым окончанием, а также с окончанием -о/-е и среднего рода с окончанием -о/-е в форме именительного падежа, например:
- гений , автор , герой , соловей ;
- домище, человечище, заборишко, голосишко;
- слово, дело, село, поле, море, собрание, варенье.
Склонение существительных среднего рода на -ие
Особое внимание обратим на существительные среднего рода, которые заканчиваются на буквосочетание -ие:
- наклоне́ние
- сочета́ние
- указа́ние.
Такие существительные склоняются по-особому: в формах единственного числа предложного падежа отметим окончание -и, а не -е, как у слов среднего рода типа «озеро» или «поле».
Понаблюдаем и сравним:
- и. п. (что?) указание, озеро
- р. п. нет (чего?) указания, озера
- д. п. удивляемся (чему?) указанию, к озеру
- в. п. читаю (что?) указание, вижу озеро
- т. п. интересуюсь (чем?) указанием, озером
- п. п. расскажу (о чём?) об указании, об озере
Третье склонение
К третьему склонению относятся имена существительные женского рода с нулевым окончанием в именительном падеже, например:
- быль
- радость
- сирень
- лазурь
- роскошь
- горечь
- блажь
- вещь .
Таблица «Три склонения имен существительных»
| Склонение | Род | Окончание | Примеры |
|---|---|---|---|
| Первое | мужской | -а, -я | юноша дядя |
| женский | птица земля | ||
| Второе | мужской | нулевое | отец день |
| средний | -о, -е | зерно счастье | |
| Третье | женский | нулевое | дочь рожь |
Разносклоняемые существительные
В отдельную группу выделим разносклоняемые существительные среднего рода на -мя:
- бремя
- вымя
- темя
- пламя
- стремя
- пламя
- время
- знамя
- имя
- племя
- семя
- существительное мужского рода «путь»
- существительное среднего рода «дитя».
Разносклоняемыми эти слова названы потому, что частью они склоняются как существительные третьего склонения, а в форме творительного падежа имеют окончание второго склонения.
Сравним:
- и.п. пламя, дитя, путь, ночь
- р.п. пламени, дитяти, пути, ночи
- д.п. пламени, дитяти, пути, ночи
- в.п. пламя, дитя, путь, ночь
- т.п. пламенем, дитятей (-ею), путём, ночью
- п.п. о пламени, о дитяти, о пути, о ночи.
Другие типы склонения существительных
В русском языке целый пласт лексики составляют существительные — бывшие прилагательные или причастия. Они сохранили грамматическую форму прежней части речи и склоняются, как прилагательные. Эти слова отнесем к отдельному типу склонения существительных, например:
- служащий
- провожающий
- прибывший
- слагаемое
- прохожий
- ванная
- запятая
- горничная
- горячее
- сказуемое
- мороженое.
Отдельный тип склонения составляют существительные, которые имеют только форму множественного числа. У этих слов невозможно определить родовую принадлежность и, следовательно, отнести к одному из трех основных типов склонения.
Примеры
трудяга, забияка, плакса, молодчина, непоседа, выскочка, злюка, неряха
грабли, брюки, весы, качели, румяна, перила, чернила, хлопья, сливки, консервы, будни, сутки, каникулы, городки, шахматы, проводы, выборы, дебаты
Особняком находятся несклоняемые существительные, которые во всех падежах сохраняют одну и ту же форму.
Примеры
метро, пальто, кенгуру, зебу, колибри, сулугуни, драже, трюмо, каноэ, алоэ, шасси, такси, жалюзи, бра
Определение склонения имени существительного
Видеоурок «Склонение имен существительных»
Скачать статью: PDFСклонение имен прилагательных по падежам: таблица
Имена прилагательные в полной форме изменяются по падежам. Чтобы узнать падеж прилагательного, надо найти существительное, к которому оно относится, и определить его падеж. В этом же падеже будет стоять и прилагательное.
Склонение имён прилагательных
Склонение прилагательных — это их изменение по падежам, числам и родам. Род, число и падеж прилагательного зависит от существительного, к которому это прилагательное относится. При склонении прилагательных у них изменяются окончания в зависимости от рода и числа.
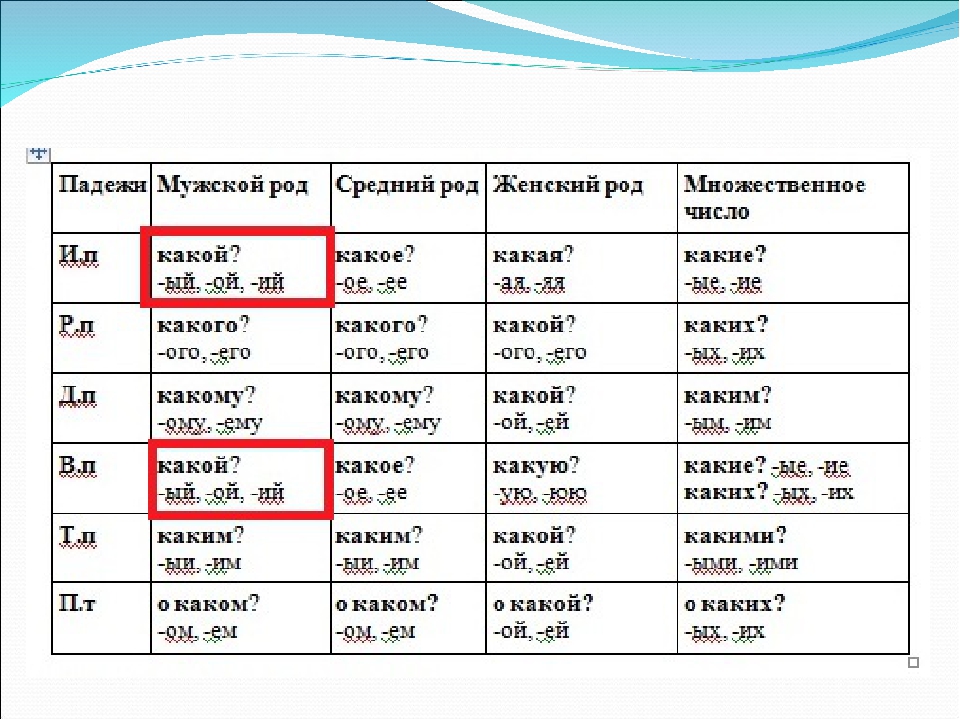
| Падежи | Мужской род | Женский род | Средний род | Множественное число |
|---|---|---|---|---|
| И.п. | -ой, -ый, -ий | -ая, -яя | -ое, -ее | -ые, -ие |
| Р.п. | -ого, -его | -ой, -ей | -ого, -его | -ых, -их |
| Д.п. | -ому, -ему | -ой, -ей | -ому, -ему | -ым, -им |
| В.п. | -ую, -юю | -ое, -ее | -ые (-ых), -ии (-их) | |
| Т.п. | -ым, -им | -ой, -ей | -ым, -им | -ыми, -ими |
| П.п. | -ом, -ем | -ой, -ей | -ом, -ем | -ых, -их |
Склонение имён прилагательных в единственном числе мужского рода
| Падеж | Вопросы | Примеры |
|---|---|---|
| И.п. | какой? | лесной шум, мудрый друг, летний день, синий кит |
| Р.п. | какого? | лесного шума, мудрого друга, летнего деня, синего кита |
| Д.п. | какому? | лесному шуму, мудрому другу, летнему дню, синему киту |
| В.п. | какой? какого? | лесной шум, мудрого друга, летний день, синего кита |
| Т.п. | каким? | лесным шумом, мудрым другом, летним днем, синем китом |
| П.п. | о каком? | о лесном шуме, о мудром друге, летнем дне, синем ките |
Винительный падеж прилагательных мужского рода совпадает по форме с именительным, если прилагательное относится к неодушевлённому существительному: вижу большой дом (В. п.) — большой дом (И. п.).
Винительный падеж прилагательных мужского рода совпадает по форме с родительным, если прилагательное относится к одушевлённому существительному: вижу большого медведя (В. п.) — нет большого медведя (Р. п.).
Прилагательные мужского рода на -ОЙ склоняются так же, как и на -ЫЙ, но всегда имеют ударные окончания: большо́й, большо́го, большо́му, больши́м, о большо́м.
Склонение имён прилагательных в единственном числе женского рода
| Падеж | Вопросы | Примеры |
|---|---|---|
| И.п. | какая? | лесная птица, синяя лента |
| Р.п. | какой? | лесной птицы, синей ленты |
| Д.п. | какой? | лесной птице, синей ленте |
| В.п. | какую? | лесную птицу, синюю ленту |
| Т.п. | какой? | лесной птицей, синей лентой |
| П.п. | о какой? | о лесной птице, о синей ленте |
Склонение имён прилагательных в единственном числе среднего рода
| Падеж | Вопросы | Примеры |
|---|---|---|
| И.п. | какое? | яркое солнце, древнее зло |
| Р.п. | какого? | яркого солнца, древнего зла |
| Д.п. | какому? | яркому солнцу, древнему злу |
| В.п. | какое? | яркое солнце, древнее зло |
| Т.п. | каким? | ярким солнцем, древним злом |
| П.п. | о каком? | о ярком солнце, древнем зле |
Склонение полных прилагательных во множественном числе
| Падеж | Вопросы | Примеры |
|---|---|---|
| И.п. | какие? | новые люди, яркие сны |
| Р.п. | каких? | новых людей, ярких снов |
| Д.п. | каким? | новым людям, ярким снам |
| В.п. | каких? какие? | новых людей, яркие сны |
| Т.п. | какими? | новыми людьми, яркими снами |
| П.п. | о каких? | о новых людях, о ярких снах |
Винительный падеж прилагательных множественного числа совпадает по форме с именительным, если прилагательное относится к неодушевлённым существительным: вижу большие дома (В. п.) — большие дома (И. п.).
Винительный падеж прилагательных множественного числа совпадает по форме с родительным, если прилагательное относится к одушевлённым существительным: вижу синих китов (В. п.) — нет синих китов (Р. п.).
Некоторые прилагательные перешли в разряд существительных, например: больной, рабочий, ванная, мастерская и так далее. Такие существительные при склонении имеют падежные окончания прилагательных: рабочий, рабочего, рабочему.
Чтобы проверить написание окончания прилагательного, надо задать к нему вопрос от существительного, например: Я давно не видел белого песка (песка какого? белого).
Окончание прилагательного будет полностью или частично совпадать с окончанием вопроса. Совпадение окончаний будет зависеть от склонения.
Склонение прилагательных делится на три типа: твёрдое, мягкое и смешанное. К твёрдому склонению относятся прилагательные с основой, которая оканчивается на твёрдый согласный: длинный, старый.
К мягкому склонению относятся прилагательные с основой, которая оканчивается на мягкий согласный: синий, летний.
Смешанное склонение
К смешанному склонению относятся прилагательные с основой, которая оканчивается на Г, К, Х, Ц или шипящий: строгий, мягкий, тихий, куцый, хороший, горячий.
| какой? | больной | дальний | мягкий |
| какого? | больного | дальнего | мягкого |
| какому? | больному | дальнему | мягкому |
| какой? | больной | дальний | мягкий |
| каким? | больным | дальним | мягким |
| о каком? | о больном | о дальнем | о мягком |
Обратите внимание, что в зависимости от типа склонения, окончание вопроса и прилагательного совпадает либо полностью, либо частично: отличие только в первой букве окончания прилагательного.
Видеоуроки по теме: склонение имен прилагательных
Сколько родов в русском языке? Особенности определения рода у имен существительных и прилагательных
Что такое род применительно к русскому языку, сколько родов в русском языке и какие выделяют?
Категория рода присуща таким частям речи, как имя существительное и имя прилагательное. Род представляет собой категорию, которая группирует словоформы на основании полового признака или его отсутствия.
С этой категорией школьники начинают знакомиться в начальных классах, постепенно углубляя и закрепляя свои знания. В пятом классе сведения о роде пополняются и закрепляются на более сложном материале.
Сколько родов в русском языке?
В русском языке представлена следующая система:
- Женский род.
- Мужской род.
- Средний род.
- Общий род.
Не определяется род у слов, употребляемых только во множественном числе.
Сколько родов в русском языке у имени существительного?
Категория рода имен существительных постоянна. Например, слово “кот” относится к мужскому роду.
Для определения рода имени существительного задаем к данному слову смысловой вопрос: он мой? она моя? оно мое?
| Смысловой вопрос | Род | Показатель рода (в И. п. ед. ч.) | Примеры |
| Мой? | Мужской | Окончания -а/-я, а также нулевое окончание | Ножик, заяц, стол |
| Моя? | Женский | Окончания -а/-я, а также нулевое окончание | Кошка, мама, собака, мебель |
| Мое? | Средний | Окончания -о/-е | Солнце, дерево, озеро |
Как видно из таблицы, род определяется только у существительных в единственном числе. Существительные, употребляемые в только во множественном числе (брюки, очки, сани), находятся вне категории рода.
При определении рода имен существительных у школьников нередко вызывают затруднения слова типа “знайка”, “умница”, “непоседа” и тому подобные. Например: он был большим непоседой и она была большой непоседой. Такие слова относятся к женскому или к мужскому роду? Здесь как раз возникает вопрос, вынесенный в заглавие: сколько родов в русском языке? Ученые на этот счет имеют две точки зрения: одни относят их к мужскому или женскому, в зависимости от контекста, другие выделяют такие слова в особый род – общий.
Также вызывают трудности несклоняемые иноязычные существительные. По написанию они напоминают слова, относящиеся к среднему роду. Действительно, большинство из них относится именно к этому роду, но есть и (скажем так) исключения из правила.
Так, согласно литературной норме, существительное “кофе” относится к мужскому роду. Неправильно будет сказать “мое кофе”. Это ошибка, верный вариант – “мой кофе”.
Существительное “евро”, по аналогии с другими названиями денежных единиц, относится к мужскому роду. По такому же принципу существительные “сулугуни”, “сирокко”, “пенальти” относятся к мужскому. На основании такой же аналогии существительные “авеню”, “салями”, “кольраби” относятся к женскому роду.
Если у вас возникли сомнения по поводу родовой принадлежности имени существительного, то следует обратиться к словарям русского языка.
Как определить род имени прилагательного?
В отличие от имени существительного, для которого категория рода является неизменяемой, у прилагательного он является изменяемой категорией и определяется в зависимости от контекста. Правило, по которому определяют род этой части речи, звучит следующим образом: род прилагательного устанавливают по определяемому слову, то есть по имени существительному.
Например:
- На девочке было надето красивое (ср. р.) платье. (“Платье” – оно мое, следовательно – это средний род, значит, и прилагательное “красивое” относится к среднему роду).
- Он был красивым (м. р.) мужчиной. Улица красива (ж. р.).
Также выделяют несклоняемые прилагательные. Например: брюки цвета хаки.
Теперь вы знаете ответ на вопрос о том, сколько родов в русском языке. Определение их мы тоже разобрали на примерах. Уметь определять род имени существительного или прилагательного очень важно – это поможет избежать грамматических ошибок.
Род – определение и примеры
Род – определение
Род – это ранжирование таксономических категорий, используемое в биологической классификации, которое находится ниже семейства и выше видов. Виды, демонстрирующие сходные характеристики, составляют род. По оценкам, количество опубликованных названий родов составило около 510 000 по состоянию на 2016 год. (1) В 2018 году в Каталоге жизни было указано 173 363 общепринятых названия родов как для существующих, так и для исчезнувших видов. В их отчет также включены названия родов без видов для определенных групп. (2) В биномиальной номенклатуре род используется как первое слово научного названия, в котором первая буква заглавная. Вместе со специфическим эпитетом они выделяются курсивом или добавляются кавычками (“”), например Homo sapiens или «Homo sapiens».
Определение рода
Биологический род определяется как таксономический ранг, состоящий из видов с общими признаками. Он включает группу (ы) видов, которые структурно сходны или филогенетически связаны.
Этимология
Термин род заимствован из латинского языка. Это означает «рождение», происхождение, , «происхождение», «род» или «тип». Форма множественного числа – род . Таким образом, значение родов относится к более чем одному роду, поскольку большинство таксономических семейств состоят из нескольких родов. Синоним: родовое наименование.
Система классификации
Род – это таксономический ранг среди восьми основных таксономических рангов в биологической классификации.Он находится ниже семейства и выше вида. Род может состоять из одного или нескольких видов. Семья, в свою очередь, состоит из одного или нескольких родов.
Классификация организмов – это систематическая группировка живых существ на основе характеристик, иерархических или филогенетических отношений. Это один из основных аспектов таксономии. Организмы исследуются с точки зрения их морфологии, анатомии, физиологии, эволюции, поведения, развития и генетики, чтобы установить взаимосвязь между ними.Затем они классифицируются по таксономическим группам и таксономической иерархии. Общие таксономические уровни: домен , королевство , тип , класс , порядок , семейство , род и виды . Таксономия рода – это ранг, обычно выше уровня вида и ниже уровня семейства. Одним из наиболее выдающихся авторов систематизированной классификации организмов является шведский ботаник Карл Линней. Система классификации, в которой организмы подразделяются на таксоны, называется Линнейской, которая была названа в его честь.Он также предложил биномиальную номенклатуру. В системе Линнея организмы группируются на основе предполагаемой гомологии, то есть сходства по анатомическим, морфологическим и физиологическим особенностям. Чем больше гомологичных структур разделяют организмы, тем больше они, вероятно, эволюционно связаны.
Система имен
В биномиальной номенклатуре род используется как первое слово научного названия. Название рода всегда пишется с заглавной буквы и курсивом. Например, биномиальное имя льва – Panthera leo .Первая часть, Panthera , является названием рода, а вторая часть, leo , является специфическим эпитетом. Таксономист (специалисты по таксономии) присваивает научное название конкретному виду. Чтобы род был полезен для описания, он должен обладать монофильностью, разумной компактностью и отличимостью. Вилли Хенниг, немецкий биолог, определил монофилию как группы, основанные на общих производных характеристиках или чертах, которые отличают группу от других групп организмов. Что касается разумной компактности, то это означает, что род не нужно без надобности расширять.Название рода должно также демонстрировать различимость в отношении соответствующих эволюционных критериев, таких как экология, морфология или биогеография. В классификации и наименовании родов номенклатурные коды обеспечивают архетипический стандарт. Биномиальное имя отличается от общеупотребительного или народного имени. Последний не стандартизирован и варьируется в зависимости от местоположения, в отличие от первого, который стандартизован и используется во всем мире.
Род против видов
Вид организмов считается основной единицей или категорией в биологической системе классификации.Чтобы попасть в ранг вида, группа должна иметь по крайней мере двух членов, способных воспроизводить потомство (особенно через половое размножение). Организмы разных видов, хотя и принадлежат к одному роду, обычно не могут скрещиваться, поскольку их потомство, вероятно, будет бесплодным. Следовательно, виды особой группы организмов будут относиться к тем, которые могут воспроизводить и производить плодовитое потомство того же вида. Таким образом, они будут иметь одинаковый набор ДНК, сходные физические и морфологические характеристики и демонстрировать общее поведение.Вид находится ниже ранга рода. Таким образом, род является более всеобъемлющим и относительно более обширным, чем вид. Тем не менее, род находится ниже таксономического семейства и, следовательно, будет менее инклюзивным, чем семейство, поскольку последнее служит коллективным зонтиком родственных родов. Определенные виды могут быть далее подразделены на подгруппы (называемые подвидами), такие как разновидности, формы и т. Д. Формат род-вид важен для наименования организма. Род – это родовое название , тогда как вид – это специфическое название в биномиальной номенклатуре.Например, Allium cepa (широко известный как лук). Allium – это общее название, а cepa – конкретное название.
Таблица 1: Различия между родом и видом
| Род | Виды |
|---|---|
| Значение рода: таксономический ранг ниже семейства и выше видов | Значение вида: основная единица в биологической классификации |
| Состоит из видов с общими признаками | Состоит из особей, которые могут скрещиваться и давать плодовитое потомство |
| Более инклюзивный, поскольку состоит из разных видов и, следовательно, состоит из большего числа организмов меньшее количество организмов | |
| Первая часть биономиального имени | Вторая часть биономического имени |
| Пример рода: Homo | Пример вида: H.sapiens |
Род и семейство
Таксономическое семейство – это группа из одного или нескольких родов. Роды в определенной семье имеют общий атрибут. Таким образом, семья обычно более инклюзивна и состоит из большего числа организмов. Роды определенной семьи произошли от одних и тех же предков и имеют относительно общие характеристики. В таксономическом ранге семья находится выше уровня рода и ниже уровня порядка.
Типовая концепция
В современной биологической классификации типовой род служит представителем таксономического семейства.Таким образом, один или несколько родов в семье будут определять последнее. В зоологии Международный кодекс зоологической номенклатуры обеспечивает фундаментальные стандарты. Соответственно, название группы семейства будет основано на типовом роде. Например, род Cricetus (Leske, 1779) является типовым родом семейства Cricetidae. Другой пример – кряква Anas platyrhynchos . Anas – типовой род семейства Anatidae. Canis lupus (собаки и волки) относятся к семейству Canidae .Фамилия происходит от общего названия Canis . Род может служить корнем , а название семейства – корнем , с названием, часто заканчивающимся на – idae . Есть также случаи, когда следующий основной таксономический ранг, особенно порядок, также основан на типовом роде. Собаки и волки, например, принадлежат к отряду хищных. В фамилии должен быть типовой род, так же как типовой род имеет типовой вид . Если оказывается, что экземпляр принадлежит к другому роду, то родовое название становится младшим синонимом .
Использование
В зоологии род может быть доступен или недоступен . Доступные наименований – это названия родов, опубликованные на основе стандартов Международного кодекса зоологической номенклатуры (ICZN), а также Международной комиссии по зоологической номенклатуре. недоступен – это те, которые не были опубликованы из-за несоответствия Кодексу ICZN. Другие возможные причины – неправильное написание и отсутствие типовых видов.
В ботанике доступное имя является правильно опубликованным именем, тогда как недоступное имя является недопустимым именем. Недействительному названию рода присваивается метка nomen invalidum (nom. Inval.) . Действительное имя в ботанике обозначается как правильное имя или текущее имя .
Название рода могло со временем меняться и заменяться другим. Это происходит, когда приходит новая информация. В результате ранее принятое имя становится синонимом .Например, Physeter (Linnaeus, 1758) имеет следующие синонимы: Catodon (Linnaeus, 1761), Cetus (Billberg, 1828), Meganeuron (Gray, 1865), Megistosaurus (Harlan, 1828). ), Phiseter (Bonnaterre, 1789), Physalus (Lacépède, 1804), Physeterus (Duméril, 1806) и Tursio (Fleming, 1822). (2) Омоним в биологической классификации – это название, которое связано с двумя разными таксонами.Например, жук-амброзия и утконос получили родовое название Platypus . Тем не менее, жук-амброзия первым получил родовое название Platypus , и, таким образом, утконос в конечном итоге получил новое родовое имя Ornithorhynchus . Эти двое не могут иметь одно и то же общее название, так как они оба из Kingdom Animalia. Однако использование одних и тех же родов для образцов из разных царств по-прежнему не рекомендуется. Существуют тысячи случаев, когда виды из разных царств относятся к одним и тем же родам.Например, Aoutus – это общее название ночных обезьян, а также золотого горошка.
Примеры
Homo (от латинского, что буквально означает «человек») – это род людей, принадлежащих к трибе Hominini семейства Hominidae, отряда приматов, класса Mammalia. Фундаментальные черты человеческого вида из рода Homo – это двуногие, противопоставление большого пальца, наличие хорды, которая в конечном итоге заменяется позвоночником, живорождение и молочные железы, производящие грудное молоко у женщин для питания новорожденных.В этот пример рода включены несколько видов. Однако сохранился только один вид – H. sapiens sapiens (современный). Ниже приводится список родов человеческих видов (род Homo).
- H. habilis
- H. rudolfensis
- H. gautengensis
- H. erectus
- H. ergaster
- H. antecessor H. antecessor H.
- H.cepranensis
- H. rhodesiensis
- H. naledi
- H. neanderthalensis
- H. floresiensis
- H. tsaichangensis
- Denisova Dein Х. с. sapiens (современный)
Эти виды обладают высокоразвитым мозгом и продвинутыми навыками, особенно в абстрактном мышлении, решении проблем, самосознании и четком общении.Они ходят на двух ногах, в прямой тележке. У них относительно меньшие зубы, чем у других приматов. Эти особенности отличают их от других родов, таких как Australopithecus . Род Australopithecus также принадлежит к трибе Hominini. Их мозг был меньше, примерно 35% от размера мозга современного человека. Как правило, они были меньше людей и были короче (от 3’11 до 4’7 ). Они были более морфологически похожи на шимпанзе и бонобо, чем на людей, поскольку их тело полностью покрыто волосами. Астралопитек , однако, играет определенную роль в эволюции человека. Предполагается, что род Homo произошел от одного из видов этого рода миллион лет назад. (4) Ardipithecus – еще один род, принадлежащий к семейству Hominidae. Представители этого рода уже вымерли. Они отошли от шимпанзе. У них хватка большого пальца стопы или большого пальца ноги, что позволяло им относительно легко переходить с одного дерева на другое. Этот род как самый ранний предок человека является предметом споров, поскольку они, вероятно, ведут себя больше как шимпанзе, чем люди. Sahelanthropus – это род, состоящий из вымерших видов из эпохи миоцена, особенно в период, близкий к тому времени, когда шимпанзе и люди расходились.
См. Также
Ссылки
- Рис, Т., Вандепитт, Л., Декок, В., и Ванхорн, Б. (2017). «IRMNG 2006–2016: 10 лет глобальной таксономической базы данных» (PDF). Информатика биоразнообразия. 12: 1–44.
- Каталог жизни – Годовой контрольный список 2018: Годовой контрольный список 2018. (2018). Получено из Catalogueoflife.org: http://www.catalogueoflife.org/annual-checklist/2018/info/ac
- WoRMS – Всемирный регистр морских видов – Physeter Linnaeus, 1758. (2019). Взято с веб-сайта Marinespecies.org: http://www.marinespecies.org/aphia.php?p=taxdetails&id=137032
- Тот, Н. и Шик, К. (2005). «Африканское происхождение» в книге «Человеческое прошлое: мировая предыстория и развитие человеческих обществ» (редактор: Крис Скарр). Лондон: Темза и Гудзон. Страница 60.
- AVH – Ссылки на этот вид на других веб-серверах.(2010). Получено с веб-сайта Anbg.gov.au: http://www.anbg.gov.au/chah/avh/help/names/index.html
- What Is a Genus: Common Trees of the Pacific Northwest. (2019). Получено с веб-сайта Oregonstate.edu: https://oregonstate.edu/trees/genus_describe.html
- Необходимые таксоны – люди. (2019). Получено с веб-сайта Ncsu.edu: https://projects.ncsu.edu/cals/course/zo150/mozley/humans.html
- Классификация живых существ: принципы классификации. (2012). Получено Паломар.Веб-сайт edu: https://www2.palomar.edu/anthro/animal/animal_2.htm
© Biology Online. Контент предоставлен и модерируется Biology Online Editors
4 1 – Knot Atlas
Непростое (составное) исполнение
Псевдокельтский орнаментальный узор с тремя узлами в виде восьмерки вдоль замкнутой треугольной петли.
Герб в окружении восьмерок
Узел презентаций
| Минимальный представитель оплетки | Презентация ссылки Морзе | Презентация дуги |
| Длина 4, ширина 3, Индекс косы 3 | [{3, 5}, {6, 4}, {5, 2}, {1, 3}, {2, 6}, {4, 1}] |
[редактировать Примечания к презентациям 4 1]
| Узел 4_1. | График, узел 4_1. |
Компьютерный разговор
Приведенные выше данные доступны в пакете Mathematica KnotTheory` . Ваш ввод (красный) реалистичен; все остальное должно иметь то же содержание, что и в реальном сеансе математики, но с другим форматированием.(Путь ниже может быть другим в вашей системе, и, возможно, также дата KnotTheory`)
In [1]: = | AppendTo [$ Path, "C: / drorbn / projects / KAtlas /"]; << KnotTheory` |
KnotTheory :: loading: Загрузка предварительно вычисленных данных в PD4Knots`. |
Выход [4] = | X 4251 X 8615 |
Out [5] = | 1, -4, 3, -1, 2, -3, 4, -2 |
(приведенный ниже путь может отличаться в вашей системе)
In [7]: = | AppendTo [$ Path, «C: / bin / LinKnot /»]; |
In [8]: = | ConwayNotation [K] |
KnotTheory :: credits: Минимальные косы, представляющие узлы с количеством пересечений до 10, были предоставлены Томасом Гиттингсом.См. ArXiv: math.GT/0401051. |
Выход [9] = |
In [10]: = | {Первый [br], Crossings [br], BraidIndex [K]} |
KnotTheory :: loading: Загрузка предварительно вычисленных данных в IndianaData`. |
In [11]: = | Показать [BraidPlot [br]] |
In [12]: = | Показать [DrawMorseLink [K]] |
KnotTheory :: кредиты: "MorseLink был добавлен в KnotTheory` Сиддартом Шанкараном из Университета Торонто летом 2005 года.« |
KnotTheory :: кредиты: «DrawMorseLink был написан Сиддартом Шанкараном в Университете Торонто летом 2005 года». |
In [13]: = | ap = ArcPresentation [K] |
Выход [13] = | ArcPresentation [{3, 5}, {6, 4}, {5, 2}, {1, 3}, {2, 6}, {4, 1}] |
Четырехмерные инварианты
Полиномиальные инварианты
Дополнительные квантовые инварианты
Дополнительные инварианты квантовых узлов для 4_1 .A1 Инварианты.
Инварианты A2.
| Масса | Инвариант |
|---|---|
| 1,0 | |
| 1,1 | |
| 2,0 | |
| 3,0 |
Инварианты A3.
| Масса | Инвариант |
|---|---|
| 0,1,0 | |
| 1,0,0 | |
| 1,0,1 |
Инварианты A4.
| Масса | Инвариант |
|---|---|
| 0,1,0,0 | |
| 1,0,0,0 |
B2 Инварианты.
| Масса | Инвариант |
|---|---|
| 0,1 | |
| 1,0 |
B3 Инварианты.
| Масса | Инвариант |
|---|---|
| 1,0,0 |
B4 Инварианты.
| Масса | Инвариант |
|---|---|
| 1,0,0,0 |
Инварианты C3.
| Масса | Инвариант |
|---|---|
| 1,0,0 |
Инварианты C4.
| Масса | Инвариант |
|---|---|
| 1,0,0,0 |
Инварианты D4.
| Масса | Инвариант |
|---|---|
| 0,1,0,0 | |
| 1,0,0,0 |
Инварианты G2.
| Масса | Инвариант |
|---|---|
| 1,0 |
.
«Подобные» узлы (в Атласе)
Тот же многочлен Александера / Конвея: {}
Тот же многочлен Джонса (с точностью до зеркального отображения): {K11n19,}
Компьютерный разговор
Приведенные выше данные доступны в пакете Mathematica KnotTheory` .Ваш ввод (красный) реалистичен; все остальное должно иметь то же содержание, что и в реальном сеансе математики, но с другим форматированием.(Путь ниже может быть другим в вашей системе, и, возможно, также дата KnotTheory`)
In [1]: = | AppendTo [$ Path, "C: / drorbn / projects / KAtlas /"]; << KnotTheory` |
In [4]: = | {A = Александр [K] [t], J = Jones [K] [q]} |
KnotTheory :: loading: Загрузка предварительно вычисленных данных в PD4Knots`. |
KnotTheory :: loading: Загрузка предварительно вычисленных данных в Jones4Knots`. |
Выход [4] = | {,} |
In [5]: = | DeleteCases [Выберите [AllKnots [], (A === Александр [#] [t]) &], K] |
KnotTheory :: loading: Загрузка предварительно вычисленных данных в DTCode4KnotsTo11`. |
KnotTheory :: кредиты: Преобразование GaussCode в PD было написано Сиддартом Шанкараном в Университете Торонто летом 2005 года. |
In [6]: = | DeleteCases [
Выбрать[
AllKnots [],
(J === Джонс [#] [q] || (J /. Q -> 1 / q) === Джонс [#] [q]) &
],
K
] |
KnotTheory :: loading: Загрузка предварительно вычисленных данных в Jones4Knots11`. |
Инварианты Васильева
| V 2,1 через V 6,9 : |
|
V 2,1 - V 6,9 предоставлены Петром Дуниным-Барковским
Цветные многочлены Джонса
Таблицы данных узлов
Таблицы данных узлов Перед использованием таблиц внимательно изучите обозначения и условности!После использования таблиц (в своей статье) , пожалуйста, поставьте ссылка на этот URL .Спасибо.
Также имейте в виду, что здесь много таблиц, несмотря на требующие кропотливого процесса, прошу независимую проверку. Таким образом, если вы обнаружите какие-либо несоответствия, сообщите мне немедленно. Точно так же сделайте, если вы сделали свой собственный проверьте, чтобы я мог поставить на него ссылку.
Обозначение
- Обозначения узлов таковы: Книга Рольфсена. Однако инварианты были вычислены с использованием KnotScape. KnotScape использует другую нумерацию! Вот как конвертировать KnotScape Рольфсену и Рольфсена для нумерации KnotScape.
- Узел отображается в зеркальном отображении, когда KnotScape рисует его. Таким образом, некоторые инварианты относятся к аверсам картин Рольфсена.
Вот как конвертировать
зеркалирование между обеими таблицами,
где узел записан как в таблице Рольфсена.
Например, Рольфсена 10 5 это KnotScape 10 56 , и в обеих таблицах
узел по-разному зеркальный. (Таким образом, лучше говоря,
Рольфсена 10 5 принадлежит KnotScape! 10 56 .)
Звездочка означает, что узел амфифиральный.Особая осторожность требуется с (Rolfsen's) 10 71 , поскольку он не является амфикхейральным, но здесь нет инварианта. (и вычисляемый с помощью KnotScape) может отличить его от его зеркальное изображение.
- Нумерация последних четырех 10 узлов пересечения Рольфсена была изменена. чтобы избежать дублирования Perko. Еще одна ошибка в таблицах Рольфсена состоит в том, что в них 10 83 и 10 86 были поменяны местами: Conway обозначение и многочлен Александера для каждого ссылались на диаграмму другого.Мы обмениваемся не диаграммы, а многочлен Александера и нотация Конвея исправить ошибку ( как таблица в новом 2003 издание книги Рольфсена, Дрор Атлас узлов Бар-Натана и Чак Таблица инвариантов узлов Ливингстона и в отличие от в Книга Каваути!).
- На этой странице по всей нумерации KnotScape, не чередуется узлы присоединяются после чередования одинаковых номер перехода. Так, например, Рольфсена 10 164 (А.k.a., 10 165 , до Perko's дублирование удалено) - это KnotScape's 10 161 , что означает (при наличии 123 чередующихся 10 узлов пересечения) Узел KnotScape 38 th , не чередующийся 10 пересечений.
- Подробнее о таблицах см. Также Вот!
Инварианты узлов
Вот таблицы Александр, Конвей, Джонс, Хомфлай, Брандт-Ликориш-Миллетт-Хо и Полиномы Кауфмана, инвариант Васильева степени 3 и подписи узлов до 10 переходов.- Бумажная форма таблиц содержит дальнейшие ссылки, подробности о соглашениях (с мотком отношения) и обозначения (с примерами, как это читать). Пожалуйста, проверьте это, если нижеперечисленные пункты оставляют вас неуверенными вы правильно понимаете таблицы.
- Обозначения многочленов являются комбинацией обозначений Ликориш-Миллетт и Адамс. Для Джонсов и Многочлен Александера, если абсолютный член находится между его минимальным и максимальных степеней, то она заключена в скобки, иначе минимальная степень равна записывается в фигурные скобки перед списком коэффициентов.Например, Джонс многочлен от 8 19 равен t 3 + t 5 -t 8 . Для полиномов Конвея и Брандта-Ликориша-Миллетта-Хо минимальный степень не указывается, так как она всегда равна нулю. Для Полином Конвея, коэффициенты нечетной (нулевой) степени опускаются.
- Для полинома ХОМФЛИ мы опускаем элементы l k m s с одним из k, s нечетным (так как они равны нулю). Для полиномиальных одночленов Кауфмана a k z s только для четности k + s.В другие коэффициенты опускаются или для k = 0 заменяются звездочкой чтобы прояснить степени a. Наше соглашение для полиномов ХОМФЛИ и Кауфмана отличается от этого Ликориш-Миллетт соотв. Кауфман, заменив l с l -1 и a с -1 .
- Затем подпись заносится в таблицу в соответствии с соглашением о том, что положительный (справа) трилистник имеет подпись 2. (Положительные ссылки имеют положительная подпись !)
- Наконец, приведем значения инварианта Васильева степени 3 v 3 в его асимметричном (изменение знака при зеркалировании) аддитивная (по связанной сумме) версия.Нормализация такова, что v 3 = 1 на положительном трилистнике. Это соглашение Поляк-Виро. (Нормализация Фидлера отличается в 4 раза).
Мутации
До 13 переходов
Вот список всех мутантных групп (91 группа, состоящая из 86 пары и 5 троек) простых узлов через 12, и 13 переходов (774 группы, состоящие 703 пары, 38 троек, 32 группы по 4 человека и одна группа из 6 человек).Чтобы найти мутантов, я сначала определил группы узлов равного объема, многочлены Александера и Джонса полином.Тогда мутации явно проявились в диаграммы минимальных пересечений.
Не обращайте внимания на зеркальное отображение узлов!
Список групп через 12 переходов подтверждает, по крайней мере, по длине, компиляция, сделанная незадолго до этого Дэвидом Де Витом, см. Вот.
14 переходов
Вот список из 14 мутантов скрещивания группы (без кодов DT): всего 4435 групп, состоящих из 3917 пар, 233 тройных, 262 четверных, 17 групп по 6 человек и 6 групп по 8 узлов.Список был определен с помощью вышеуказанного типа проверки. Осталось более 7 пар:
(14 41721 , 14 42125 )
(14 41739 , 14 42126 )
(14 41763 , 14 42021 )
(14 42947 , 14 6 43476 42953 , 14 43572 )
(14 43904 , 14 46158 )
(14 43907 , 14 46187 )
Узлы в первой и третьей паре различаются по Полином Уайтхеда с двойным мотком.Вторая пара была самой сложно сломать. Даниэль Матей отличил это абелианизацией ядер гомоморфизмов или низкоиндексных подгрупп группы основная группа 2-кратно разветвленной крышки. Последние 4 пары являются мутантами, но отображают мутацию только на 15 диаграммах скрещивания.
15 переходов
Вот список из 15 скрещенных мутантов группы (без кодов DT): всего 29049 групп, состоящих из 24884 пары, 1000 троек, 2909 четверных, 172 группы по 6 человек и 84 группы по 8 узлов.78 пар (отмеченных звездочкой) отображают мутацию. только в 16 схемах скрещивания и 6 пар только в 17 схемах скрещивания (отмечены восклицательным знаком). Еще 34 пары с равный объем, многочлен Александера и многочлен Джонса. 14 из они различаются полиномом двойного мотка Уайтхеда, а остальные 20 - с помощью вышеупомянутых теоретико-групповых тестов.
Зеркальное отображение
Поскольку я игнорировал различие между узлами и зеркальными изображениями, Я не перечислял хиральные узлы, которые являются мутантами по своим зеркальные изображения.Таких узлов не бывает до 13, а до 15 переходов, но есть 13 узлов из 14 переходов. Они есть Вот.Представители тесьмы и индекс тесьмы
Вот таблица индексов кос простые не чередующиеся узлы через 13 пересечений.(MWF резок для чередования узлов через 17 переходов.) Звездочка (*) означает, что только 2cMWF резкий,
знак амперсанда (&) означает, что только 3cMWF является резким.
Вот таблицы косички представителей прайма узлов через 12 переходов.
- Представители кос имеют минимальную (= индекс косы) ширину,
- если до 18 переездов, из (самых коротких) минимальная длина косы из минимальная (ширина) тесьма представителей. (Обратите внимание, что узлы не могут иметь минимальную ширину тесьмы представители минимальной длины или наоборот, например 10 136 !)
- Обозначения для узлов через 10 пересечений взяты из книги Рольфсена: для 11 и 12 переходов это KnotScape; дублирование Perko и ошибка 10 83 и 10 86 обрабатываются как сказано выше.
- Зеркальное отражение узла не требуется!
- Коса представители 12 1316 , Исправлены 12 1319 и 12 1417 после комментария Чака Ливингстона.
Вот список основных
узлы 14-16 пересечений индекса косы
2,3. Вот непеременных
индекса оплетки 4,
с узлами 16 1059154 и 16 1153788 не определились
(они могут, но вряд ли присоединятся).
Индекс плетения 4 определяется MWF для чередующихся узлов.
(глянь сюда).
Роды / Канонические роды
В этой таблице показаны роды (и канонические роды) не чередующихся простых 11 и 12 узлов пересечения, и 13 пересекающихся не чередующихся узлов.- До 10 скрещиваний и для чередования сучков рода и канонические роды такие же, и задаются степенью Полином Александера.
- Для всех узлов до 13 пересечений канонический род дается половиной степень переменной Александера в полиноме мотка (Неравенство Мортона точное).
- Если степень полинома Александера совпадает с половиной степень переменной Александера в полиноме мотка, затем род приравнивается к каноническому роду; имеется 49 безопеременных 11 и 12 узлов пересечения и 287 не меняющихся 13 узлов пересечения, где это не тот случай.
- Из 49 узлов 11 и 12 пересечений 33 все еще имеют минимальный род. канонические поверхности, доказанные дисковым разложением Габая. так же сделать 176 из 287 не чередующихся 13 узлов.Эти узлы отмечен звездочкой.
- Остальные 16 узлов 11 и 12 переходов и 111 узлов 13 переходы отмечены восклицательным знаком.
- Остальные 11 и 12 узлов скрещивания имеют род 2 и канонический род 3. Минимальные родовые поверхности могут быть найдены методом Рудольфа. конструкция плетеной поверхности. Минимальность следует из Многочлен Александера (степени 2), за исключением 3 узлов.
- Осталось 3 узла: 11 409 (узел K-T), 12 1311 и 12 1412 .Известно, что первый узел имеет диск рода два. разложимая поверхность Габая. Плетеные поверхности Genus 2 для другие были найдены (стабильно) водопроводным эквивалентом Хопфа, а род 2 плетеной поверхности на 12 1412 найдено диск-кольцо разложимы по Миками Хирасаве, поэтому эти узлы также имеют род 2. Позже я заметил, что они также следуют (гораздо легче) из результат Кобаяши, так как у них есть незаузлованный номер 1.
- 111 узлов 13 переходов подвергаются той же процедуре.Сначала мы находим скрученные поверхности, которые являются кандидатами в минимальный род. Затем мы используем полином Александера, результат Кобаяши (который говорит этот род ≥2, когда число незаузлованных = 1), и сантехника Хопфа тест на эквивалентность. Последнее означает, что мы пытались связать плетеный Поверхности Хопфа (де) переходя к поверхностям минимального рода других узлов мы знаем. Это решило 106 узлов.
- Остальные 5 узлов пересечения 13 5764 , 13 5826 , 13 5841 , 13 5897 и 13 8765 .От них я мог только ожидать, что верхняя род является точным, и я смог установить, что спроектированный род = 3 узла (13 5764 , 13 8765 ) - хопф эквивалент сантехники (так что род = 3 для одного улаживает другой).
- Эти 5 узлов были позже рассчитаны Стефаном Фридлом, Натан Данфилд, Николас Джексон и Стефано Видусси используют витой Многочлены Александера. Это единственное доказательство, которое я им пока знаю. (Фридл и Тэхи Ким также подтвердили пересечение узлов 11 и 12 для которых многочлен Александера не дает рода.)
- Наша таблица для 11 узлов пересечения подтверждает выполненный расчет. ранее Джейк Расмуссен, используя узел Озсват-Сабо Floer гомология.
Волокно- Известно, что 10 или менее пересекающихся узлов являются волокнистыми.
если (и только если) многочлен Александера моничен.
- Это было проверено Миками Хирасавой, что
11 пересекающихся узлов расслоены, если (и только если) узел Александра
полином является моническим и степени, соответствующей роду.
(См. K-T узел и узел Конвея имеют тривиальный многочлен, и
11 440 имеет монический полином меньшей степени.)
- Расслоенность 12 пересекающихся узлов определяется следующим образом.
Тринадцать 12 перекрестных узлов с моническим многочленом Александера степени
соответствующие роду не расслоены. Фридл и Ким рассчитали это по формуле
скрученные полиномы Александера (см.
здесь), а позже
Сообщается, что Джейк Расмуссен провел независимую проверку, используя
Узел Озсвата-Сабо гомологии Флоера. (Если у вас другой
проверка, пожалуйста, сообщите мне. Используя методы Габая, я пока
проверил 8 узлов, в которых поверхности минимального рода каноничны.)
Эти узлы отмечены знаком доллара. Другие узлы с моником
Многочлены Александера степени, соответствующей роду, расслоены.
- Теперь также решена проблема расслоенности для 13 пересекающихся узлов.
Всего 2759 узлов с моническим многочленом Александера степени = род.
И род> 1. (Волокнистые сучки 1-го рода - это только трилистник и
фигура-8-узел.) Из них 78 узлов не волокнистые. Эти узлы
помечены знаком доллара. Остальные 2681 узел с моникой
Многочлены Александера степени, соответствующей роду, расслоены.В
решение (кроме 15 узлов) использует дисковое разложение Габая
канонические поверхности (когда канонические поверхности минимального рода существуют)
и тест на эквивалентность сантехники более простым узлам (когда канонические
поверхностей минимального рода не существует). Это дает результат кроме
на 15 узлов последнего типа. Я не мог решить это сам и
проконсультировался у С. Фридла, который сообщил о них без волокон после его
расчет с Dunfield / Jackson / Vidussi. Видимо другой
узлы - это , которые еще не прошли полную независимую проверку .
Генераторы рода
Вот список образующих рода 2 (24 узла),
род 3 (4017 узлов) и (в формате gzip) род 4,
разделены между специальными (1480 238 узлов, 22 МБ)
и неспециальные (1 934 581 узел, 25 МБ), шт.
Значение «генераторы» описано в моей статье.
"Узлы (канонического) рода два" на моем список документов.
Вкратце это означает, что генераторы «генерируют»
все диаграммы простых узлов этого (канонического) рода с помощью мух, пересекая
изменения и полная версия твиста.( Примечание: Идентификаторы узлов не имеют особого значения,
и никакого отношения к существующим таблицам узлов; они только что пришли
техническими процедурами, использованными для создания списков.)
Амфикейральные узлы
Вот список простых амфихиральных узлов до 16 переходов.Александр Стойменов,
stoimeno_stoimenov.net
Отношения ManyToMany> Коллекции доктрин: ManyToMany, формы и другие сложные отношения
-
В этом курсе
-
Все SymfonyCasts
- Учебники
- Стоимость
- Войти
- Зарегистрироваться
ДОРОГА
Symfony 3 >
КУРС
Коллекции доктрины: ManyToMany, формы и другие сложные отношения
Купить доступ к курсу Скачать Код курса Это видео Сценарий курсаFrontiers | Палеофитогеографические закономерности на границе перми и триаса
Введение
Переход от перми к триасу был отмечен серьезными нарушениями в морских и наземных экосистемах.Что наиболее важно, массовое вымирание в конце пермского периода стало самым серьезным событием вымирания морских животных (например, Benton and Twitchett, 2003; Stanley, 2016; Delfino et al., 2020). Сроки и влияние этого события на биоразнообразие наземных животных и наземных растений еще не установлены (например, Benton and Newell, 2014; Cascales-Miñana and Cleal, 2014, Cascales-Miñana et al., 2016; Fielding et al. , 2019; Nowak et al., 2019; Gastaldo et al., 2020; Mays et al., 2020). Однако общепринято считать, что структура сообществ наземных флор претерпела важные изменения, и растет согласие с тем, что эти изменения следует рассматривать в первую очередь как вопрос последовательных экологических нарушений, а не как вопрос резкой потери биоразнообразия (Hochuli et al. ., 2016; Мишра и др., 2017; Филдинг и др., 2019; Новак и др., 2019; Вайда и др., 2020).
Палеофитогеографические аспекты этих изменений в глобальном масштабе в основном изучались путем признания различных фитохорий (или фитопровинций) - фитогеографических единиц, которые определяются по наличию общих таксонов и, следовательно, стратиграфически ограничены ареалом этих таксонов ( напр., Вахрамеев и др., 1978). Хотя географическое распространение определяющих растений, безусловно, зависело от условий окружающей среды, фитохории разных времен не обязательно отражают эквивалентную среду.Напротив, биомы определяются как районы со сравнимыми, контролируемыми климатом сообществами растений и животных (Walter, 1985). Концепция биома успешно применялась к окаменелостям растений вместе с данными об отложениях, чувствительных к климату с древних времен, включая сакмарский (цисуральский / ранняя пермь; Rees et al., 2002), вордский / кунгурский (гваделупский / средний пермский период; Ziegler) , 1990; Rees et al., 1999a; Rees et al., 1999b; Rees et al., 2002; Willis, McElwain, 2014) и юрский период (Rees, Ziegler, 1996; Rees et al., 2000; Уиллис и МакЭлвейн, 2014). Биомы триаса и юры Евразии были представлены Ziegler et al. (1993). Глобальные карты биомов от раннего до среднего триаса еще не доступны. Климатические модели использовались для оценки распределения биомов в Сакмарском (Rees et al., 2002), Вордском / Кунгурском (Kutzbach and Ziegler, 1993; Rees et al., 1999a; Rees et al., 1999b; Rees et al. al., 2002), чансин (лопинги / поздняя пермь; Fluteau et al., 2001; Roscher et al., 2011) и поздний триас (Sellwood, Valdes, 2006).Данные по споровым пыльцам также использовались для палеофитогеографии, включая рассмотрение климатических условий, на что указывают палинологические комплексы в раннем триасе (Ярошенко, 1997) и в конце среднего – начале позднего триаса (Visscher and Van der Zwan, 1981). Чумаков и Жарков (2002) и Чумаков и Жарков (2003) использовали палино- и макрофлору вместе с литологическими, палеогеографическими и палеоклиматическими данными для реконструкции климатических поясов для ранней и поздней перми и раннего триаса.Палеофитогеографическое распространение и климатические последствия макрофлоры южной Гондваны также исследовались Кунео (1996) для перми и Артабе и др. (2003) и Spalletti et al. (2003) для триаса.
Мы представляем здесь поэтапный анализ распределения биомов, основанный на сообществах макрофоссилий наземных растений от учьяпинга до ладинца, в попытке отследить глобальные климатические сдвиги с влиянием на растительные сообщества. Это исследование также показывает, как состав биомов меняется со временем и как неоднородность летописи окаменелостей растений влияет на неопределенность в отношении границ между биомами.
Материалы и методы
Источники данных и выборка
Это исследование основано на подмножестве данных Nowak et al. (2019), большая часть которых была взята из более ранних публикаций (Дополнительные данные; см. Также дополнительные данные 5 и дополнительные ссылки Nowak et al., 2019). Наше подмножество представляет собой 8 892 проявления макрофоссилий наземных растений от Wuchiapingian до ладинского яруса, датированных с разумной достоверностью и с достаточно точной информацией о географическом положении изученных обнажений (дополнительная таблица S1).По сравнению с исходным набором данных мы внесли многочисленные исправления в отношении таксономических синонимов и данных о местонахождении. Встречи с неопределенной идентификацией на уровне рода не учитывались. Кроме того, местам были присвоены координаты широта / долгота.
Представленные здесь палеогеографические карты соответствуют преобразованию глобальных тектонических реконструкций плит в трехмерные топографические поверхности (палеотопография) или палео-ЦМР (цифровые модели рельефа). Конверсия проистекает из работы Vérard et al.(2015a) и Vérard et al. (2015b). Тектоническая модель плит соответствует первой версии модели Паналезиса (все еще разрабатываемой), которая описана в работах Верара (2019a) и Верара (2019b). Заменяя модель UNIL, разработанную в Лозаннском университете (например, Stampfli and Borel, 2002; Hochard, 2008), модель Panalesis создается с нуля, но основана на тех же методах и ноу-хау. Определение синтетических изохрон, в частности, помогает реконструировать не только континентальные области, но и океанические области, в том числе исчезнувшие в результате процессов субдукции, такие как Панталассический океан.Глобальные реконструкции являются результатом итерационных процессов, в которых тектонические плиты (с закрытыми границами плит и 100% покрытой земной поверхностью) перемещаются в соответствии с геологическими данными широкого типа, геометрией плит, кинематикой плит и геодинамическими сценариями, определенными в трех измерениях и время (см., например, рис. 2.5.11-13 в Hochard, 2008).
Климатические данные по осадкам от поздней перми до среднего триаса были получены из Boucot et al. (2013); см. дополнительные рисунки S1 – S3.
Расчеты и интерпретация
Современные координаты местонахождений окаменелостей растений и данные об осадках были переведены в палеокоординаты поздней перми и среднего триаса в соответствии с моделью Паналезиса (Vérard, 2019a; Vérard, 2019b). Региональное таксономическое разнообразие рассчитывается по стадиям для биомов (Таблица 1), палео-широтных срезов 10 ° (Таблица 2) и основных палеогеографических регионов (Таблица 3; Рисунок 1) в виде разнообразия выборки в ячейках на уровне рода, т. Е. Количества роды, о которых сообщалось на каждой стадии (не считая неопределенных определений).Эндемизм (таблица 4) рассчитывается как соотношение родов растений только в одном крупном палеогеографическом регионе на определенной стадии. Случаи, когда один и тот же род встречается в другом регионе на другой стадии, игнорируются.
ТАБЛИЦА 1 . Число местонахождений ископаемых растений и таксономическое разнообразие на уровне родов по стадиям и биомам.
ТАБЛИЦА 2 . Число местонахождений ископаемых растений и таксономическое разнообразие на уровне родов на один этап и 10 ° широтный срез.
ТАБЛИЦА 3 .Число местонахождений ископаемых растений и таксономическое разнообразие на уровне родов по стадиям и палеогеографическим регионам.
РИСУНОК 1 . Палеогеографическая карта поздней перми – раннего триаса (250 млн лет) с указанием основных регионов.
ТАБЛИЦА 4 . Число местонахождений окаменелостей растений и таксономическое разнообразие на уровне родов на стадии, включая число и частоту эндемичных родов.
Основываясь на известных экологических ограничениях и распределении таксонов растений в течение рассматриваемого временного интервала, мы определили, в каких биомах отдельные роды растений, как ожидается, будут встречаться обычно или исключительно (дополнительная таблица S2).Используемые здесь биомы соответствуют десяти биомам Зиглера (1990), хотя не все из них применимы (см. Ниже). Для каждой комбинации местность-стадия в наборе данных мы вычислили наиболее вероятный биом, сначала объединив вхождения из одновозрастных местонахождений в пределах движущегося окна с радиусом 100 км (рассчитано по палеогеографическим координатам в предположении сферической Земли со средним радиусом 6371.009 км), чтобы учесть краткосрочную изменчивость, не полностью зарегистрированные сообщества и фактически эквивалентные местонахождения, о которых сообщалось с разным уровнем географической точности.На основе объединенных вхождений и вышеупомянутых назначений биомов было создано взвешенное голосование: каждое вхождение рода в пуле подсчитывается с весом, равным обратной величине количества биомов, с которыми он связан. Присвоение биома считается вероятным, если на него указывает> 50% взвешенных голосов за местность и этап. Если присутствует хотя бы один род, который считается исключительным элементом определенного биома, то соответствующая коллекция также с уверенностью назначается этому биому.В случае коллекций, в которых расчет не указывает на какой-либо биом с достаточной степенью уверенности, биом назначается вручную на основе таксономического состава, географического положения и сходства с соседними сообществами. Микросреды, возникающие на местном уровне из-за воздействия близлежащей береговой линии, дельтовых условий и / или изменений уровня моря (например, Kustatscher et al., 2017), не принимаются во внимание. Назначенные биомы были спроектированы на палеогеографических картах для каждого этапа, и мы добавили предварительную экстраполяцию областей биома с учетом палеотопографии и климатических данных по осадкам (Рисунки 2–7).
РИСУНОК 2 . Местонахождение макроскопических ископаемых растений учьяпинги с соответствующими биомами и предполагаемые площади расширения биомов, спроецированные на палеогеографическую реконструкцию для поздней перми – раннего триаса (250 млн лет назад). Пунктирные символы обозначают рассчитанные биомы, пустые символы обозначают ручные биомы.
РИСУНОК 3 . Местонахождения макрофоссилий растений Чансин (Changhsingian) с соответствующими биомами и предполагаемые площади расширения биомов, спроецированные на палеогеографическую реконструкцию для поздней перми – раннего триаса (250 млн лет назад).Пунктирные символы обозначают рассчитанные биомы, пустые символы обозначают ручные биомы.
РИСУНОК 4 . Местонахождения макрофоссилий индийских растений с соответствующими биомами и предполагаемые площади расширения биомов, спроецированные на палеогеографическую реконструкцию для поздней перми – раннего триаса (250 млн лет назад). Пунктирные символы обозначают рассчитанные биомы, пустые символы обозначают ручные биомы.
РИСУНОК 5 . Местонахождение макрофоссилий оленекских растений с ассоциированными биомами и предполагаемые площади расширения биомов, спроецированные на палеогеографическую реконструкцию для поздней перми – раннего триаса (250 млн лет назад).Пунктирные символы обозначают рассчитанные биомы, пустые символы обозначают ручные биомы.
РИСУНОК 6 . Местоположение макрофоссилий анисовых растений с соответствующими биомами и предполагаемые площади расширения биомов, спроецированные на палеогеографическую реконструкцию для среднего триаса (240 млн лет назад). Пунктирные символы обозначают рассчитанные биомы, пустые символы обозначают ручные биомы.
РИСУНОК 7 . Ладинские местонахождения макрофоссилий растений с ассоциированными биомами и предполагаемые площади расширения биомов, спроецированные на палеогеографическую реконструкцию для среднего триаса (240 млн лет назад).Пунктирные символы обозначают рассчитанные биомы, пустые символы обозначают ручные биомы.
Биомы
Следующие десять биомов были определены Зиглером (1990) и служат основой для нашего анализа:
Тропический вечно влажный биом (биом 1: экваториальный и вечнозеленый тропический биом Зиглера, 1990) представляет собой вечнозеленые дождевые леса в тропики, которые развиваются в постоянно жарких (средняя T = 26–29 ° C) и влажных условиях (10–20 мм / месяц) и, как правило, представляют собой наиболее таксономически разнообразный биом.Типичными для этого биома являются обильные многоэтажные леса с высокими деревьями и лианами. Леса улучшают постоянные условия, задерживая воздух, удерживая влагу и создавая влагу за счет непрерывного испарения. Биом также включает торфяные почвы / угольные болота и характерные для них растительные сообщества. Это наиболее распространено вблизи экватора, но может встречаться и в средних широтах. Растительные сообщества, произрастающие в этих условиях окружающей среды, обладают высоким природоохранным потенциалом.
Тропический летний и влажный биом (биом 2: тропический и субтропический летне-влажный биом Циглера, 1990) включает в себя множество растительных сообществ, от лиственных лесов до саванн с разной степенью лесов и пастбищ, все из которых становятся продуктивными в течение влажные летние месяцы в низких и средних широтах с сильной сезонностью.Они являются переходным звеном между тропическим вечно влажным биомом и субтропическим пустынным биомом. Хотя эти флоры являются продуктивными и дифференцированными, их потенциал сохранения ограничен из-за потенциального окисления почв в засушливые сезоны.
Субтропический пустынный биом (биом 3: прибрежный и внутренний биом тропической пустыни Циглера, 1990) характеризуется нехваткой воды в течение каждого месяца в году. Растительность, как правило, скудная, а растения, которые встречаются, приспособлены к выживанию, а не к конкуренции.В растительности преобладают ксерофиты и суккуленты, которые могут сохраняться в постоянно засушливой среде. Крупные плоские корневые системы предназначены для сбора влаги; размеры листьев уменьшены, чтобы избежать высыхания. Потенциал сохранности этих растений исключительно низкий. Осадочные индикаторы - эвапориты и эоловые пески.
Зимний биом (биом 4 Циглера, 1990) относится к склерофилловым вечнозеленым и обычно невысоким деревьям с кожистыми листьями с большим разнообразием.Зимняя влажность в этом биоме уникальна и связана сегодня с развитием полярного фронта и связанных с ним внетропических циклонов. Продуктивность биомассы и потенциал сохранения этой растительности низкие. Этот биом отнесен к теплому умеренному биому для данного исследования, потому что мы обнаружили, что невозможно различить два с точки зрения таксономического состава.
Биом умеренно теплого пояса (биом 5: западный и восточный биом умеренно теплого климата Циглера, 1990) относится к вечнозеленым лесам умеренного пояса, подверженным сезонным изменениям климата.Сегодня осадки хорошо распределяются в течение всего года благодаря чередованию зимних фронтальных систем и летних конвективных систем. Суточный минимум температуры самых холодных месяцев выше нуля (в отличие от биома с прохладным умеренным климатом), но растительность проводит зиму в состоянии покоя. Продуктивность составляет половину тропического вечнозеленого биома, но разнообразие в целом очень велико. Местами могут развиваться торфяные болота.
Биом с прохладным умеренным климатом (биом 6: западный и восточный биом с прохладным умеренным климатом Циглера, 1990) отмечает лиственные леса, подверженные сильным морозам и холодной зиме.Лиственным деревьям требуется вегетационный период не менее четырех месяцев со средней дневной температурой выше 10 ° C, чтобы у них было достаточно времени для образования листьев. В этом биоме может происходить ежегодное изменение сообщества из-за плотного полога летом и голых деревьев в другое время года.
Пустынный биом в средних широтах (биом 7 Циглера, 1990) включает пустыни и степи с холодными зимами. Сегодня в этом биоме преобладают высокотравные или низкотравные прерии из-за пустынной среды и ежегодного дефицита воды.Продуктивность биомассы от умеренной до низкой, потенциал сохранения очень низкий. Флоры, указывающие на такие степи, не могли быть выделены в нашем наборе данных, но можно предположить, что некоторые из обширных территорий без комплексов ископаемых растений в умеренном поясе соответствуют этому биому. На рисунках, представленных здесь, этот биом был объединен с биомом субтропической пустыни, так как границы между ними были бы чисто умозрительными.
Холодно-умеренный биом (биом 8 Циглера, 1990) в настоящее время характеризуется бореальными хвойными лесами и тайгами (широко расставленные деревья с почвенным покровом из мхов и лишайников).Сегодня он отсутствует в Южном полушарии, но хорошо развит в Северном полушарии. Осадки от слабых до умеренных, но вода не является ограничивающим фактором из-за низкой скорости испарения и короткого вегетационного периода. Определяющей чертой является короткий вегетационный период, который зависит от температуры и солнечного света, а не от осадков.
Арктический биом (биом 9 Циглера, 1990) представляет собой безлесную тундру с крохотной флорой трав и небольших кустарников из-за сокращенного вегетационного периода (также может составлять всего один месяц в современных эквивалентах).Современные тундры состоят из злаков, мхов, лишайников и некоторых карликовых кустарников. Этот биом не удалось идентифицировать в нашем наборе данных из-за отсутствия характерных таксонов.
Ледниковый биом (биом 10 Циглера, 1990) является в основном абиотическим и может быть обозначен отложениями, такими как тиллиты и дропстоуны. Поскольку он лишен растений, он игнорируется в нашем анализе, но следует отметить, что ледниковые отложения (дропстоуны и глендониты) были зарегистрированы в поздней перми в Антарктиде и Австралии (Boucot et al., 2013, и ссылки там). Они встречаются вблизи или даже на более низких широтах, чем сообщества позднепермских растений, и, следовательно, не служат признаком крупномасштабного оледенения.
Результаты
Учьяпингиан
Учьяпингиан представлены в этом исследовании 125 изученными участками, в которых в общей сложности насчитывается 150 родов растений (Таблица 4). Это единственный этап в исследуемом интервале, для которого по имеющимся палеофлорным данным выделяются шесть различных биомов (рис. 2).Он отмечен широким распространением тропического вечнозеленого биома, который присутствует в Северном и Южном Китае, Турции и на некоторых террейнах Тетии (Индокитай, Шан, Цянтянь). Основываясь на этом распределении и учитывая сходство с ранее описанными моделями биома для средней перми (Ziegler, 1990; Rees et al., 1999a; Rees et al., 1999b, Rees et al., 2002), а также климатическими моделями для поздней перми (Roscher et al., 2011) кажется вероятным, что тропический вечно влажный биом также охватывал другие районы вдоль экваториального и северного побережья Тетиса, а также экваториального восточного побережья Пангеи.Это самый разнообразный биом в учьяпинге, включающий 114 родов (17 порядков) в Северном полушарии и 24 рода (11 порядков) в Южном полушарии (таблица 1; дополнительная таблица S3). Общими элементами тропического вечно влажного биома являются Lepidodendrales (6 родов) среди ликофитов, Equisetales (11 родов) и Sphenophyllales (4 рода) среди сфенофитов, Marattiales (11 родов) и Osmundale
Frontiers | Молекулярные сигнатуры и филогеномный анализ рода Burkholderia: предложение о разделении этого рода на измененный род Burkholderia, содержащий патогенные организмы, и новый род Paraburkholderia gen.ноя обитатели окружающей среды
Введение
Род Burkholderia представляет собой морфологически, метаболически и экологически разнообразную группу грамотрицательных бактерий (Yabuuchi et al., 1992; Coenye and Vandamme, 2003; Mahenthiralingam et al., 2005; Palleroni, 2005; Compant et al. , 2008). Burkholderia видов повсеместно распространены в окружающей среде (Coenye and Vandamme, 2003). Они населяют широкий спектр экологических ниш, от почвы до дыхательных путей человека (Coenye and Vandamme, 2003).Группа из 17 близкородственных видов Burkholderia , комплекса Burkholderia cepacia (BCC), ответственна за распространенные и потенциально летальные легочные инфекции у лиц с ослабленным иммунитетом, таких как люди с муковисцидозом (Mahenthiralingam et al., 2002, 2005; Biddick et al., 2003; Hauser et al., 2011). Burkholderia pseudomallei , вид Burkholderia , связанный с BCC, является возбудителем мелиоидоза, потенциально смертельной септической инфекции, на долю которой приходится до 20% всех внебольничных септицемий в некоторых регионах (White, 2003; Лимматуротсакул и Павлин, 2011).Другие виды, связанные с BCC, являются возбудителями основных инфекций как у животных ( Burkholderia mallei ), так и у растений ( Burkholderia glumae и Burkholderia gladioli ) (Whitlock et al., 2007; Nandakumar et al., 2009) .
Несмотря на большое разнообразие и различную патогенность среди> 70 членов группы, все вида Burkholderia в настоящее время относятся к одному роду (Coenye and Vandamme, 2003; Palleroni, 2005). Филогения и таксономия рода Burkholderia в первую очередь определяется на основе анализа последовательности 16S рРНК (Yabuuchi et al., 1992; Паллерони, 2005; Ярза и др., 2008). Выводы, полученные в результате анализа 16S рРНК, были дополнительно подтверждены другими филогенетическими методами, включая анализ на основе гена recA (Payne et al., 2005), анализ на основе гена acdS (Onofre-Lemus et al., 2009), ДНК –Гибридизация ДНК (Gillis et al., 1995), анализ жирных кислот целых клеток (Stead, 1992), анализ мультилокусных последовательностей (Tayeb et al., 2008; Spilker et al., 2009; Estrada-de los Santos et al., 2013), анализ прироста / потери генов (Zhu et al., 2011) и филогенетический анализ полногенома (Ussery et al., 2009; Segata et al., 2013). Во многих из этих филогенетических исследований представители рода Burkholderia можно разделить на две или несколько отдельных филогенетических групп, при этом одна группа состоит из представителей BCC и родственных видов (Payne et al., 2005; Tayeb et al. , 2008; Yarza et al., 2008; Spilker et al., 2009; Ussery et al., 2009; Gyaneshwar et al., 2011; Vandamme, Dawyndt, 2011; Zhu et al., 2011; Estrada-de los Santos et al. al., 2013; Segata et al., 2013). Хотя есть некоторые общие черты среди близкородственных групп видов Burkholderia , нет никаких известных морфологических, биохимических или молекулярных характеристик, специфичных для более крупных филогенетических групп внутри рода (например, BCC и родственных видов).
Появление методов секвенирования следующего поколения привело к быстрому увеличению числа последовательностей генома, доступных для видов бактерий (Mardis, 2008). Доступность этих последовательностей для представителей рода Burkholderia дает нам лучшие средства для оценки филогенетических отношений между различными видами (Ciccarelli et al., 2006; Wu et al., 2009). Важно отметить, что большие наборы данных последовательностей позволяют использовать сравнительные геномные методы для обнаружения новых молекулярных маркеров, которые могут предоставить независимые доказательства для различных филогенетических групп в пределах рода Burkholderia (Gupta, 1998, 2014; Gao and Gupta, 2012). В этой работе мы описываем один тип молекулярного маркера, вставки или делеции консервативных последовательностей (CSI), которые однозначно присутствуют в белковых последовательностях из определенной группы организмов, которые могут быть использованы для выделения различных филогенетических групп видов Burkholderia независимо друг от друга. традиционных филогенетических методов (Gupta, 1998, 2001; Gao, Gupta, 2012).Наш сравнительный анализ геномов Burkholderia привел к идентификации 42 уникальных CSI, которые очерчивают различные филогенетические группы внутри рода в ясных молекулярных терминах. Клада Burkholderia , содержащая BCC и родственные организмы (Clade I), была подтверждена как филогенетическими данными, так и 6 идентифицированными CSI. Мы также идентифицировали 3 CSI, специфичных для BCC, 4 CSI, специфичных для группы B. pseudomallei , и 5 CSI, специфичных для патогенных растений Burkholderia spp.Остальные представители рода Burkholderia сформировали еще одну монофилетическую кладу (Clade II) на наших филогенетических деревьях, которая поддерживалась двумя CSI. Внутри Clade II мы идентифицировали две меньшие клады Burkholderia , которые поддерживались 16 и 6 CSI. Группирование представителей рода Burkholderia по крайней мере в две большие монофилетические группы также наблюдалось в большом количестве предшествующих филогенетических исследований (Payne et al., 2005; Tayeb et al., 2008; Yarza et al., 2008; Spilker et al., 2009; Ussery et al., 2009; Гианешвар и др., 2011; Чжу и др., 2011; Эстрада-де-лос-Сантос и др., 2013 г .; Segata et al., 2013). Основываясь на филогенетических данных и идентифицированных нами CSI, мы предлагаем разделить род Burkholderia на два рода: измененный род Burkholderia , содержащий клинически важные и фитопатогенные представители рода, и новый род Paraburkholderia gen. ноя укрывательство экологических видов.
Материалы и методы
Филогенетический анализ
Конкатенированное выравнивание последовательностей 21 высококонсервативного белка (т.е.ArgRS, EF-G, GyrA, GyrB, Hsp60, Hsp70, IleRS, RecA, RpoB, RpoC, SecY, ThrRS, TrpS, UvrD, ValRS, 50S рибосомные белки L1, L5 и L6 и 30S рибосомные белки S2, S8 и S11 ) был использован для филогенетического анализа. Из-за их присутствия в большинстве бактерий эти белки широко используются для филогенетических исследований (Gupta, 1998, 2009; Kyrpides et al., 1999; Harris et al., 2003; Charlebois and Doolittle, 2004; Ciccarelli et al., 2006). ). Аминокислотные последовательности этих консервативных белков были получены из базы данных NCBI для всех видов / штаммов, перечисленных в Таблице 1, которая включает 45 секвенированных видов рода Burkholderia .Кроме того, из NCBI были извлечены три генома других представителей класса Betaproteobacteria (а именно Cupriavidus necator N-1, Bordetella pertussis Tohama I и Neisseria meningitides MC58), которые служили в качестве внешних групп в нашем анализе. база данных. В зависимости от наличия генома были отобраны типовые штаммы для большинства видов. Множественные выравнивания последовательностей для этих белков были созданы с использованием Clustal_X 1.83 и объединены в один файл выравнивания (Jeanmougin et al., 1998). Плохо выровненные регионы из файла выравнивания были удалены с использованием Gblocks 0.91b, и полученное выравнивание, которое содержало 7688 выровненных символов, в конечном итоге было использовано для филогенетического анализа (Castresana, 2000). Дерево максимального правдоподобия (ML), основанное на 100 повторностях бутстрапов этого выравнивания, было построено с использованием MEGA 6.0 при использовании модели замены Jones-Taylor-Thornton (Jones et al., 1992; Tamura et al., 2013).
Таблица 1. Геномные характеристики секвенированных представителей рода Burkholderia .
Консенсусное дерево последовательности гена 16S рРНК с максимальной вероятностью было также создано для 101 последовательности, которая включала 97 репрезентативных штаммов из рода Burkholderia и четыре внегрупповые последовательности из родов Cupriadivus и Ralstonia . Последовательности, использованные в исследовании, были получены из проекта Ribosomal Database Project (RDP III) (Cole et al., 2009) и NCBI. Все последовательности были выровнены с использованием MAAFT 7 (Katoh and Standley, 2013), и дерево ML, основанное на 1000 повторностях начальной загрузки этого выравнивания, было построено с использованием General Time Reversible Model (Tavaré, 1986) в MEGA 6.0 (Тамура и др., 2013).
Идентификация молекулярных маркеров (CSI)
Было проведенопоисков BLASTp для всех белков из хромосом 2 и 3 (номера доступа NC_008061 и NC_008061) Burkholderia cenocepacia J2315 (Holden et al., 2009) для выявления CSI, которые являются общими для разных представителей рода Burkholderia . Были отобраны виды, которые оказались наиболее популярными с высокими показателями гомологов (значения E <1e −20 ) из рода Burkholderia и других внешних групп.Множественные выравнивания последовательностей были созданы с использованием Clustal_X 1.83 (Jeanmougin et al., 1998). Эти выравнивания визуально проверяли на наличие вставок или делеций (инделей), ограниченных одним или всеми членами рода Burkholderia и фланкированными по крайней мере 5-6 консервативными аминокислотными остатками с обеих сторон в соседних 30-40 аминокислотах. кислоты. Запросы Indel, которые не сопровождались консервативными областями, в дальнейшем не оценивались. Видовая специфичность запросов indel, удовлетворяющих вышеуказанному критерию, была дополнительно оценена путем выполнения поиска BLASTp на коротких сегментах последовательности, содержащих вставки или делеции, и их фланкирующих консервативных областях (длиной 60–100 аминокислот).Поиск проводился в базе данных NCBI non-redundant (nr), и минимум 250 совпадений BLAST были проверены на наличие или отсутствие CSI. Результаты этих анализов были оценены, как подробно описано в нашей недавней работе (Gupta, 2014). Файлы сигнатур для CSI, которые были специфичны для представителей рода Burkholderia , были созданы и отформатированы с использованием программ SIG_CREATE и SIG_STYLE (доступных на Gleans.net), как описано Gupta (2014). Представленные здесь файлы выравнивания последовательностей содержат информацию для всех обнаруженных вставок или делеций из интересующей группы Burkholderia , но только ограниченное количество из видов, которые служат внешними группами.Информация о последовательностях для разных штаммов разных видов не показана, но все они демонстрировали сходную картину. Наконец, если не указано иное, показанные здесь CSI конкретно находятся в указанных группах, и аналогичные CSI не были обнаружены в 250 попаданиях Blast с запросными последовательностями.
Результаты
Схема ветвления видов Burkholderia в деревьях конкатенированных белков и 16S рРНК
Последовательности генома 45 видов Burkholderia были доступны из базы данных генома NCBI на момент написания этой работы (NCBI, 2014).Некоторые характеристики этих геномов перечислены в Таблице 1. Размеры геномов секвенированных видов Burkholderia сильно различаются (от 3,75–11,29 МБ), и количество белков в них также варьируется в аналогичной пропорции. В этой работе мы создали филогенетическое дерево ML на основе конкатенированных аминокислотных последовательностей 21 консервативного домашнего и рибосомного белков, полученных из 45 секвенированных видов Burkholderia (рис. 1). Виды Burkholderia сформировали две большие клады в дереве ML на основе белков: одну, состоящую из BCC и родственных организмов (Clade I), и другую, состоящую в основном из экологических или плохо охарактеризованных видов Burkholderia (Clade II).Внутри клады I также наблюдаются три отчетливые клады меньшего размера. Первая из этих клад (Clade Ia) полностью состоит из секвенированных видов BCC, вторая клада (Clade Ib) - группы B. pseudomallei и близкородственных видов, а третья клада (Clade Ic) состоит из патогенных видов растений. , B. glumae и B. gladioli . Кладу II можно также разделить на две меньшие клады, Кладу IIa и Кладу IIb. Clade IIa отделен от Clade IIb длинной ветвью, что позволяет предположить, что между двумя группами произошло большое генетическое расхождение.Помимо двух основных клад Burkholderia , два вида - Burkholderia sp. JPY347 и Burkholderia rhizoxinica , рано разветвленные на дереве и не ассоциированные ни с Clade I, ни с II.
Рис. 1. Филогенетическое дерево максимального правдоподобия секвенированных геномов представителей рода Burkholderia на основе конкатенированных последовательностей 21 консервативного белка . Для укоренения дерева использовали Cupriavidus necator N-1, Bordetella pertussis Tohama I и Neisseria meningitides MC58.Результаты анализа начальной загрузки указаны для каждого узла. Основные клады Burkholderia (Clades I и II) и их основные подклассы указаны в скобках.
Мы также построили филогенетическое дерево ML на основе 16S рРНК для 97 штаммов Burkholderia и видов-кандидатов (рис. 2). В этом филогенетическом дереве на основе 16S рРНК мы наблюдали в целом сходные паттерны с нашей филогенетикой, основанной на белках. Клада, состоящая из BCC и родственных организмов (Clade I), была четко разделена.Три субклада в Clade I, BCC (Clade Ia), группа B. pseudomallei (Clade Ib) и патогенные виды растений (Clade Ic) были хорошо разделены, хотя некоторые виды демонстрировали аберрантное ветвление (например, B). . oklahomensis и B. pseudomultivorans ). Большое скопление оставшихся видов Burkholderia , примерно соответствующих кладу II в нашем филогенетическом дереве на основе конкатенированных белков, также наблюдалось в дереве 16S рРНК. Однако из-за значительного количества несеквенированных видов Burkholderia , которые присутствуют в базе данных 16S рРНК, сложно точно идентифицировать группы внутри Clade II дерева 16S рРНК, которые соответствуют Clades IIa и IIb в нашем филогенетическом дереве на основе конкатенированных белков. .Поддержка начальной загрузки для ветвей в дереве на основе 16S рРНК также была значительно ниже, чем в конкатенированном дереве белков, что указывает на то, что некоторые из наблюдаемых паттернов ветвления могут быть ненадежными. Однако клады, состоящие из BCC и родственных организмов (Clade I), имеют сильную поддержку бутстрапа и были идентифицированы в большом количестве предыдущих филогенетических исследований на основе 16S рРНК (Yabuuchi et al., 1992; Palleroni, 2005; Yarza et al. , 2008; Суарес-Морено и др., 2012).
Рисунок 2.Дерево максимального правдоподобия на основе последовательностей гена 16S рРНК 97 представителей рода Burkholderia . Номера доступа для секвенированной 16S рРНК, используемой для каждого организма, указаны в скобках после названия организма. Дерево было укоренено с использованием четырех видов из родов Cupriadivus и Ralstonia . Результаты анализа начальной загрузки указаны для каждого узла. Основные клады Burkholderia (Clades I и II) и субклады внутри Clade I обозначены скобками.
Молекулярные сигнатуры, отличающие кладу I и кладу II Burkholderia
Редкие генетические изменения, такие как вставки и делеции в основных генах / белках, которые происходят у общего предка, могут быть унаследованы различными видами умерших, связанных с этим общим предком (Gupta, 1998; Rokas and Holland, 2000; Gogarten et al. , 2002; Гупта, Гриффитс, 2002). Из-за редкости и специфического присутствия этих редких генетических изменений в родственной группе организмов они могут служить важными молекулярными маркерами и предоставлять новые средства для понимания эволюционных взаимосвязей между различными близкородственными видами (Gupta, 1998; Gupta and Griffiths , 2002; Гао, Гупта, 2012).
Сравнительный анализ белковых последовательностей видов Burkholderia , проведенный в настоящей работе, выявил ряд CSI, которые служат для четкого различения ряда различных клад в пределах рода Burkholderia . Эти исследования привели к идентификации 6 CSI, специфичных для Clade I Burkholderia , состоящих из BCC и родственных организмов, что позволило четко отличить эту группу от всех других Burkholderia .Эта клада, которая содержит все хорошо охарактеризованные патогены в пределах рода, представляет собой наиболее клинически значимую группу в пределах Burkholderia . Все виды этой клады потенциально патогенны для человека, животных или растений, и большинство из них было выделено из клинических образцов человека (Simpson et al., 1994; Mahenthiralingam et al., 2002, 2005; Biddick et al., 2003; O ' Carroll et al., 2003). Один из примеров CSI, специфичного для Clade I Burkholderia , показан на рисунке 3A.В этом случае делеция одной аминокислоты присутствует в высококонсервативной области периплазматического белка, связывающего аминокислоты. Индель с обеих сторон фланкирован высококонсервативными участками, что указывает на то, что это не результат артефактов выравнивания, а надежная генетическая характеристика. Этот CSI присутствует во всех секвенированных членах Clade I Burkholderia , но отсутствует во всех других бактериальных гомологах этого белка. Наша работа идентифицировала 5 дополнительных CSI в др. Широко распространенных белках, которые специфичны для Clade I Burkholderia , и выравнивание последовательностей для этих CSIs показано на Supplemental Figures 1–5, а сводка их характеристик представлена в Таблице 2.
Фиг. 3. Частичное выравнивание последовательностей (A) периплазматического белка, связывающего аминокислоты, показывающее делецию 1 аминокислоты, идентифицированную у всех членов Clade I рода Burkholderia (B) дегидрогеназы, показывающей вставку 1 аминокислоты ( в рамке), идентифицированные только у представителей Clade II рода Burkholderia . Эти CSI не были обнаружены в гомологах последовательностей этих белков из каких-либо других секвенированных бактерий. В каждом случае показана информация о последовательности для видов Burkholderia и ограниченного числа других бактерий, но, если не указано иное, аналогичные CSI были обнаружены у всех членов указанной группы и не обнаружены ни у каких других видов бактерий в топ-250 BLAST. хиты.Штрих (-) в выравнивании указывает идентичность с остатком в верхней последовательности. Идентификационные номера GenBank (GI) для каждой последовательности указаны во втором столбце. Информация о последовательностях для других CSI, специфичных для представителей Clade I и Clade II рода Burkholderia , представлена на дополнительных рисунках 1–5 и дополнительных рисунках 6 соответственно, а их характеристики приведены в таблице 2.
Таблица 2. Сохраненные индели сигнатур, специфичные для двух основных клад в пределах рода Burkholderia .
Две дополнительные CSI, идентифицированные в этой работе, специфичны для вида Clade II Burkholderia , который состоит в основном из организмов окружающей среды. Один из этих CSI, показанный на рисунке 3B, состоит из вставки одной аминокислоты в дегидрогеназный белок, который уникально обнаружен у представителей Clade II Burkholderia и отсутствует у всех других видов Burkholderia , а также всех других бактериальных групп. Выравнивание последовательности для другого CSI, специфичного для Clade II Burkholderia (делеция 2 аминокислот в семействе LysR белка-регулятора транскрипции) показано на дополнительном рисунке 6, а его характеристики суммированы в таблице 2.
CSI, различающих различные основные группы внутри Clade I Burkholderia
Виды из клады I рода Burkholderia ответственны за ряд болезней человека, животных и растений (Biddick et al., 2003; Mahenthiralingam et al., 2005). Члены клады I (то есть BCC и родственная Burkholderia ) обычно делятся на 3 основные группы, которые соответствуют кладам, идентифицированным на наших филогенетических деревьях. Первая группа, члены BCC (Clade 1a), являются преобладающими патогенами у пациентов с муковисцидозом, вторая группа - B.pseudomallei (Clade Ib), содержит возбудителей мелиоидоза и сапа, а в третью группу входят патогенные растения Burkholderia видов (Clade Ic) (White, 2003; Mahenthiralingam et al., 2005; Whitlock et al., 2007; Нандакумар и др., 2009). Наш анализ выявил 3 CSI, которые специфичны для всех членов клады BCC (Clade 1a). Один пример CSI, специфичного для клады BCC, показан на фиг. 4A. Этот CSI состоит из вставки из 2 аминокислот в консервативную область репрессора утилизации гистидина, который обнаруживается только у членов BCC.Выравнивания последовательностей для двух других CSI, специфичных для клады BCC, показаны на дополнительных рисунках 7, 8, а их характеристики приведены в таблице 3.
Фигура 4. Частичное выравнивание последовательностей (A) репрессора утилизации гистидина, показывающее вставку из 2 аминокислот (выделена рамкой), идентифицированную во всех членах комплекса Burkholderia cepacia (Clade Ia) в пределах рода Burkholderia (B) a периплазматический олигопептид-связывающий белок, показывающий вставку из 1 аминокислоты (в рамке), идентифицированную во всех членах группы Burkholderia pseudomallei (Clade Ib) в пределах рода Burkholderia (C) SMP-30 / глюконолаконаза / LRE-подобная область - содержащий белок, показывающий вставку из 1 аминокислоты (заключен в рамку), идентифицированный у всех представителей фитопатогенной клады Burkholderia (Clade Ic) .Эти CSI не были обнаружены в гомологах последовательностей этих белков из каких-либо других секвенированных бактерий в 250 лучших совпадениях BLAST. Информация о последовательностях для других CSI, специфичных для субкладов внутри клады I рода Burkholderia , представлена на дополнительных рисунках 7-15, а их характеристики суммированы в таблице 3.
Таблица 3. Сохраненные индели сигнатур, специфичные для групп внутри Clades I и II .
Наша работа также определила 4 CSI, характерных для B.pseudomallei (Clade Ib), которая содержит наиболее распространенный человеческий патоген в пределах рода, B. pseudomallei (Wiersinga et al., 2006). Один пример CSI, специфичного для группы B. pseudomallei , который состоит из вставки из 1 аминокислоты в консервативной области периплазматического олигопептид-связывающего белка, показан на фиг. 4B. Выравнивания последовательностей для трех других CSI в трех разных белках, которые специфичны для группы B. pseudomallei , показаны на дополнительных рисунках 9-11, а их характеристики суммированы в таблице 3.
Мы также идентифицировали 5 CSI, которые специфичны для основной патогенной группы растений в пределах рода Burkholderia (Clade 1c), который содержит виды B. glumae и B. gladioli . Пример CSI, представляющий эту группу, показан на рисунке 4C. Этот CSI состоит из вставки из 1 аминокислоты в консервативную область белка, содержащего SMP-30 / глюконолаконазу / LRE-подобную область, который обнаружен у представителей Clade 1c рода Burkholderia , но отсутствует у всех других Burkholderia и все другие группы бактерий.Выравнивание последовательностей для других 4 CSI показано на дополнительных рисунках 12–15, а их ключевые особенности выделены в таблице 3.
CSI, специфичных для двух групп внутри Clade II Burkholderia
Вид внутри клады II из рода Burkholderia населяет множество экологических ниш, но существует мало свидетельств их колонизации здоровыми или ослабленными иммунитетами пациентов (Coenye and Vandamme, 2003). Ветвление различных групп внутри клады II плохо разрешено в деревьях 16S рРНК, и в настоящее время не хватает данных о последовательностях, которые можно использовать для создания деревьев на основе конкатенированных наборов генов, которые надежно разрешают взаимосвязи клады, в то же время в достаточной степени отражая общую разнообразие видов внутри клады (рисунки 1, 2) (Cole et al., 2009; NCBI, 2014). Несмотря на ограниченные данные о последовательностях, мы смогли идентифицировать две устойчивые группы в Clade II, которые поддерживаются рядом CSI. Первый Clade, Clade IIa, в основном состоит из неклассифицированных представителей рода Candida и видов Burkholderia (рис. 1). Clade IIa поддерживается 16 CSI, идентифицированными в этой работе. Один из примеров CSI, специфичного для Clade IIa, состоящего из вставки из 1 аминокислоты в 3-фосфоглицератдегидрогеназу, показан на Фигуре 5A.Эта вставка присутствует в высококонсервативной области этого белка у всех секвенированных членов Clade IIa и отсутствует во всех других Burkholderia и всех других бактериальных группах. Выравнивание последовательностей для других 15 CSI, специфичных для Clade IIa Burkholderia spp. показаны на дополнительных рисунках 16–30, а их характеристики приведены в таблице 3.
Фигура 5. Частичное выравнивание последовательностей (A) 3-фосфоглицератдегидрогеназы, показывающее вставку из 1 аминокислоты (в рамке), идентифицированную у всех представителей Clade IIa рода Burkholderia (B) 4-гидроксиацетофенонмонооксигеназа, показывающая 1 аминокислоту вставка (в рамке) идентифицирована только у представителей Clade IIb рода Burkholderia .Эти CSI не были обнаружены в гомологах последовательностей этих белков из каких-либо других секвенированных бактерий в 250 лучших совпадениях BLAST. Информация о последовательностях для других CSI, специфичных для субкладов внутри Clade II рода Burkholderia , представлена на дополнительных рисунках 16–35, а их характеристики суммированы в таблице 3.
Вторая группа в пределах Clade II Burkholderia (Clade IIb) состоит из большого разнообразия экологических видов Burkholderia (Coenye and Vandamme, 2003; Suarez-Moreno et al., 2012). Наш анализ выявил 6 CSI, специфичных для этой большой группы из видов Burkholderia . Один из примеров CSI, специфичных для представителей Clade IIb рода Burkholderia , показан на Фигуре 5B. CSI состоит из вставки одной аминокислоты в 4-гидроксиацетофенонмонооксигеназу, которая присутствует только у представителей Clade IIb рода Burkholderia , но не в белковых гомологах из любой другой секвенированной бактериальной группы. Информация для других 5 CSI, специфичных для представителей Clade IIb рода Burkholderia , показана на дополнительных рисунках 31–35, а их характеристики суммированы в таблице 3.
Обсуждение
Род Burkholderia - одна из крупнейших групп видов в классе Betaproteobacteria (Palleroni, 2005; Parte, 2013). Этот род содержит множество бактерий, которые населяют широкий спектр экологических ниш, включая ряд бактерий, обладающих патогенным потенциалом (Yabuuchi et al., 1992; Coenye and Vandamme, 2003; Mahenthiralingam et al., 2005; Palleroni, 2005; Compant и др., 2008). Филогения рода Burkholderia была изучена с использованием широкого набора методологий, основанных на фенотипических, биохимических, генетических и геномных характеристиках (Stead, 1992; Gillis et al., 1995; Пейн и др., 2005; Тайеб и др., 2008; Онофре-Лемус и др., 2009; Spilker et al., 2009; Ussery et al., 2009; Гианешвар и др., 2011; Вандамм и Давиндт, 2011; Чжу и др., 2011; Эстрада-де-лос-Сантос и др., 2013). Эти исследования позволили по-новому взглянуть на эволюционные взаимоотношения видов в пределах рода Burkholderia . Однако до настоящего времени не было внесено никаких таксономических изменений из-за отсутствия отдельных отличительных характеристик, идентифицированных для различных филогенетических линий внутри рода (Estrada-de los Santos et al., 2013).
В настоящей работе мы выделили две основные группы видов в пределах рода Burkholderia : кладу I, которая содержит всех патогенных членов рода, и кладу II, которая содержит большое разнообразие видов, обитающих в окружающей среде. Было обнаружено, что эти две группы отчетливо разветвляются в филогенетическом дереве с высоким разрешением, основанном на большом количестве конкатенированных белковых последовательностей, полученных в этой работе (Рисунок 1). Доказательства отличия организмов клады I от других видов Burkholderia наблюдались в широком диапазоне предыдущих филогенетических исследований (Payne et al., 2005; Тайеб и др., 2008; Ярза и др., 2008; Spilker et al., 2009; Ussery et al., 2009; Гианешвар и др., 2011; Вандамм и Давиндт, 2011; Чжу и др., 2011; Суарес-Морено и др., 2012; Эстрада-де-лос-Сантос и др., 2013 г .; Segata et al., 2013). Важно отметить, что мы также идентифицировали 6 и 2 CSI, которые служат отдельными молекулярными характеристиками Clade I и Clade II, соответственно (Рисунок 6 и Таблица 2). Эти CSI являются первыми дискретными признаками, которые были идентифицированы и уникальны для Clade I или Clade II рода Burkholderia .Эти CSI действуют как независимая проверка филогенетических тенденций, выявленных в этом и других исследованиях, и предоставляют четкие доказательства того, что виды из клады I отличаются от всех других Burkholderia и происходят от общего предка, за исключением всех остальных Burkholderia. . Хотя информация о последовательностях членов Clade II в настоящее время несколько ограничена, основываясь на общем присутствии ими двух CSI, вполне вероятно, что они также происходят от общего предка, за исключением других бактерий.
Рис. 6. Сводная диаграмма, изображающая распределение идентифицированных CSI и предлагаемых названий двух основных групп (Clade I и II) в пределах Burkholderia . Основные клады Burkholderia обозначены скобками и выделены.
Кроме того, мы идентифицировали молекулярные свидетельства в виде большого количества CSI, которые подтверждают различимость нескольких меньших групп в пределах рода Burkholderia .Наиболее важные из этих групп, комплекс B. cepacia (BCC; Clade Ia) и группа B. pseudomallei (Clade Ib), поддерживаются 3 и 4 идентифицированными CSI соответственно. BCC - это группа условно-патогенных микроорганизмов, которые колонизируют иммунодефицитных человеческих хозяев и являются одними из наиболее распространенных и смертельных инфекций у пациентов с муковисцидозом (Mahenthiralingam et al., 2002, 2005; Biddick et al., 2003; Hauser et al., 2011) . 17 видов, которые составляют BCC, тесно связаны и образуют плотный монофилетический кластер в пределах рода Burkholderia (Vandamme and Dawyndt, 2011).Группа B. pseudomallei состоит из 4 близкородственных видов: B. pseudomallei , возбудителя высоколетального мелиоидоза сепсиса (White, 2003; Limmathurotsakul and Peacock, 2011), B. mallei , возбудителя. сапа лошадей и случайных инфекций человека (Whitlock et al., 2007), а также в основном непатогенных организмов, Burkholderia thailandensis и Burkholderia oklahomensis (Deshazer, 2007).Выявленные CSI являются высокоспецифичными характеристиками этих двух важных патогенных групп и представляют собой новые и полезные мишени для разработки диагностических тестов либо для BCC, либо для группы B. pseudomallei (Ahmod et al., 2011; Wong et al. , 2014). Мы определили CSI для трех других групп в пределах рода Burkholderia : группа патогенных растений Burkholderia , родственных BCC, и группа B. pseudomallei (Clade Ic), группа, содержащая безымянные и кандидатные Burkholderia виды ( Clade IIa) и группа, состоящая из экологической Burkholderia (Clade IIb).Мы выделили 6, 16 и 6 CSI для этих трех групп соответственно. Эти CSIs обеспечивают важные отличительные характеристики для этих групп, особенно для Clades IIa и IIb, которые являются родственными группами, у которых нет других идентифицированных отличительных характеристик (Suarez-Moreno et al., 2012).
Филогенетический анализ, идентифицированные CSI и патогенные характеристики различных видов Burkholderia , представленные в этой работе, убедительно свидетельствуют о том, что род Burkholderia состоит как минимум из двух различных линий.Одна линия состоит из BCC и родственных организмов (Clade I), а другая - из широкого круга организмов окружающей среды (Clade II). Эта последняя клада филогенетически очень разнообразна, и информация о ее последовательностях ограничена. Таким образом, возможно, что в будущем эта последняя клада может быть обнаружена как состоящая из более чем одной отдельной бактериальной линии, однако в настоящее время ясно, что клада I и клада II представляют разные линии. Доказательства отличия представителей Clade I от других видов Burkholderia были выявлены в ряде предыдущих филогенетических исследований (Payne et al., 2005; Тайеб и др., 2008; Ярза и др., 2008; Spilker et al., 2009; Ussery et al., 2009; Гианешвар и др., 2011; Вандамм и Давиндт, 2011; Чжу и др., 2011; Суарес-Морено и др., 2012; Эстрада-де-лос-Сантос и др., 2013 г .; Segata et al., 2013). Эстрада-де-лос-Сантос и др. (2013) недавно завершили филогенетический анализ рода Burkholderia , используя мультилокусный анализ последовательностей генов atpD, gltB, lepA и recA в сочетании с геном 16S рРНК, что дает убедительные доказательства наличия двух отдельные эволюционные линии в пределах рода Burkholderia .Однако эти авторы воздержались от формального предложения разделения рода на два рода из-за нехватки дифференцирующих признаков для этих двух групп. Наш сравнительный анализ геномов Burkholderia выявил набор отличительных молекулярных характеристик, которые четко различают две эволюционные линии в пределах рода Burkholderia в дополнение к филогенетическим данным. В свете обилия филогенетических и молекулярных доказательств наличия двух различных эволюционных линий в пределах рода Burkholderia и различных профилей патогенности представителей этих двух групп, мы предлагаем, чтобы род Burkholderia был разделен на два отдельных рода.Первый из этих монофилетических родов, который включает все клинически значимые виды и четко отличается от всех других видов Burkholderia , сохранит название Burkholderia (Clade I). Остальным из видов Burkholderia (Clade II), которые включают широкий спектр экологических видов, мы предлагаем название Paraburkholderia gen. ноя Исправленное описание рода Burkholderia и описание Paraburkholderia gen.ноя представлены ниже. Краткое описание сочетаний новых видов в пределах Paraburkholderia gen. ноя представлены в таблице 4.
Таблица 4. Описание новых комбинаций рода Paraburkholderia gen. ноя .
Измененное описание рода Burkholderia (Yabuuchi et al., 1993 Emend. Gillis et al., 1995)
Род включает типовой вид B. cepacia (Yabuuchi et al., 1993). Виды этого рода представляют собой грамотрицательные, прямые или слегка изогнутые палочки, подвижность которых опосредована одним или несколькими полярными жгутиками. Только у B. mallei жгутики отсутствуют и он неподвижен. У этого вида нет влагалищ или протезов, и они не проходят стадий покоя. Большинство видов способны накапливать и использовать поли-β-гидроксибутират (ПОБ) для роста. Эти виды в основном являются аэробными хемоорганотрофами, но некоторые виды способны к анаэробному дыханию, используя нитрат в качестве конечного акцептора электронов.Содержание G + C у представителей рода колеблется от 65,7 до 68,5%. Члены рода образуют отличную монофилетическую кладу на филогенетических деревьях, и они отличаются от всех других бактерий консервативными последовательностями, описанными в этой работе в следующих белках: периплазматический белок, связывающий аминокислоты, 4-гидроксибензоат 3-монооксигеназа, 6-фосфоглюконатдегидрогеназа, субъединица альфа саркозиноксидазы, предполагаемый липопротеин и предполагаемая лиаза (таблица 2).
Описание рода Paraburkholderia Gen.Ноябрь
В род входит типовой вид Paraburkholderia graminis comb. ноя (Basonym: Burkholderia graminis , Viallard et al., 1998) Виды этого рода представляют собой грамотрицательные прямые или слегка изогнутые палочки с одним или несколькими полярными жгутиками. Остальные морфологические и метаболические характеристики сходны с родом Burkholderia . Содержание G + C у представителей рода колеблется от 61,4 до 65,0%. Вид не связан с людьми.Члены этого рода обычно образуют отдельную кладу по соседству с родом Burkholderia на филогенетических деревьях, и у них отсутствуют молекулярные сигнатуры, специфичные для Burkholderia . Большинство секвенированных членов этого рода содержат индели консервативных последовательностей, описанные в этой работе, в белковых последовательностях безымянной дегидрогеназы и регулятора транскрипции семейства LysR (Таблица 2).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: http://www.frontiersin.org/journal/10.3389/fgene.2014.00429/abstract
Ссылки
Achouak, W., Christen, R., Barakat, M., Martel, M.-H., and Heulin, T. (1999). Burkholderia caribensis sp. nov., бактерия-продуцент экзополисахарида, выделенная из микроагрегатов вертисола на Мартинике. Внутр. J. Syst. Бактериол . 49, 787–794.DOI: 10.1099 / 00207713-49-2-787
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ахмод, Н. З., Гупта, Р. С., и Шах, Х. Н. (2011). Идентификация специфической индели Bacillus anthracis в гене yeaC и разработка быстрого анализа пиросеквенирования для отличия B. anthracis от группы B. cereus . J. Microbiol. Методы 87, 278–285. DOI: 10.1016 / j.mimet.2011.08.015
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Айзава, Т., Ве, Н. Б., Накадзима, М., и Сунаири, М. (2010a). Burkholderia heleia sp. nov., азотфиксирующая бактерия, выделенная из водного растения, Eleocharis dulcis , которое растет в сильнокислых болотах на территориях с кислыми сульфатными почвами во Вьетнаме. Внутр. J. Syst. Evol. Microbiol . 60, 1152–1157. DOI: 10.1099 / ijs.0.015198-0
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Айзава, Т., Ве, Н. Б., Виджарнсорн, П., Накадзима, М., и Сунаири, М.(2010b). Burkholderia acidipaludis sp. nov., устойчивые к алюминию бактерии, выделенные из водяного каштана китайского ( Eleocharis dulcis ), произрастающего в сильно кислых болотах Юго-Восточной Азии. Внутр. J. Syst. Evol. Microbiol . 60, 2036–2041. DOI: 10.1099 / ijs.0.018283-0
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Айзава Т., Виджарнсорн П., Накадзима М. и Сунаири М. (2011). Burkholderia bannensis sp.nov., кислотонейтрализующая бактерия, выделенная из торпедной травы ( Panicum repens ), произрастающей в сильно кислых болотах. Внутр. J. Syst. Evol. Microbiol . 61, 1645–1650. DOI: 10.1099 / ijs.0.026278-0
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Биддик, Р., Спилкер, Т., Мартин, А., и ЛиПума, Дж. Дж. (2003). Доказательства передачи Burkholderia cepacia, Burkholderia multivorans и Burkholderia dolosa среди лиц с муковисцидозом. FEMS Microbiol. Lett . 228, 57–62. DOI: 10.1016 / S0378-1097 (03) 00724-9
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бремер, К. О., Вандамм, П., да Силва, Л. Ф., Гомес, Дж., И Стейнбюхель, А. (2001). Полигидроксиалканоат-накапливающая бактерия, выделенная из почвы плантации сахарного тростника в Бразилии. Внутр. J. Syst. Evol. Microbiol . 51, 1709–1713. DOI: 10.1099 / 00207713-51-5-1709
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст |
.