Состав чисел от 1 до 10 домики: Домики «Состав числа» от 1 до 10, от 11 до 20 — РОСТОВСКИЙ ЦЕНТР ПОМОЩИ ДЕТЯМ № 7
Состав чисел от 1 до 10. IV. Новый материал. Онлайн игры «Числовые домики».
Тема: Число 10. Состав числа 10.
Тип урока: Ознакомление с новым материалом.
- Познакомить с числом 10 и его составом.
- Учить операциям сложения и вычитания в пределах 10 с опорой на его состав.
- Закрепить знания составов чисел до 9, действий в пределах 9.
- Учить самостоятельности и умению слушать других.
- Развивать речь, память, внимание и логическое мышление.
I. Орг. момент.
Этот ученик назовет номер и поприветствует следующего ученика и так далее по кругу. Затем дайте студентам бумагу и цветные карандаши и попросите их работать парами, чтобы проследить обе руки с вытянутыми пальцами, а затем наметить трассировку. Позвоните 11 волонтерам и назначьте каждому номер между 0 и попросите их сделать цифровую карту для своего назначенного номера. Собирайте и перемешайте цифровые карты.
Развивающие онлайн игры по математике «Состав числа — лампочки»
Затем вытащите соединительные кубы, а затем покажите ученикам цифровую карту.
Прозвенел звонок, начинается урок.
II. Актуализация знаний.
Ребята, посмотрите на доску. В домиках № 7 и № 9 некоторые жильцы получили странное послание. Хотите узнать какое?
Тогда в тетрадях запишите этих жильцов – пропущенные числа в парах состава чисел 7 и 9, а на доске работают двое.
Сейчас расположите числа в порядке убывания, и в соответствии с буквенным кодом прочитайте им послание. Что у вас получилось?
IV. Новый материал
Спросите их, сколько историй в башне. Повторите с другими цифрами 0 – 10 в случайном порядке. Когда они могут с комфортом сделать башни для заданных чисел, покажите две карточки с цифрами, например, 6 и попросите учащихся сделать две башни и сравнить их. Затем попросите учащихся поделиться своими сравнениями с классом с использованием описательной лексики.
Попросите учащихся посмотреть цифровую карту, которую вы указали. Просмотрите со студентами, как сделать цифру. Вернитесь к классу, чтобы вы писались в той же ориентации, что и студенты. Затем проследите фигуру в воздухе большими ударами. Поощряйте учащихся делать это с вами.
Что такое клад?
Хотите найти математический клад?
Какими качествами должны обладать кладоискатели?
А кто ещё всегда ищет клады?
Откроем учебник на стр. 35 № 8. Посмотрите, пираты нас хотят опередить, они уже на острове. Давайте узнаем, найдут ли они клад, для этого вычислим примеры по стрелкам. Что у вас получилось?
Что заметили? Верно, ответ предыдущего примера становится началом следующего. Такие примеры называются “круговыми”. Ответ последнего должен быть началом всей цепочки:
Вы можете найти рекомендации по составлению справочника ресурсов для написания нумерации для студентов, которые испытывают трудности с написанием цифр. В качестве записи на этот урок или в качестве записи в своем учебном портфолио, студенты кладут две башни на лист бумаги, прослеживают вокруг каждой башни и записывают цифры для каждого из них.
Затем дайте каждому учащемуся копию Деятельного листа Десяти кадров. Попросите учащихся смоделировать каждый номер, который вы показываете, поставив один счетчик на раздел в десяти кадрах, начиная с лица смайлика и перемещаясь в направлении стрелки, пока верхняя строка не будет заполнена.
Где можно было ещё замкнуть круг?
Это нам позволит обогнать пиратов. Пусть они ходят по кругу, а мы пойдём вперёд к цели. Вот и нужное место. Надо копать.
III. Физкультминутка.
Копаем землю и её выбрасываем их ямы (имитация движений под счёт до 9).
IV. Новый материал.
Ну, вот и заветный сундук, но на нём замок. На замке числа. Какие, узнаем, выполнив математический диктант. Все пишут в тетради, а на двое доске. Внимание:
Отобразите цифровую карту и наблюдайте за учениками, когда они размещают соединительные кубы. Попросите их удалить кубы, прежде чем моделировать следующий номер. В десяти кадрах используется концепция контрольных чисел и помогает учащимся создавать визуальные изображения для каждого номера. Например, это устройство позволяет легко видеть, что 6 составляет 1 больше 5, а 6 на 4 меньше, чем.
Например, это устройство позволяет легко видеть, что 6 составляет 1 больше 5, а 6 на 4 меньше, чем.
Увлекательное и эффективное обучение счету
Вы можете пожелать, чтобы они записывали каждое число, когда они моделируют его в десяти кадрах.
Студенты также могут использовать инструмент «Десять кадров» для изучения номеров до этого. Этот интерактивный инструмент можно использовать на мобильных устройствах.
Запишите число, следующее за 7 (8)
Запишите число, предыдущее 5 (4)
Запишите число, стоящее между 6 и 8 (7)
Запишите соседей числа 2 (1 и 3)
Запишите число, стоящее справа от 8 (9)
Запишите число, стоящее слева от 6 (5)
Я задумала число, оно предшествует 3 (2)
Елена Илларьевна задумала число, оно следует за 5 (6)
Проверяем (взаимопроверка) работу на доске.
Наконец, раздайте каждому ученику три палочки, белый клей и пинто-бобы. Попросите учащихся приклеить 10 бобов на каждую палочку, разделив их поровну в две группы.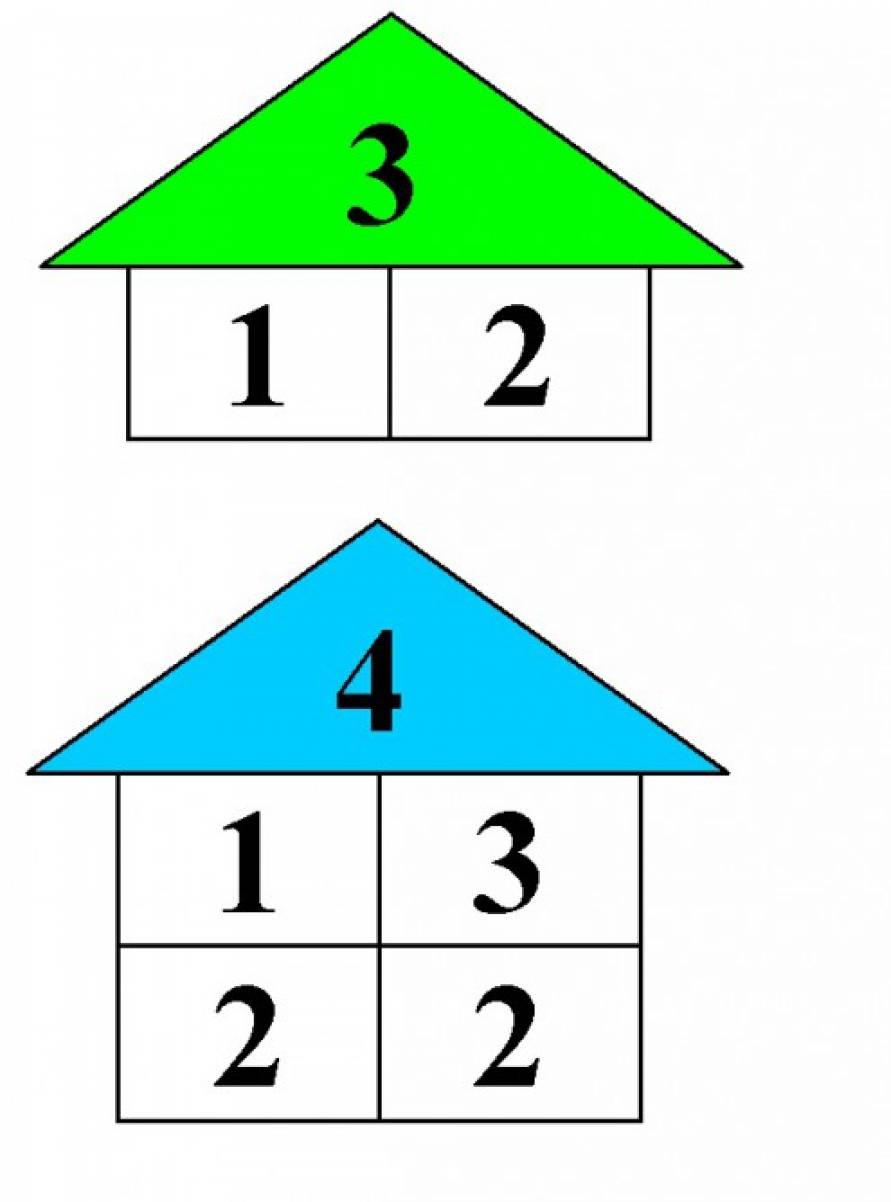 Изобретатель бобовых палочек, Боб Виртц, предложил это размещение, чтобы ученики знали о соотношении чисел к эталонам 5, и он использовал этот же принцип, когда он изобрел десять кадров.
Изобретатель бобовых палочек, Боб Виртц, предложил это размещение, чтобы ученики знали о соотношении чисел к эталонам 5, и он использовал этот же принцип, когда он изобрел десять кадров.
Попросите учащихся сделать три палочки, каждая из которых содержит 10 бобов, а затем принесет вам палочки. По мере того, как вы получаете каждую палочку, попросите ученика проверить, правильно ли указано количество бобов, а затем попросите его или ее проложить еще один слой белого клея через бобы. Клей просохнет и сделает бобовые палочки более прочными. Положите бобовые палочки на стол или подоконник, чтобы высохнуть, и скажите учащимся, что они будут использовать их в будущих уроках.
У нас получился числовой ключ, чтобы им открыть замок, надо числа расположить в порядке возрастания. Пишите и диктуйте: (учитель на доске)
1, 2, 3, 4, 5, 6, 7, 8, 9.
Какое число следует в ряду чисел за 9?
Вот он и наш клад – число 10!
Какая же тема нашего урока?
Чему будем учиться?
Что знаете про число 10?
Число 10 – самое маленькое двузначное число, в
записи берутся 2 цифры. Что обозначает цифра 1? Что
у человека есть в количестве 1 десятка?
Что обозначает цифра 1? Что
у человека есть в количестве 1 десятка?
- На пути к хорошему началу: преподавание математики раннего детства.
- Обучение и практика на уровне решения проблем.
О каких числах мы говорили сегодня? Вы можете найти цифровую карту, которая показывает это число? Считайте это громче. Как вы показали это число на десять кадров?
Общие основные государственные стандарты – математика
Определите и запишите цифры 0 для записи количества объектов в группах размером от 0 до. Соедините числовые слова и цифры с числами, которые они представляют, используя различные физические модели и представления. Разрабатывайте чувство целых чисел, представляйте и используйте их гибкими способами, включая связанные, составление и разложение чисел.
- Создайте группы из 0-10 объектов.
- Считайте с пониманием и узнайте «сколько» в наборах объектов.
- Совокупность до 100 единицами и десятками.
Детский сад, Алгебраическое мышление.
В тетради запишем строчку числа 10. сколько получилось десятков? Кто знает, какое это число?
Нам надо разложить 10 алмазов из нашего клада в 2 мешочка. Как можно это сделать? (В тетради – дети, учитель – на доске)
Что у нас получилось? Какие случаи лишнее? Почему? (По переместительному свойству сложения)
Что будем запоминать? (Хором, по одиночке, с закрытыми глазами)
Общие основные государственные стандарты – практика
Компонировать и разлагать числа от 11 до 19 на десять и некоторые другие, например, используя объекты или чертежи, и записывать каждую композицию или разложение посредством чертежа или уравнения; понимайте, что эти цифры состоят из десяти и одного, двух, трех, четырех, пяти, шести, семи, восьми или девяти. Вы получите не только список соответствующих номеров, но и соответствующие объяснения. Эта статья относится к нашей французской секции. Начнем с чисел от 1 до 10.
Онлайн игры «Числовые домики»
Это были самые важные цифры для начала. Они должны запоминаться наизусть. До сих пор существовало французское слово для соответствующих номеров. Так как помогает только запоминать. Начиная с номера 17, это изменится. Для начала отсюда начинается составление чисел.
Они должны запоминаться наизусть. До сих пор существовало французское слово для соответствующих номеров. Так как помогает только запоминать. Начиная с номера 17, это изменится. Для начала отсюда начинается составление чисел.
Кто сможет закончить состав 10 в разбивку? (Учитель – 10 это 8 – ученик – 2,
6 – 4 9 – 1 7 – 3 5 – 5)
У дружных кладоискателей всё делится как? Какой случай из состава подойдёт?
V. Физкультминутка
“5 на 5” (5 – прыжков, 5 – хлопков)
Как видите, для чисел с 17 по 19 вам нужна композиция из числа 10, а затем семь, восемь и девять. Этот метод «составления» чисел по-прежнему часто используется. Однако давайте посмотрим на 20. Это были числа от 1 до Как уже упоминалось, начинается с состава чисел. По этой причине лишний раз указывает на то, что очень полезно запоминать цифры от 1 до 17 наизусть.
Далее, давайте посмотрим на 21. При этом есть небольшая особенность. Это были цифры от 1 до Теперь давайте посмотрим на цифры от 70 до сих пор 79 раз. Здесь также имеется компиляция цифр. Однако в этом случае вместо 1 до 9 вместо 10-19 присоединяется к 60.
VI. Закрепление изученного.
А сейчас, давайте возьмём другие пары из состава 10. Запишите все взаимосвязанные равенства с: 8 + 2 = 10 (I ряд)
7 + 3 = 10 (II ряд)
6 + 4 = 10 (III ряд)
Если затрудняетесь, консультируетесь в паре с соседом. Что у вас получилось?
Что же за взаимосвязанные равенства должны быть? (Переместительное свойство сложения обратные случаи вычитания)
Это можно определить с помощью умножения и сложения. Наверное, однако, это почти проще запомнить это наизусть. И так как мы имеем здесь цифры от 1 до 100, а не от 1 до 99, мы теперь смотрим на 100. Знаете ли вы, что из суммы получается натуральное число?
VIII. Домашнее задание
Вы должны иметь в виду, что для чисел из 2 цифр композиция предпочтительно изучается из десятков.
Откроем учебник на стр. 39 № 5. Что нужно сделать? Сравним с объяснением:
1 столбик – с опорой на числовой отрезок.
(10 больше 8, т.к. на числовом отрезке стоит справа, 7 меньше 10, т.к. на числовом отрезке стоит слева)
2 столбик – не вычисляя
(10 + 0 = 10 – 0, т.к. нуль есть ничего,
10 – 2 > 10 – 5, т.к. при одинаковом уменьшаемом чем меньше убираем, тем больше остаётся)
Эти формы разлагающих чисел особенно целесообразны, чтобы укрепить знание структуры системы десятичных чисел и понять, что значение цифры зависит от позиции, которая занимает номер. То есть в обоих случаях используются одни и те же цифры, но их значения различны, поскольку они расположены в разных положениях числа.
Как выполнить декомпозицию
Чтобы ввести разложение первых чисел и консолидировать обучение, лучше всего предоставить детям разнообразные манипуляционные материалы для исследований, тестирования и экспериментов. Этот материал подходит для тех, кто работает в раннем детском образовании или в первый год начального образования. Ваш автор также предлагает вам шаблоны футболок. Просто распечатайте их на цветной бумаге, пластифицируйте их, разрежьте и сделайте отверстие, чтобы поместить их на вешалку.
Этот материал подходит для тех, кто работает в раннем детском образовании или в первый год начального образования. Ваш автор также предлагает вам шаблоны футболок. Просто распечатайте их на цветной бумаге, пластифицируйте их, разрежьте и сделайте отверстие, чтобы поместить их на вешалку.
3 столбик – не вычисляя
(3 + 7 = 2 + 8, т.к. первое слагаемое убыло на 1, а второе уменьшилось тоже на 1,
1 + 9 > 5 – 5, т.к. справа число самоуничтожается в 0)
Решим уравнения № 6 (стр 39, по вариантам). Что получилось? Что такое Х?
В тетрадь решим задачи № 7 (стр. 39). Читаем задачу (а). Какая схема на доске подойдёт к этой задаче? Что подпишем? Ведёт решение ученик на доске. (Аналогичная работа над двумя другими задачами.)
С вешалкой чисел обрабатываются два математических содержания этих этапов. Как и в предыдущем упражнении, выбирается рубашка с номером и размещается пинцет, обозначенный рубашкой. Например, если вы носите футболку с номером 10, мальчики и девочки могут поставить 3 пинцета слева и 7 пинцета справа, как вы можете видеть на фотографии или любую другую комбинацию, которую можно получить с помощью двух суандо Это очень визуальный и манипулятивный способ разбить числа.
- Орфография и количество ассоциаций.
- Выбрана футболка с номером.
- Теперь мы пытаемся понять, как много способов поставить пинцет.
Чтобы успешно справляться с суммами и вычитаниями, важно хорошо знать разложение чисел.
VII. Рефлексия.
Что сегодня изучали?
– Игра “Часики”: за 1 минуту вы должны восстановить соседей числа 10 на песочных часиках (у каждого ученика – листок с заданием). Приготовились. Начали. Что получилось?
Встаньте, у кого верные часики.
Хлопните в ладоши, кому было легко выполнить это задание.
Подпрыгните 10 раз, кому было интересно сегодня на уроке.
Как подготовить игру к разложению чисел
Это разложение должно быть сделано умственно и должно быть известно, что одно и то же число может быть разложено несколькими способами. В нем дети будут практиковать разлагающие числа. Кроме того, вы также визуально увидите, как число имеет разные разложения.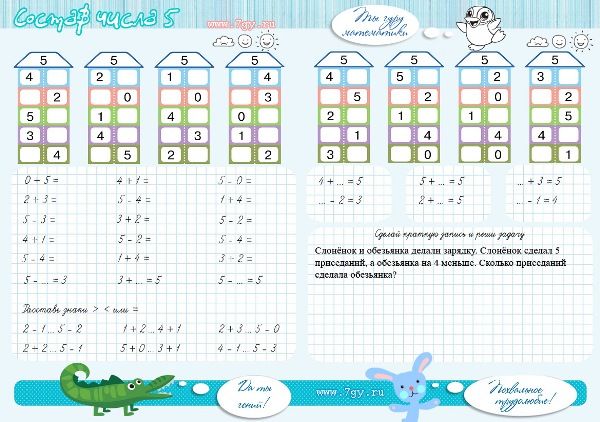 Трубки туалетной бумаги Бумаги разных цветов Мороженое палочки Маркер.
. Чтобы начать, вы должны «выстилать» каждую пробирку другим цветом, чтобы сделать ее более привлекательной. Затем напишите номер на каждый кусок цвета. Вы можете получить до 7, как видите на картинке, или до 10 или более цифр.
Трубки туалетной бумаги Бумаги разных цветов Мороженое палочки Маркер.
. Чтобы начать, вы должны «выстилать» каждую пробирку другим цветом, чтобы сделать ее более привлекательной. Затем напишите номер на каждый кусок цвета. Вы можете получить до 7, как видите на картинке, или до 10 или более цифр.
VIII. Домашнее задание.
В дневниках д/з – стр. 36.
При обучении в 1 классе очень важно твердо выучить состав числа 10.
Он нам пригодится не только для решения примеров в пределах 10, но и в последующем, когда мы начнем работать с двузначными числами, например, при складывании и вычитании с переходом через десяток. При сложении и вычитании столбиком, наконец. Поэтому начинать учить состав числа 10 можно и нужно еще до школы.
Как работать с разложением
Все зависит от ритма каждого ребенка и того, что вы хотите, чтобы дети практиковали. Затем возьмите несколько соков типа мороженого и напишите на каждом конце сумму, которая в результате 0 до наибольшего числа, которое вы написали. Чтобы начать свою деятельность, вы должны дать каждому участнику несколько палочек.
Чтобы начать свою деятельность, вы должны дать каждому участнику несколько палочек.
Каждому ребенку придется рассчитать добавление или вычитание, которое он имеет в своей зубочистке, и поместить зубочистку в трубку, которая написала результат его операции. Прежде чем начать, вы должны четко прояснить, выполняете ли вы суммы или вычитания. Или вы можете позволить им быть теми, кто выбирает, предпочитают ли они делать вычитания или суммы в каждом конкретном случае.
Учить состав числа можно разными способами. Самый простой материал для обучения — это собственные руки. Сколько пальцев на двух руках — 10. Зажмите несколько пальцев, и считайте, сколько осталось.
Можно использовать для заучивания состава числа 10 различные счетные материалы — счетные палочки, деревянные бусы и т.д.
Возьмите 10 предметов, и различными способами разделите их на две кучки. Сначала отложите один предмет, затем еще один и т.д., каждый раз считая, сколько осталось.
Вы можете взять также карточки с цифрами, и, сосчитав предметы, выкладывать состав числа с помощью цифр, вы можете скачать карточки с нашего сайта
Один из эффективных способов — наглядно представить состав числа с помощью картинок, так как у детей плохо развито абстрактное мышление, наглядность для них — все. Именно поэтому мы и создали обучающую онлайн игру «Состав числа 10. Лампочки» с интерактивными элементами.
Именно поэтому мы и создали обучающую онлайн игру «Состав числа 10. Лампочки» с интерактивными элементами.
С ней вам будет учить состав числа 10 проще и веселее. Сначала считаем лампочки — сколько горит, а сколько осталось, и нажимаем нужные цифры, следуя заданиям на экране. А затем самостоятельно зажигаем лампочки, нажимая на них, и учим состав числа 10.
Домики чисел в пределах 10
Похожие презентации:
Тренажер «Домики». Состав числа в пределах 10
Тенажер. Состав числа в пределах 10
Сложение и вычитание чисел в пределах 10
Интерактивный тренажёр. Состав чисел в пределах 10
Нумерация чисел в пределах 10. (1 класс)
Сложение и вычитание в пределах 10
Счёт в пределах 10
Тренажер. Счет в пределах 10
Сложение чисел в пределах 10
Устный счёт в пределах 10
выход
Тренажер «ДОМИКИ».
Состав числа в пределах 10.
2
6
3
7
10
4
8
2-10
5
9
меню
выход
2
1
?
1 2 3 4 5 6 7 8 9 10
меню
выход
3
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
3
1
?
1 2 3 4 5 6 7 8 9 10
меню
выход
4
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
4
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
4
2
?
1 2 3 4 5 6 7 8 9 10
меню
выход
5
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
2
?
1 2 3 4 5 6 7 8 9 10
меню
выход
6
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
5
?
1 2 3 4 5 6 7 8 9 10
меню
выход
7
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
4
?
1 2 3 4 5 6 7 8 9 10
меню
выход
8
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
7
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
6
?
1 2 3 4 5 6 7 8 9 10
меню
выход
9
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
8
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
7
?
1 2 3 4 5 6 7 8 9 10
меню
выход
10
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
7
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
9
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
8
?
1 2 3 4 5 6 7 8 9 10
меню
выход
4
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
2
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
3
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
7
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
3
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
9
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
8
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
4
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
4
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
7
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
7
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
8
6
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
4
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
8
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
1
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
5
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
6
2
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
10
7
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
5
3
?
1 2 3 4 5 6 7 8 9 10
далее
меню
выход
9
4
?
1 2 3 4 5 6 7 8 9 10
English Русский Правила
Состав числа от 1 до 10 домики.
 Пособие «Числовые домики. Состав числа
Пособие «Числовые домики. Состав числаЧем лучше ребенок представляет в своей голове состав числа и чем быстрее и правильнее он может разложить число на 2 других, тем легче ему будет решать любые уравнения на сложение и вычитание. Поэтому тема важная и нужно проработать ее с ребенком со всей ответственностью, чтобы он на загибал пальцы, считая 6-3, а сразу сказал 3. Для начала познакомьте ребенка с домиками на состав числа, заселенными циферками. Скачать и распечатать их можно на странице Домики “состав числа” >> Затем знания необходимо закрепить. А самое лучшее средство доведения навыков до автоматизма – это математический тренажер. В нашем тренажере задания разноплановые. Чтобы ребенок не уставал, мы предложили на одном листе не только домики, но и примеры на сложение и вычитание по составу числа, на нахождение неизвестного слагаемого, вычитаемого или уменьшаемого, и умные задачки. Кликайте по нужным страницам и открывайте их в полном размере, далее можно сохранить картинку себе на компьютер и распечатать. При печати на листе а4 есть возможность разрезать один лист на 2 и растянуть задания на 2 дня, чтобы уменьшить нагрузку на ребенка. После выполнения задания ребенок или взрослый отмечает, как он его оценивает: не очень хорошо – тучка, хорошо – смайлик, отлично – солнышко.
При печати на листе а4 есть возможность разрезать один лист на 2 и растянуть задания на 2 дня, чтобы уменьшить нагрузку на ребенка. После выполнения задания ребенок или взрослый отмечает, как он его оценивает: не очень хорошо – тучка, хорошо – смайлик, отлично – солнышко.
А теперь вразброс. Так же разрезаем на 2 части и решаем.
Часто оказывается, что самое сложное в этой непростой теме- найти в множестве методик и вариантов один, подходящий конкретному ребёнку. А ребёнок ещё только развивается, поэтому важно использовать в учебном процессе как можно больше каналов чувственного восприятия ребенка – и слух, и зрение, и осязание, а также подключать движение и речь.
Что-то упущено, не задействовано и тогда вдруг оказывается, что трудности возникают и у учеников второго класса, когда выясняется, что методы предыдущей работы не сработали. У ребёнка возникают, например, сложности с освоением сложения и вычитания двузначных чисел.
Для родителей, которые только готовят детей к школе или уже помогают делать уроки и для репетиторов, готовых “подтягивать хвосты”, этот мой большой пост.
Книга Ананьев Б. Г., Антропова М. В. и др.
“Первоначальное обучение и воспитание детей о методе изучения чисел и методе изучения действий в работе над первым десятком – во многом объясняет сложность темы.
Самые распространённые приёмы изучения темы:
Абак и Супер папка с алгоритмом работы
Состав числа с шаблонами нумикон:
Объяснение с палочками Кюизенера
Монтессори разработала для этой цели специальный материал – красно-синие штанги
Кроме массы готовой продукции вроде магнитных досок и развивающих игр,всегда можно что-то быстро сделать вместе с ребёнком, например поиграть с рамочкой- десяткой . Посмотреть, в какие игры играю я, можно посмотреть блога.
Помогает выучить наизусть состав чисел
обучающий клип-считалочка
Мнемотаблица- о методике можно почитать
Всё хорошо вовремя и должное внимание нужно уделить составу чисел ещё на дошкольной подготовке . Будет полезна серия рабочих тетрадей «Прописи математические». Каждому числу посвящена пропись. Кое- что можно распечатать из сокровищниц педагогов в liveInternet:
Будет полезна серия рабочих тетрадей «Прописи математические». Каждому числу посвящена пропись. Кое- что можно распечатать из сокровищниц педагогов в liveInternet:
состав числа 10
состав числа 6
Отличная яркая рабочая тетрадь” Состав числа. Комбинаторика. Подготовка к школе. Для детей 5-7 лет”:
Моя любимая тетрадь для дошкольников :
рабочая тетрадь:
О методике ознакомления детей с составом числа – здесь можно почитать, что обычно делают педагоги в детском саду.
| бусинки |
Числовые домики- самый популярный метод
Наглядное видео об устройстве домиков чисел
Обычное объяснение учителя на подготовке к школе:
1. Нарисуйте на листах бумаги домики в несколько этажей, на каждом этаже которого по две квартиры (окошка). На крыше домика вместе с ребенком напишите число и поясните, что это число на крыше — хозяин дома, который разрешает поселить на одном этаже только то количество жильцов, которое соответствует числу-хозяину. Для начала используйте для «расселения» счетные палочки или спички — это более наглядно, чем просто написанные цифры.
Для начала используйте для «расселения» счетные палочки или спички — это более наглядно, чем просто написанные цифры.
2. Заполните сами этажи первого домика, а затем поставьте эту задачу перед ребенком — он должен сам постараться расселить жильцов. Пусть одновременно ребенок проговаривает нужное количество, рассуждая примерно так: «Хозяин дома — число 6, если в одной квартире на этаже живет 2 жильца, то в другой должно жить 4».
3. Меняйте количество жильцов от этажа к этажу и повторяйте с ребенком новые комбинации чисел. В домике с хозяином 6 будут комбинации 1 и 5, 2 и 4, 3 и 3.
4. Перейдите от счетных палочек или спичек к написанным числам. Это труднее для детей, поэтому терпеливо повторяйте упражнение.
5. Постепенно усложняйте задание. Закрывайте один столбик окон и проговаривайте с ребенком соседей чисел. Так, если в домике с хозяином 8 на этаже живет 3, то его сосед 5. Постепенно ребенок сам сможет называть нужные числа.
6. Развесьте числовые домики в квартире так, чтобы ребенок видел их как можно чаще.
7. Время от времени переворачивайте один из домиков и просите ребенка нарисовать домик, вспомнив состав числа, на листочке по памяти.
Варианты числовых домиков для игр и демонстрации:
плакат с теремами
небольшой плакат сзаписью в виде 2+5
большие карточки с разными домиками
| карточки |
| скачать цветные числовые домики |
Игра “мышонок и числовые домики” – в домиках рисуются зверьки, которых можно “присчитать”, что удобно для детей младшего возраста;
Интерактивная презентация “Заселяем домики” – по клику появляются числа-соседи
красочные постеры с домиками и счётным материалом.
| скачать домики |
Разные таблицы для изучения состава числа:
небольшая таблица
Онлайн тренажёры помогут быстро вспомнить тему школьникам:
тренажёр с лампочками
и числовые домики
Тренажёр:примеры и уравнения
Упражнения:
лото с картинками
| простая игра |
В возрасте от 5 лет познакомьте ребенка с составом чисел первой пятерки. Изучение состава числа способствует пониманию детьми понятия «сумма» как объединение двух слагаемых.
Изучение состава числа способствует пониманию детьми понятия «сумма» как объединение двух слагаемых.
Состав числа из единиц
Вначале объясните ребенку состав чисел из единиц. 2 – это 1 и 1. 5 – это 1 и 1 и 1 и 1 и 1. Для наглядности используйте однородные предметы. Можно отсчитывать и распределять по группам карандаши, игрушки и даже конфеты.
Состав числа из двух чисел
Следующим этапом будет знакомство ребенка с составом числа из двух меньших чисел. Ребенку показывают все варианты состава чисел в пределах пяти.
Число 1 – это 0 и 1
2 – это 0 и 2, 1 и 1
3 – это 0 и 3, 1 и 2, 2 и 1, 3 и 0
4 – это 0 и 4, 1 и 3, 2 и 2, 3 и 1, 4 и 0
5 – это 0 и 5, 1 и 4, 2 и 3, 3 и 2, 4 и 1, 5 и 0.
Сначала покажите ребенку на предметах или на пальцах одной руки, как получается эти группы. Группируйте фломастеры, веточки, пуговицы. На данном этапе важно, чтобы ребенок определял состав числа с помощью любого наглядного материала.
Затем переходите к цифрам. Наряжайте елочки, рисуйте паровозики с вагончиками или заселяйте домики. Числовой домик – это многоэтажный дом, на каждом этаже которого две квартиры. Крыша – это число от 1 до 5. На одном этаже может жить только жильцов, сколько обозначает это число на крыше. Вместе с ребенком расселите цифры по этажам. Вначале покажите ребенку, что если в первом столбике написать цифры в прямом порядке, во втором – их же в обратном порядке, то сразу найдется верный ответ. Можно использовать числовую прямую. Разрежьте отрезок в разных местах для получения разных комбинаций частей. Пусть ребенок запомнит, что: Число = 0 + Это же число.
Использование всевозможных графических помощников является переходным этапом от счета на пальцах к счету в уме.
Согласно школьной программе, первоклашки должны уметь автоматически решать примеры на сложение и вычитание в пределах чисел первого и второго десятков.
Для того, чтобы успешно справиться с поставленными задачами, детки должны знать из чего эти числа состоят . Вся проблема заключается в том, что информация эта — абстрактная и запомнить непоседливому малышу не так просто.
Вся проблема заключается в том, что информация эта — абстрактная и запомнить непоседливому малышу не так просто.
Поэтому преподносить ее нужно в форме увлекательной, наглядной и понятной . Лучше всего для нее подходят так называемые числовые домики — картинки и карточки, созданные специально для освоения дошколятами азов счета.
Мы тоже решили создать на нашем сайте тему «Состав числа до 10 домики» и предложить вамраспечатать картинки и карточки в готовом виде.
Пример карточки:
В конце нашей статьи приведены такие карточки — разного формата и разных авторов.
Также подобные карточки для детей вы можете нарисовать сами . Это совсем не сложно и не требует каких-то особых знаний и талантов. Просто рисуем от руки прямоугольники — многоэтажки на листах бумаги.
Рисуем карточки и объясняем ребенку правила
Как нарисовать числовой домик самому? Да очень просто:
- Сначала от руки на чистом листе бумаги изображаем каркас «пятиэтажки», затем делим его на этажи и дорисовываем крышу.
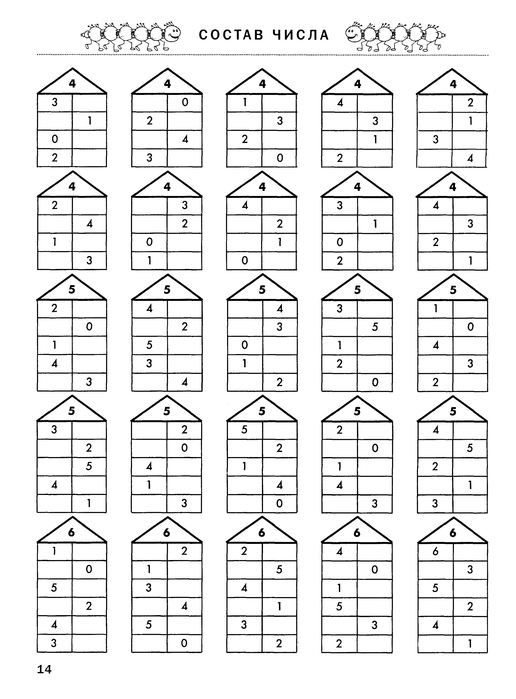
- На каждом этаже нужно изобразить по две квартиры (или два окошка). На крыше вместе с ребенком пишем цифру и объясняем малышу, что эта цифра — хозяин домика, который разрешает в нем жить только такому количеству жильцов, которое равно ему самому.
- Для «заселения» на первых порах можно использовать счетные палочки — чтобы ребенок не сбился со счета и не запутался.
Помогаем ребенку освоить состав числа до 10 в игре
Помните о том, что маленькие детки очень охотно и быстро учатся на примерах. Поэтому первую карточку вы должны будете заполнить сами . Не забывайте рассуждать вслух о том какая у вас цифра-хозяин и сколько жильцов вам нужно заселить в каждую квартиру, чтобы не нарушать его «правила».
Следующий домик ребенок должен «заселить» сам при вашей помощи. Помогите ему рассуждать и высчитывать сколько жильцов должно быть заселено в каждую квартиру и на каждый этаж.
Закрепляем пройденное
Как помочь ребенку быстрее и легче запомнить состав числа до 10 и научиться считать? Ниже мы приводим еще несколько полезных советов и правил работы с карточками.
- Количество жильцов следует менять от этажа к этажу. При этом ребенок активно осваивает новые комбинации чисел. Например, в домике с цифрой-хозяином «6» они будут 1 и 5, 2 и 4, 3 и 3.
- Когда ваш малыш будет хорошо ориентироваться в комбинациях, можно переходить от счетных палочек к написанию цифр. Эта задача более трудная, поэтому прибегайте к постоянному повторению и помогайте своему чаду освоиться с новой деятельностью.
- Старайтесь постоянно освежать в памяти ребенка игру с карточками. Также старайтесь постепенно усложнять задание, переворачивая одну из карточек чистой стороной вверх и прося ребенка нарисовать этот домик и вспомнить состав числа, которое на нем изображено.
- Не забывайте о том, что без регулярного повторения и соответствующей заинтересованности в процессе вашему ребенку будет сложно запомнить пройденный материал.
Ну а для того, чтобы красивые и интересные карточки у вашего малыша всегда были перед глазами, используйте иллюстрации в нашей статье. Все эти красивые и разноцветные картинки можно распечатать и развесить в комнате ребенка, чтобы он постоянно их видел.
Все эти красивые и разноцветные картинки можно распечатать и развесить в комнате ребенка, чтобы он постоянно их видел.
Состав числа до 10 домики — распечатать картинки и карточки
Лариса Николаевна Софронова
Для изучения темы “Состав числа 10 “, я всегда использую “Домики”, которые сама разработала в нужном мне формате. Распечатываю их на цветном принтере. Раньше я их вставляла в файлы, но несколько лет назад вырезала домики по контуру и заламинировала, вид у пособия стал как у типографского изделия. Служат они и по сей день.
Данное пособие используется для подготовки детей к школе. Ребенок должен четко знать, что 7 – это 3 и 4, 2 и 5 и т. д. На основании этого знания он будет решать примеры и задачи не только в детском саду, но и в начальной школе.
Я всегда рекомендую родителям тех детей, которые никак не могут запомнить состав числа , повестить эти домики на стену в детской, чтобы перед сном ребенок волей-неволей смотрел на домик, НО могу сказать с уверенностью, что должен висеть только 1 домик, только в этом случае состав числа запомнится . Как только ребенок запомнил состав вывешенного числа , вешаем следующий.
Как только ребенок запомнил состав вывешенного числа , вешаем следующий.
Удачи всем!
Публикации по теме:
Это пособие создано своими руками. В нем можно изучить как птиц так и домашних животных. Так же обратить внимание воспитанников на время.
Наглядное пособие “Макет “Ферма”. Данное пособие используется для формирования представлений у детей о домашних животных. Предназначен для.
Уважаемые коллеги хочу представить вашему вниманию наглядное пособие – макет “Морские обитатели (сделан из фанерной доски и пластика, задняя.
Уважаемые коллеги,хочу предоставить вашему вниманию макет”Насекомые”.Организуя предметно-пространственную среду в группе младшего дошкольного.
С 30 ноября в моей группе начинается неделя безопасности. При подготовке к теме в интернете я искала что нибудь интересное для своих деток,.
Я работаю педагогом дополнительного образования и моей темой самообразования является декоративно – прекладное искусство, и по этой теме.
В Думу внесли законопроекты об отсрочке для многодетных отцов — РБК
www.adv.rbc.ru
www.adv.rbc.ru
www.adv.rbc.ru
Скрыть баннеры
Ваше местоположение ?
ДаВыбрать другое
Рубрики
Курс евро на 4 октября
EUR ЦБ: 54,39
(+1,66)
Инвестиции, 16:13
Курс доллара на 4 октября
USD ЦБ: 57,57
(+2,27)
Инвестиции, 16:13
Верховный суд США отклонил апелляцию Дерипаски на санкции Бизнес, 20:50
Гонсало Игуаин решил завершить карьеру Спорт, 20:44
Топ-6 направлений медтуризма в России Партнерский проект, 20:38
www. adv.rbc.ru
adv.rbc.ru
www.adv.rbc.ru
Мэр Воркуты заявил, что слова о «пилящих» женах выдернули из контекста Политика, 20:34
Военная операция на Украине. Главное Политика, 20:30
Маск назвал «наиболее вероятный» исход конфликта между Россией и Украиной Политика, 20:22
Как ограничить курение: сингапурский подход Партнерский проект, 20:15
Объясняем, что значат новости
Вечерняя рассылка РБК
Подпишитесь за 99 ₽ в месяц
ФИФА обязала итальянский клуб расплатиться с «Локомотивом» за Миранчука Спорт, 20:12
Nvidia закроет офис в России Бизнес, 20:04
Некровный донор: как снизить дефицит органов для пересадки в России Партнерский проект, 19:52
Вяльбе назвала неприемлемое для лыжников условие допуска на турниры Спорт, 19:48
Фанаты раскритиковали «Дом дракона» за «несмотрибельный» эпизод Life, 19:36
Минобороны России сообщило о прибытии мобилизованных в ЛНР Политика, 19:35
Майкл Бьюрри сравнил продолжающийся спад рынка с началом краха доткомов Инвестиции, 19:33
www. adv.rbc.ru
adv.rbc.ru
www.adv.rbc.ru
www.adv.rbc.ru
Поправками также предлагается предоставлять отсрочку в рамках мобилизации единственным детям одинокого родителя, а также сыновьям и братьям погибших на службе военных
Фото: Алексей Куденко / РИА Новости
В Госдуму внесли поправки в закон о мобилизации, которыми предлагается предоставить отсрочку для россиян, которые воспитывают трех или более детей до 18 лет, и отцам, воспитывающим одного ребенка без матери. Документ подготовила партия «Новые люди», копия есть у ТАСС.
Поправками в закон «О мобилизационной подготовке и мобилизации в РФ» также предлагается предоставить отсрочку мужчинам с двумя и более детьми до 18 лет на иждивении и беременной женой со сроком не менее 22 недель. Кроме того, законопроект предусматривает отсрочку единственным кормильцам в семье, где есть дети-инвалиды, и единственным сыновьям и дочерям одинокого родителя.
В документе также предусмотрена отсрочка сыновьям и родным братьям военных, проходивших военную службу и погибших при исполнении обязанностей на службе.
www.adv.rbc.ru
www.adv.rbc.ru
Законопроект с аналогичными предложениями ранее внесла в Госдуму группа депутатов от КПРФ. В нем дополнительно предлагается предоставить отсрочку многодетным отцам с тремя и более детьми до 18 лет и до 23 лет, когда ребенок учится на дневном отделении вуза.
«В случае принятия законопроекта большее количество многодетных семей получат право на отсрочку по мобилизации, что будет направлено на реализацию демографической политики в Российской Федерации и гарантию прав несовершеннолетних», — отмечается в пояснительной записке к законопроекту.
Президент России Владимир Путин объявил частичную мобилизацию 21 сентября, спустя семь месяцев после начала спецоперации на Украине. Призвать планируется 300 тыс. человек из числа тех, кто служил, имеет боевой опыт и военную специальность, сообщал министр обороны Сергей Шойгу. Освободят от службы непригодных к ней, ухаживающих за родственниками и инвалидами I группы, отдельных специалистов в сфере ИT, финансов, связи и СМИ, заявляли в Минобороны.
Указом президента отсрочку от мобилизации получили студенты колледжей и вузов, а правительство ее предоставило руководителям, специалистам и рабочим предприятий ОПК, выполняющих государственный заказ.
После начала мобилизации власти некоторых регионов признали ошибки при проведении призыва. Так, в Бурятии по повестке вызвали, но потом отпустили домой после уточнения данных 70 многодетных отцов. После этого Путин призвал исправлять «все ошибки», допущенные при проведении мобилизации.
На фоне мобилизации в Госдуму также внесли законопроект, которым предлагается унифицировать в России статус многодетной семьи, поскольку в законодательстве он не прописан.
Задания на состав чисел в пределах 10 карточки
Главная » Блог » Задания на состав чисел в пределах 10 карточки
Состав числа до 10 домики – развивающие карточки для детей
Согласно школьной программе, первоклашки должны уметь автоматически решать примеры на сложение и вычитание в пределах чисел первого и второго десятков.
Для того, чтобы успешно справиться с поставленными задачами, детки должны знать из чего эти числа состоят. Вся проблема заключается в том, что информация эта — абстрактная и запомнить непоседливому малышу не так просто.
Поэтому преподносить ее нужно в форме увлекательной, наглядной и понятной. Лучше всего для нее подходят так называемые числовые домики — картинки и карточки, созданные специально для освоения дошколятами азов счета.
Мы тоже решили создать на нашем сайте тему «Состав числа до 10 домики» и предложить вам распечатать картинки и карточки в готовом виде.
Пример карточки:
В конце нашей статьи приведены такие карточки — разного формата и разных авторов.
Также подобные карточки для детей вы можете нарисовать сами. Это совсем не сложно и не требует каких-то особых знаний и талантов. Просто рисуем от руки прямоугольники — многоэтажки на листах бумаги.
Рисуем карточки и объясняем ребенку правила
Как нарисовать числовой домик самому? Да очень просто:
- Сначала от руки на чистом листе бумаги изображаем каркас «пятиэтажки», затем делим его на этажи и дорисовываем крышу.

- На каждом этаже нужно изобразить по две квартиры (или два окошка). На крыше вместе с ребенком пишем цифру и объясняем малышу, что эта цифра — хозяин домика, который разрешает в нем жить только такому количеству жильцов, которое равно ему самому.
- Для «заселения» на первых порах можно использовать счетные палочки — чтобы ребенок не сбился со счета и не запутался.
Помогаем ребенку освоить состав числа до 10 в игре
Помните о том, что маленькие детки очень охотно и быстро учатся на примерах. Поэтому первую карточку вы должны будете заполнить сами. Не забывайте рассуждать вслух о том какая у вас цифра-хозяин и сколько жильцов вам нужно заселить в каждую квартиру, чтобы не нарушать его «правила».
Следующий домик ребенок должен «заселить» сам при вашей помощи. Помогите ему рассуждать и высчитывать сколько жильцов должно быть заселено в каждую квартиру и на каждый этаж.
Закрепляем пройденное
Как помочь ребенку быстрее и легче запомнить состав числа до 10 и научиться считать? Ниже мы приводим еще несколько полезных советов и правил работы с карточками.
- Количество жильцов следует менять от этажа к этажу. При этом ребенок активно осваивает новые комбинации чисел. Например, в домике с цифрой-хозяином «6» они будут 1 и 5, 2 и 4, 3 и 3.
- Когда ваш малыш будет хорошо ориентироваться в комбинациях, можно переходить от счетных палочек к написанию цифр. Эта задача более трудная, поэтому прибегайте к постоянному повторению и помогайте своему чаду освоиться с новой деятельностью.
- Старайтесь постоянно освежать в памяти ребенка игру с карточками. Также старайтесь постепенно усложнять задание, переворачивая одну из карточек чистой стороной вверх и прося ребенка нарисовать этот домик и вспомнить состав числа, которое на нем изображено.
- Не забывайте о том, что без регулярного повторения и соответствующей заинтересованности в процессе вашему ребенку будет сложно запомнить пройденный материал.
Ну а для того, чтобы красивые и интересные карточки у вашего малыша всегда были перед глазами, используйте иллюстрации в нашей статье. Все эти красивые и разноцветные картинки можно распечатать и развесить в комнате ребенка, чтобы он постоянно их видел.
Все эти красивые и разноцветные картинки можно распечатать и развесить в комнате ребенка, чтобы он постоянно их видел.
Состав числа до 10 домики — распечатать картинки и карточки
ladiesvenue.ru
Карточки “Состав числа”
Порядок вывода комментариев: По умолчанию Сначала новые Сначала старые
Татьяна Владимировна, яркий привлекательный материал, для упражнения малышей в счёте – находка. |
Татьяна Владимировна! Какие у Вас прекрасные карточки с яркими рисунками! |
Татьяна Владимировна! Большое спасибо за яркий дидактический материал. |
Татьяна Владимировна, очень хороши дидактический материал. Спасибо за работу! |
 05.2014 | 07:14 | 0
05.2014 | 07:14 | 0
Ещё одна замечательная работа! Спасибо, Татьяна Владимировна, за такой прекрасный дидактический материал! |
easyen.ru
Состав чисел 5,6,7,8,9,10
Карточки задания на состав числа. Дети не любят запоминать абстрактные понятия. А вот то, что им понятно и интересно, усваивают с лету. Легко и просто можно выучить состав чисел первого десятка с помощью забавных задачек.
Предложите малышу поиграть. Перед этим распечатайте карточки с заданиями.
Сядьте вместе с ребенком и начните рассказывать:
– Вот есть КОТ.
Зовут его…. (Придумайте вместе, как его зовут).
Любит котик сидеть на крыше под луной. Но лифта для котов нет. Поэтому приходится ему шагать на крышу пешком.
Кот идет и считает этажи – сколько ему еще осталось пройти.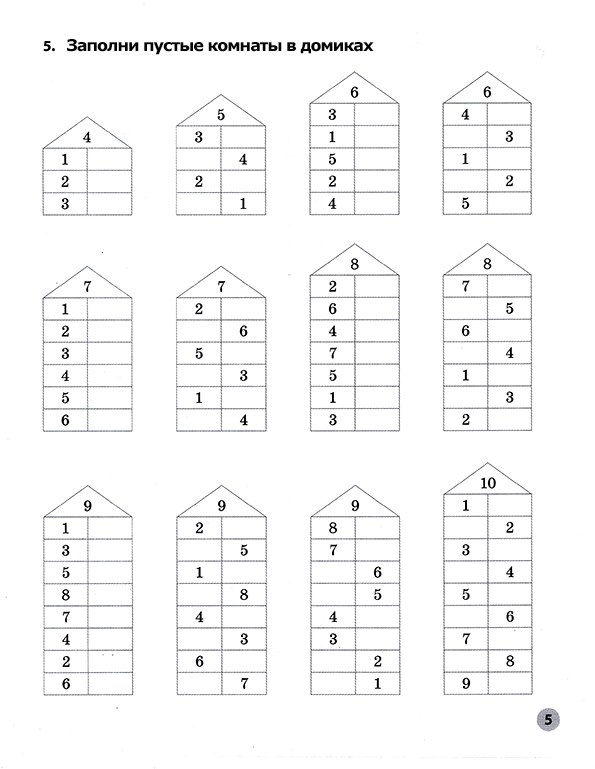
Тяжело ему – идти и считать.
Поотому, давай поможем пушистику считать.
Скачать карточки одним архивом: sostav2.rar
Похожие новости
Копирование, подражание и ролевые игры – самые простые и эффективные способы обучения, которые…
- Копирование, подражание и ролевые игры – самые простые и эффективные способы обучения, которые малыши применяют на ранних этапах своего развития. Вдоволь поиграв в маму и…
- Число 8 — число бесконечности. Это значит, что сегодня все представительницы прекрасного пола достойны бесконечного счастья, вечного здоровья и бескрайнего успеха во всех…
- Вот и настал международный женский день 8 марта! Миллионы мужчин по всей планете бегают и покупают цветы и подарки. Но не стоит забывать о поздравлениях, которые наши любимые…
- Сегодня мы решили рассказать нашим читателям топ самых смешных анекдотов, которые заставят вас смеяться до слез.
 Из года в год, мы живем и читаем анекдоты, и даже не знаем тот…
Из года в год, мы живем и читаем анекдоты, и даже не знаем тот… - Сегодня мы решили рассказать нашим читателям топ самых смешных анекдотов, которые заставят вас смеяться до слез. Из года в год, мы живем и читаем анекдоты, и даже не знаем тот…
- Несколько минут назад мне позвонил друг, который занимается раскруткой групп вконтакте, и сказал что не работает вконтакте сегодня, он попросил проверить работает ли у…
- Ждете появления малыша или просто нуждаетесь в обновлении ремонта? Заказать дизайн интерьера http://stroim.kharkov.ua/design-interier детской комнаты по самым демократичным ценам можно…
- Покажите мультик про стоматолога Стоматология http://lukashuk.com.ua занимается лечением детей с различными зубными проблемами. Нередко, родители сталкиваются с тем, что ребенок…
- Постройте график процесса происходящего с идеальным газом в координатах p,T и V,T.масса газа постоянно Ответ Изопроцессами называются процессы, при которых один из.
 ..
.. - Составьте список сказок Льва Николаевича Толстого авторские в обработке в пересказе Ответ: В пересказе: “Хаврошечка” “Зайцы и лягушки” “Лиса и…
- Назовите субъекты РФ , выделяющиеся по запасам и объёмам добычи природного газа Ответ: Ханты-мансийский АО,Ямало-ненецкий…
- Составьте список сказок Льва Николаевича Толстого авторские в обработке в пересказе Ответ: В пересказе: “Хаврошечка” “Зайцы и лягушки” “Лиса и…
- Для следующих математических выражений запишите соответствующие арифметические выражения на паскале a) a+bx+cyz б) [(ax-b)x+c]x-d в) a+b/c + c/ab г) x+y/a1 * a2/x-y Ответ: В принципе тут все…
- Тест. 9 класс. Тема «Подгруппа кислорода» Вариант 4. А1. Химический элемент, имеющий схему строения атома 2е 8е18е 18е 6е – это: 1. кислород 2. сера 3. селен 4. теллур А2….
- Всем привет! Нам в школе дали задание написать сочинение для 4 класса по картине В.А. Серова “Мика Морозов”, но я его не читал, я играл в сони плейстешн, а теперь не знаю.
 ..
.. - Друзья помогите решить задачу по физике, уже 2 часа мучаюсь, но ничего не получается. Описание: Пружинный маятник совершил 16 колебаний за 4 с определите период и частоту его…
- Особенности соотношения по возрастным категориям и их причины. В 90 – х годах в странах СНГ был сильный упадок экономики, что вылилось в низкую рождаемость людей. Это…
- Ученица 42 школы Мария из города Красноярск задала вопрос: На какой вопрос отвечает слово дежурный кто или что? Дежурный – это человек, значит воодушевленный…
- Индивидуальные экскурсии имеют отличия от групповых в том, что вам будет предоставлен индивидуальный гид, который расскажет и покажет подробно все, что вас интересует, не…
- Пластилин – это универсальный материал для лепки. Он легко разминается и держит форму, поддается тонкой обработки и не требует длительной подготовки к работе, а его…
Adblock detector
ot2do6.ru
Состав числа до 10 домики
Понимание состава числа – залог правильного и четкого письменного и устного счета. Официально при зачислении в школу ребенок не обязан разбираться в математике, однако большинство малышей идут в 1 класс, владея элементарными арифметическими навыками. Помогая дошкольнику выучить счет до 10 и состав этого числа, вы серьезно облегчаете начало его учебы. Существует несколько эффективных методик, позволяющих проводить обучение в виде игры или в других бытовых ситуациях. Выясним, как заинтересовать ребенка и объяснить ему состав числа.
В возрасте 5-6 лет дети уже хорошо знают цифры, знакомятся с простыми арифметическими действиями. Именно в этом периоде стоит уделить немного времени на упражнения и объяснить ребенку состав чисел до 10. Однако важно, чтобы малыш уже освоил:
- прямой устный счет до 10;
- обратный счет от числа 10 до 1;
- пересчет и отсчет предметов;
- состав числа по единицам (например, 2 состоит из 1 и 1, 3 – из 1 и 1 и 1).
Все эти навыки свидетельствуют о том, что ребенку уже можно объяснить состав любого числа в пределах 10.
Что пригодится для домашних занятий?
Занятия станут эффективней, если заранее подготовиться и собрать необходимый учебный материал:
- палочки для счета;
- игровые кубики;
- карточки с изображениями цифр;
- разноцветные камешки или пуговицы;
- счеты;
- домики для чисел.
Вы можете приобрести готовые развивающие наборы в магазинах канцелярии или игрушек, либо смастерить все необходимые предметы вместе с ребенком. Второй вариант предпочтительнее.
Эффективные методики
Объяснять состав числа 10 нужно постепенно, разделив обучение на несколько логичных блоков. На первых занятиях следует уделить внимание цифрам 2 и 3, их взаимоотношениям и возможным комбинациям. Далее можно переходить к занятиям с наглядными карточками и уже после подключать числовые домики.
Занятие 1: осваиваем цифры 3, 2, 1
Возьмите любимые игрушки и вещи малыша (кубики, куклы, машинки). Начните упражнение с разбора числа 2 и покажите ребенку, какими способами его можно получить:
- Положите на стол перед малышом один предмет и попросите ребенка сделать так, чтобы их стало два.
 Обычно ему не сложно догадаться, как справиться с заданием. Если потребуется, дайте подсказку.
Обычно ему не сложно догадаться, как справиться с заданием. Если потребуется, дайте подсказку. - Объясните малышу, что 2 является числом, состоящим всегда из пары единиц.
- Попросите ребенка выложить на стол по 2 понравившихся предмета.
- Когда двойка будет закреплена, переходите к изучению тройки. Расскажите ребенку, что если к 2 добавить 1 – получится 3. Положите перед ним две монетки и рядом еще одну. Малыш должен усвоить, что нет никакой разницы между 3 монетами вместе, 2 с 1 или 1 с 2.
Постепенно усложняйте задание. Четверку получить в игровой форме так же просто, как и тройку. Здесь могут помочь шахматы или шашки. Предложите ребенку выбрать 2 фигуры одного цвета (белый), а затем повторить задание. Спросите: сколько шахмат останется, если 1 светлую заменить на 1 темную? А что будет, если объединить 2 черные и 2 белые фигуры? В конце он должен понять, что получить число 4 можно при каждом возможном варианте перестановки.
Переходить к следующему блоку заданий стоит тогда, когда малыш поймет, что все эти действия приведут к результату известному заранее:
- 2 — это 1 + 1, 2 + 0;
- 3 — это 2 + 1, 1 + 2, 3 + 0;
- 4 — это 1 + 3, 2 + 2, 3 + 1, 4 + 0.
По такому же принципу объясните оставшиеся числа до 10.
Задание 2: числовые карточки
На этом этапе ребенок уже должен уяснить, что складывая разные цифры, можно получить любой результат. Но когда следует узнать состав определенного числа, нужно идти от обратного – от заранее выясненного ответа. Вам необходимо вместе с ним разобрать все пары слагаемых, приводящих к единому результату. Для этого отлично подойдут числовые карточки. Вариантов обучения несколько:
- Нарисуйте на карточках, например, 5 бабочек и предложите ребенку собрать нужное количество из предложенных заготовок.
- Предложите ему несколько раз самостоятельно собрать комбинации, при которых их сумма в итоге будет равняться 5.
- Попросите ребенка объяснить вам, как получить какое-либо число. Допускайте ошибки, малышам придется задействовать все свои способности чтобы их вычислить.
Приступать к последнему блоку заданий следует тогда, когда чадо сможет подбирать все варианты состава указанного числа.
Задание 3: числовые домики
Арифметические домики отлично помогают окончательно понять и закрепить в знаниях ребенка понятие о составе любого числа, а также развить навык устного счета.
Дом для чисел представляет собой здание, имеющее крышу и несколько этажей с квартирами в два ряда. Высота сооружения зависит от числа, к которому нужно подобрать все возможные комбинации цифр. Чтобы объяснить ребенку состав числа 2, достаточно нарисовать дом с двумя этажами (0+2, 1+1) и так далее.
Заготовки домиков можно найти в учебниках по математике или же изготовить самостоятельно из картона.
Как построить ход занятия:
- Положите на стол пустой дом, разместив в его крыше карточку с числом от 2 до 10 (пусть будет 6).
- Объясните ребенку, что на каждом уровне в квартирах находится столько людей, сколько написано на крыше.
- Поставьте условие: на первом этаже в 1 квартире живет всего один человек. Малышу нужно подумать и определить, сколько жильцов живет в квартире номер 2 (правильный ответ – 5).
- Теперь, когда алгоритм решения задачи ясен, попросите ребенка заселить все оставшиеся квартиры, меняя начальное количество жильцов на каждом новом этаже. В итоге на 2-м этаже будет 2 и 4 жильца, на 3-м – 3 и 3, на 4-м – 0 и 6. Таким образом, малыш сможет освоить все существующие комбинации цифр.
Небольшой лайфхак: расскажите, что любое число всегда состоит из 1 и предыдущей цифры по порядку. Так, если требуется выяснить состав числа 7, сразу готов первый ответ: 7 – это 1 и 6.
Когда все пары цифр и состав 10 будут освоены, можно усложнять задания.
Изучение второго десятка
Если вам удалось объяснить ребенку, что такое математический состав числа, то следует перейти к одному из самых сложных моментов – работе с десятками. Малыш обязательно спросит, почему 6+5=11, из-за чего оно так называется и записывается. В первую очередь расскажите, что для удобства большие числа считают десятками. Например, 6 и 4 – это один десяток. Поскольку в задаче требовалось прибавить 5, а 4 мы уже добавили, то не хватает всего единички. Поэтому выходит:
- 6+5 – это 6+4 и еще 1;
- 6+4=10;
- последнюю единицу записываем вместо 0, получается 11.
Сначала ребенку ничего не будет понятно, но спустя время он усвоит основный принцип работы с десятками. Облегчить задачку можно с помощью наглядного упражнения:
- попросите отсчитать 10 конфет и сложить в одну миску;
- теперь нужно отложить еще 7 лакомств в другую емкость;
- под каждой миской следует разместить карточку с изображением числа, соответствующему количеству конфет;
- попросите малыша сложить все вместе и сказать, сколько конфет получилось;
- для большего понимания объясните, что 10 в двузначном счете обозначают 1;
- правильный вариант: ребенок пишет единицу и 7 рядом, что означает 17.
Подобные примеры можно проводить и с большим количеством предметов. Например, дошкольник должен знать, что 32 – это 3 десятка плюс еще 2 единицы.
Как помочь ребенку?
Числа от 2 до 10 в повседневной жизни встречаются очень часто, а обучение счету – во многом креативный процесс. Помочь с освоением цифр просто даже без покупки специальных приспособлений:
- Малышу нравится спорт? Подсчитывайте с ним количество забитых мячей в ворота.
- Ребенок любит природу? Разглядывайте деревья на лужайке, сравнивайте, с какой стороны их больше, а с какой меньше.
- Чадо постоянно что-то рисует? Предложите ему изобразить определенное количество предметов на листе. Увлекается лепкой? Попросите смастерить фигурку с 3 лапами, 2 хвостами и 1 ухом.
- Хитрите. В течение дня спрашивайте у малыша «если я возьму у тебя одно печенье, то сколько у тебя останется?» и подобное.
В комфортной игровой атмосфере ребенок быстро разберется с составом всех чисел и будет считать 1 2 3 4 5 и до десятка правильно.
Если вашему малышу плохо дается устный счет, то можно предположить проблемы с мышлением, вниманием или недостатком концентрации. Направить энергию в нужное русло позволят игры от BrainApps. Сервис предлагает свыше 100 тренажеров, позволяющих развивать и совершенствовать интеллектуальные способности. Зарегистрировавшись на сайте, выберите необходимую категорию игр:
- Для улучшения памяти. Тренажеры типа «Числовой охват», «Запоминай и прокликай» и «Найди пару» разовьют объем памяти, улучшат ее точность и усовершенствуют объем внимания.
- Для развития мышления. Игры «Фруктовая математика», «Сравнение цифр по памяти» и «Перемещения» улучшат зрительную и пространственную память, разовьют логику и научат ребенка быстро находить ответы на поставленные вопросы.
- Для совершенствования внимания. Тренажеры «Найди фигуру», «Космос» и «Концентрация» помогут малышу лучше концентрироваться и направлять внимание в нужное русло.
Совместные игры с дошкольником онлайн – не только увлекательное, но и полезное хобби. Чадо сможет развиваться, достигать поставленных целей и соревноваться с вами или сверстниками. Все, что нужно – подключение к сети и свободное время.
Рекомендации родителям
Понимая важность объяснения состава чисел, старайтесь придерживаться предложенных рекомендаций, которые помогут дошколенку спокойно освоить и запомнить новую информацию:
- У всех бывают кризисы, но нужно помнить, что это временное явление.
- Давайте больше свободы. Часто требования родителей не соответствуют возможностям малыша. Подумайте, все ли ваши запреты обоснованы.
- Учитывайте детское мнение. Нужно четко понимать, что у дошкольника есть собственное мнение и суждения. Постарайтесь их принять.
- Заставлять малоэффективно. Назидание и приказной тон во время занятий не дадут положительного эффекта. Если малышу не хочется учить числа и примеры с цифрами 1 2 3 кажутся ему страшными, узнайте, в чем причина. Объясните, зачем и почему нужно учиться.
- Терпение и оптимизм – ваши лучшие спутники. Хорошее настроение, атмосфера любви, поддержки и взаимопонимания направляют стремления в нужное русло.
Также рекомендуется придерживаться стратегии позитивного оценивания малыша:
- Демонстрируйте свое доброжелательное отношение.
- Совместно находите и анализируйте ошибки.
- Обсуждайте варианты и способы устранения ошибок вместе.
- Поддерживайте и выражайте уверенность в том, что у него обязательно все получится.
Не забывайте закреплять результаты и включать в программу обучения развивающие мультфильмы и другие видео.
iqclub.ru
Заполни пустые места
Для того, чтобы состав чисел от 1 до 10 действительно отложился в детских головках, предлагаю скачать карточки-задания на состав числа
В детском саду в старших группах есть занятия по подготовке детей к школе. Малышей учать считать и решать простенькие примеры и задачи.
А для этого детям дают задание запомнить состав числа.
Скажем, 5 это 1 плюс 4, или 2 плюс 3, или 3 плюс 2, или 4 плюс 1.
Малыши заучивают это как стишок или скороговорку, зачастую просто не вникая или не понимая смысла.
Для того, чтобы состав чисел от 1 до 10 действительно отложился в детских головках, предлагаю скачать карточки-задания на состав числа. Для того, чтобы заполнить пустые места надо подумать.
Вот первые из них.
Скачать карточки одним архивом: sostav. rar
ot2do6.ru
Статистика состава домохозяйства – Объяснение статистики
Увеличение числа домохозяйств, состоящих из одиноких взрослых
В 2021 году в ЕС проживало 197 миллионов домохозяйств, в среднем 2,2 члена на домохозяйство.
На рисунке 1 показано, что самый большой средний размер домохозяйства в 2021 г. был зарегистрирован в Словакии (2,9 члена), за ней следуют Хорватия (2,8 члена) и Португалия (2,7 члена), а наименьший размер был зарегистрирован в Швеции, Финляндии и Эстонии (каждая с 1,9члены).
В период с 2009 по 2021 год в большинстве государств-членов ЕС (23 из 27) было зафиксировано снижение среднего количества людей на домохозяйство. Наибольшее снижение с 2009 года было зафиксировано в Эстонии (с 2,4 до 1,9 члена), Латвии (с 2,6 до 2,1 члена) и на Мальте (с 2,9 до 2,5 члена).
Рисунок 1: Среднее количество людей на домохозяйство, 2009 и 2021 гг.0,5% в период с 2009 по 2021 год. Однако домохозяйства, состоящие из одного взрослого (т. е. домохозяйства, состоящие только из одного взрослого, живущие с детьми или без детей) росли намного быстрее, на 27,4 % за тот же период (см. Рисунок 2). Этот рост еще более заметен в домохозяйствах, состоящих из одиноких взрослых без детей (+28,5 %). См. Рисунок 3, чтобы сравнить данные за 2009 и 2021 годы по наличию детей.
В домохозяйствах, состоящих из пары (согласно юридическому статусу в браке или фактических отношениях), с детьми или без детей, зарегистрирован более медленный рост на 4,1 % с 2009 года. . Это увеличение обусловлено увеличением числа пар, живущих без детей.
Кроме того, количество домохозяйств с двумя или более взрослыми (ни один из которых не живет вместе как пара), также называемых в этой статье домохозяйствами «другого типа», сократилось на 5,6 % в период с 2009 по 2021 год в ЕС. Сокращение в этом типе домохозяйств еще выше для домохозяйств с детьми (-11,8 %), чем для бездетных (-3,1 %).
Рисунок 2: Домохозяйства по типу, ЕС, 2009–2021 гг.
Источник : Евростат (lfst_hhnhtych)
Рисунок 3: Домохозяйства по типу и наличию детей, ЕС, 2009 и 2021 гг.
Источник : Евростат (lfst_hhnhtych) между ребенком и взрослым. В соответствии с международными стандартами ребенком считается член домохозяйства в возрасте до 18 лет. Соответственно, взрослый – это лицо в возрасте 18 лет и старше.
Относительные различия в домохозяйствах мужчин и женщин, молодежи и пожилых людей
Что касается изменения численности взрослого населения в период с 2009 по 2021 год, то в зависимости от возраста и пола возникли разные закономерности (см. рис. 4).
Что касается одиноких взрослых, то как мужчины, так и женщины зафиксировали большее число одиноких людей во всех возрастных группах в 2021 году по сравнению с 2009 годом. Рост превысил 15 % для всех категорий.
Как среди мужчин, так и среди женщин доля одиноких взрослых росла быстрее, чем доля взрослых, живущих в паре. Этот вывод наблюдается для мужчин и женщин всех возрастных групп, за исключением женщин в возрасте 65 лет и старше, для которых более высокий рост был зарегистрирован для тех, кто живет в паре (+18,7 % для одиноких женщин в возрасте 65 лет и старше против +33,2 % для женщин). ровесники, проживающие в паре).
Число взрослых мужчин, живущих в одиночестве, увеличивалось относительно быстрее, чем число взрослых женщин, независимо от возрастной группы (хотя в 2021 году мужчин, живущих в одиночестве, по-прежнему было меньше, чем женщин в большинстве возрастных групп, см. ниже). Мужчины в возрасте 55–64 лет испытали наиболее значительный рост в период с 2009 по 2021 год — на 72,4 %. Для сравнения, с 2009 года для женщин той же возрастной категории было зарегистрировано увеличение на 34,4 %. Количество одиноких пожилых мужчин в возрасте 65 лет и старше увеличилось на 50,4 % в 2021 году по сравнению с 2009 годом., в то время как соответствующее увеличение для женщин в этом возрасте составило 18,7 %.
Рисунок 4: Темпы роста взрослого населения в ЕС по типу домохозяйства, полу и возрасту
Источник : Евростат (lfst_hhindws)
взрослых, проживающих в одиночестве, в паре или в домохозяйстве другого типа (т. е. с другим взрослым, но не в паре или с несколькими взрослыми) в зависимости от пола и возраста (см. рис. 5).
Более 40 % взрослых в возрасте 18 лет и старше жили в паре (42,3 % женщин и 45,6 % мужчин соответственно). Более трети женщин и мужчин проживали в домохозяйствах другого типа (34,2 % и 35,5 % соответственно). Одинокие женщины составляли почти четверть от общего числа женщин (23,6 %), в то время как одинокие мужчины составляли почти пятую часть от общего числа мужчин (18,9 %).
Среди молодых людей в возрасте от 18 до 24 лет 82,4 % мужчин по сравнению с 76,4 % женщин жили в домашнем хозяйстве другого типа (например, с родителями/семьей или в качестве соседей по квартире). Еще одно важное отличие среди молодежи состоит в том, что 9В паре жили 0,9% женщин, в то время как у мужчин этой возрастной категории 4,8%.
В ЕС 39,8 % женщин в возрасте 65 лет и старше жили одни по сравнению с 20,0 % мужчин той же возрастной категории. Около 4 из 10 женщин (40,1 %) и 6 из 10 мужчин (61,7 %) жили как пара, вероятно, потому, что женщины в среднем живут дольше.
Рисунок 5: Доля взрослого населения ЕС по типу домохозяйства, полу и возрасту, 2021 г.
Источник : Евростат (lfst_hhindws)
Наличие и количество детей
В 2021 году в ЕС зафиксировано увеличение на 14,5 % домохозяйств без детей и уменьшение на 3,4 % домохозяйств с детьми по сравнению с 2009 годом (см. Рисунок 6).
На национальном уровне в большинстве стран (23 из 26, по которым имеются данные) в период с 2009 по 2021 год было зарегистрировано увеличение числа домохозяйств без детей. Самый высокий рост был зарегистрирован на Мальте (+65,7 %), за ней следует Люксембург (+ 41,8 %), Кипр (+39,4 %) и Швеция (+35,8 %). Только Словакия (-1,6 %), Болгария (-1,9 %).%) и Греции (-7,4 %) в 2021 году было меньше домохозяйств без детей, чем в 2009 году.
Что касается домохозяйств с детьми, то в странах-членах ЕС наблюдается относительно иная тенденция. Количество домохозяйств с детьми сократилось в 16 странах, при этом наибольшее снижение произошло в Литве (-21,7 %). В Словакии он оставался стабильным и увеличился в 9 странах, включая Люксембург и Мальту, где он увеличился более чем на 10 % (+13,7 % и +11,5 % соответственно).
Рисунок 6: Темпы роста домохозяйств с детьми и без детей
Источник : Евростат (lfst_hhnhtych)
Распределение домохозяйств в 2021 году показывает, что доля домохозяйств, в которых живут дети, может значительно различаться в разных странах (см. рис. 7).
На уровне ЕС примерно в четверти домохозяйств (24,4 %) есть дети. В верхней части шкалы дети жили более чем в 30 % домохозяйств в Ирландии, Словакии, Кипре, Португалии и Румынии. Напротив, дети были обнаружены менее чем в 20 % домохозяйств в Германии и Финляндии.
Около двух третей (64,0 %) домохозяйств с детьми на уровне ЕС состояли из пар. В большинстве стран ЕС это наиболее распространенный тип домохозяйства с детьми. Швеция, Финляндия, Нидерланды и Греция зафиксировали самые высокие доли, при этом пары составляют более 70 % от общего числа домохозяйств с детьми. Болгария и Латвия показали самые низкие доли: менее 50 % домохозяйств с детьми составляют пары.
Самые высокие доли родителей-одиночек среди домохозяйств с детьми в ЕС были обнаружены в Эстонии, Дании, Литве и Латвии (более 20 %). В Словакии, Хорватии, Греции и Словении была самая низкая доля родителей-одиночек (во всех зарегистрирован процент менее 5 %). Во всем Европейском Союзе родители-одиночки составляли 12,6 % домохозяйств с детьми.
Другие типы домохозяйств составляют менее одной четверти (23,4 %) от общего числа домохозяйств с детьми на уровне ЕС. Эта доля сильно различалась среди стран ЕС: от более 35 % в Хорватии, Болгарии, Румынии и Словакии до менее 15 % в Финляндии, Эстонии, Дании и Швеции.
Среди домохозяйств без детей в ЕС в 2021 году около половины из них состояли из одиноких взрослых (47,4 %), около одной трети составляли пары (32,4 %) и одну пятую составляли домохозяйства с двумя взрослыми (которые не были парой) или более (20,2 %).
Рисунок 7: Домохозяйства по типу и наличию детей, 2021 г.
Источник : Евростат (lfst_hhnhtych)
Среди домохозяйств с детьми наиболее распространены домохозяйства с одним ребенком. В 2021 году в ЕС почти в половине семей с детьми был только 1 ребенок (49,4 %). В Болгарии, Португалии, Румынии и Литве самая высокая доля домохозяйств с 1 ребенком — более 55 %. Напротив, самые низкие доли были в Ирландии и Швеции, где домохозяйства с 1 ребенком составляли менее 40 % домохозяйств с детьми (см. Рисунок 8).
На уровне ЕС в 2021 г. 38,6 % домохозяйств с детьми имели 2 детей. Домохозяйства с двумя детьми чаще всего встречаются в Швеции, Нидерландах и Словении, что составляет 47,0 %, 44,0 % и 42,4 %, соответственно, домохозяйств с детьми в этих странах.
В ЕС 12,0 % домохозяйств с детьми в 2021 г. состояли из домохозяйств с 3 и более детьми. Ирландия, Финляндия, Хорватия, Бельгия, Франция, Швеция, Греция и Нидерланды зафиксировали самую высокую долю домохозяйств с 3 детьми и более, все выше 15 %. С другой стороны шкалы, на Мальте, в Литве, Чехии, Испании, Италии, Португалии и Болгарии менее 1 из 10 семей с детьми имеют 3 или более детей.
Рисунок 8: Домохозяйства по количеству детей, 2021
Источник : Eurostat (LFST_HHNHTYCH)
Источник Данные для таблиц и графиков
- Скачать файл Excel – Домохозяйства 2021 ДАННЫ источники
Все статистические данные, представленные в этой статье, получены из Обследования рабочей силы Европейского Союза (EU-LFS).
EU-LFS является крупнейшим европейским выборочным обследованием домохозяйств, предоставляющим ежеквартальные и годовые результаты участия в трудовой деятельности людей в возрасте 15 лет и старше. Он охватывает жителей частных домохозяйств и не включает жителей коллективных домохозяйств.
В результаты не включаются призывники, проходящие военную или общественную службу. EU-LFS основан на одних и тех же целевых группах населения и использует одни и те же определения во всех странах, что означает, что результаты сопоставимы между странами.В рамках специальной темы «Статистика домохозяйств» ОРС ЕС в настоящее время охватывает статистику состава домохозяйств, а также количества и размера домохозяйств.
Отчетный период: Годовые результаты получаются как средние значения за четыре квартала года.
Охват: Результаты EU-LFS в настоящее время охватывают все государства-члены Европейского Союза, государства-члены ЕАСТ Исландию, Норвегию, Швейцарию, а также страны-кандидаты Черногорию, Северную Македонию, Сербию и Турцию.
Для Кипра обследование охватывает только районы Кипра, контролируемые правительством Республики Кипр.
Тем не менее, данные о домохозяйствах EU-LFS недоступны для Исландии, Норвегии и Швейцарии.Европейские агрегаты: ЕС относится к сумме государств-членов ЕС. Если данные по стране недоступны, при расчете соответствующих агрегатов учитываются данные по той же стране за самый последний доступный период. Такие случаи указаны.
Определения
Ребенком считается член домохозяйства в возрасте до 18 лет.
Примечания по странам
В Германия , начиная с первого квартала 2020 года, обследование рабочей силы (ОРС) является частью новой системы интегрированных обследований домохозяйств. Технические проблемы и COVID-19Кризис оказал большое влияние на процессы сбора данных в 2020 году, что привело к низкой доле ответивших и необъективной выборке. Дополнительную информацию см. здесь.
В Нидерланды данные ОРС за 2021 год по-прежнему собираются с использованием скользящей контрольной недели вместо фиксированной контрольной недели, т.
е. опрошенных спрашивают о ситуации за неделю до интервью, а не за заранее выбранную неделю.Испания и Франция оценили привязанность к работе и включили в число занятых тех, кто в отчетную неделю имел неизвестную продолжительность отсутствия, но рассчитывал вернуться на ту же работу, как только это позволят меры по охране здоровья.
Временной ряд
Регламент (ЕС) 2019/1700 вступил в силу 1 января 2021 г. и привел к разрыву временного ряда EU-LFS для нескольких государств-членов ЕС.
Дополнительная методологическая информация
Дополнительную информацию о EU-LFS можно найти в онлайн-публикации «Обзор рабочей силы ЕС», которая включает восемь статей о технических и методологических аспектах обследования. Методология EU-LFS, действующая с момента сбора данных в 2021 году, описана в методологии, начиная с 2021 года. Подробную информацию о списках кодирования, пояснительных примечаниях и классификациях, используемых с течением времени, можно найти в документации.
Контекст
В дополнение к Обследованию рабочей силы (LFS) статистика доходов и условий жизни ЕС (EU-SILC) также является источником статистических данных о домохозяйствах. EU-SILC — это многоцелевой инструмент, ориентированный в основном на доход. Однако также собирается информация о жилищных условиях, социальной изоляции, занятости и образовании.
2022 Кодекс этики и стандарты практики
Поиск Кодекса этики Кодекс этики Обучение
Кодекс этики и стандарты практики Национальной ассоциации риэлторов®
Вступает в силу с 1 января 2022 г.
Преамбула
Обязанности клиентов Клиенты
Обязанности перед общественностью
Обязанности РИЭЛТОРОВ®
Пояснительные примечания REALTOR-ASSOCIATE®s.Хотя Кодекс этики устанавливает обязательства, которые могут быть выше тех, которые предусмотрены законом, в любом случае, когда Кодекс этики и закон противоречат друг другу, обязательства закона должны иметь преимущественную силу.
Преамбула
Земля под всем. От его мудрого использования и широкого распределения собственности зависит выживание и рост свободных институтов и нашей цивилизации. РИЭЛТОРЫ® должны признать, что интересы нации и ее граждан требуют наиболее эффективного использования земли и самого широкого распределения собственности на землю. Они требуют создания надлежащего жилья, строительства функционирующих городов, развития производственной промышленности и ферм, а также сохранения здоровой окружающей среды.
Такие интересы налагают обязательства, выходящие за рамки обычной торговли. Они налагают серьезную социальную ответственность и патриотический долг, которому РИЭЛТОРЫ® должны посвятить себя и к которому они должны усердно готовиться. Поэтому РИЭЛТОРЫ® усердно поддерживают и улучшают стандарты своего призвания и разделяют со своими коллегами-РИЭЛТОРАМИ® общую ответственность за его честность и честь.
Признавая и признавая свои обязательства перед клиентами, покупателями, общественностью и друг перед другом, РИЭЛТОРЫ® постоянно стремятся получать и оставаться в курсе вопросов, касающихся недвижимости, и, как знающие профессионалы, они охотно делятся плодами своего опыта и учиться с другими.
Они выявляют и предпринимают шаги путем обеспечения соблюдения настоящего Кодекса этики и оказания помощи соответствующим регулирующим органам для устранения практики, которая может нанести ущерб обществу или дискредитировать или обесчестить профессию в сфере недвижимости. РИЭЛТОРЫ®, непосредственно осведомленные о поведении, которое может нарушать Кодекс этики, связанное с незаконным присвоением средств или имущества клиента или заказчика, умышленной дискриминацией или мошенничеством, приводящим к существенному экономическому ущербу, доводят такие вопросы до сведения соответствующего Совета или Ассоциации РИЭЛТОРОВ. ®. (Изм. 1/00) Понимая, что сотрудничество с другими профессионалами в сфере недвижимости отвечает интересам тех, кто пользуется их услугами, РИЭЛТОРЫ® настаивают на эксклюзивном представлении клиентов; не пытаться получить несправедливое преимущество перед своими конкурентами; и они воздерживаются от нежелательных комментариев о других практикующих. В тех случаях, когда запрашивается их мнение или когда РИЭЛТОРЫ считают, что комментарий необходим, их мнение предлагается объективно, профессионально, без влияния каких-либо личных мотивов или потенциальной выгоды или выгоды.
Термин REALTOR® стал обозначать компетентность, справедливость и высокую честность, возникающие в результате приверженности высоким идеалам нравственного поведения в деловых отношениях. Никакое побуждение к прибыли и никакие инструкции со стороны клиентов никогда не могут оправдать отход от этого идеала.
В толковании этого обязательства РИЭЛТОРЫ® не могут руководствоваться более надежным руководством, чем то, которое передавалось из поколения в поколение, воплощенное в Золотом правиле: «Что бы вы ни хотели, чтобы другие поступали с вами, то и вы поступайте с ними». ».
Принимая этот стандарт как свой собственный, РИЭЛТОРЫ® обязуются соблюдать его дух во всей своей деятельности, независимо от того, осуществляется ли она лично, через партнеров или других лиц или с помощью технических средств, и вести свой бизнес в соответствии с принципами, изложенными ниже. (Изм. 07.01.)
Обязанности перед клиентами и заказчиками
Статья 1
Представляя покупателя, продавца, арендодателя, арендатора или другого клиента в качестве агента, РИЭЛТОРЫ® обязуются защищать и продвигать интересы их клиент.
Это обязательство перед клиентом является первичным, но оно не освобождает РИЭЛТОРОВ® от их обязанности честно относиться ко всем сторонам. Обслуживая покупателя, продавца, арендодателя, арендатора или другую сторону, не являющуюся агентом, РИЭЛТОРЫ® по-прежнему обязаны вести себя честно со всеми сторонами. (Изм. 01.01.) Стандарт практики 1-1
Риэлторы®, выступая в качестве доверителей в сделках с недвижимостью, по-прежнему несут обязательства, предусмотренные Кодексом этики. (Изм. 1/93)
Стандарт практики 1-2
Обязанности, налагаемые Кодексом этики, охватывают все действия и сделки, связанные с недвижимостью, независимо от того, осуществляются ли они лично, электронными средствами или любыми другими способами.
Обязанности, возлагаемые Кодексом этики, применимы независимо от того, действуют ли РИЭЛТОРЫ® в качестве агентов или в юридически признанном неагентском качестве, за исключением того, что любые обязанности, возложенные исключительно на агентов законом или нормативным актом, не должны возлагаться настоящим Кодексом этики на РИЭЛТОРОВ® действует в неагентском качестве.
В данном Этическом кодексе термин «клиент» означает лицо (лица) или организацию (организации), с которыми РИЭЛТОР® или фирма РИЭЛТОРА® состоит в агентских или юридически признанных неагентских отношениях; «клиент» означает сторону сделки с недвижимостью, которая получает информацию, услуги или преимущества, но не имеет договорных отношений с РИЭЛТОРОМ® или фирмой РИЭЛТОРА®; «потенциальный покупатель» означает покупателя, продавца, арендатора или арендодателя, который не является объектом представительских отношений с РИЭЛТОРОМ® или фирмой РИЭЛТОРА®; «агент» означает лицензиата в сфере недвижимости (включая брокеров и торговых партнеров), действующего в рамках агентских отношений, как это определено законодательством штата или нормативным актом; и «брокер» означает лицензиата в сфере недвижимости (включая брокеров и торговых партнеров), действующего в качестве агента или в юридически признанном неагентском качестве. (Принято 01/95, изменено 01/07)
Стандарт практики 1-3
РИЭЛТОРЫ®, пытаясь получить листинг, не должны намеренно вводить владельца в заблуждение относительно рыночной стоимости.
Standard of Practice 1-4
РИЭЛТОРЫ®, стремящиеся стать представителями покупателя/арендатора, не должны вводить покупателей или арендаторов в заблуждение относительно экономии или других преимуществ, которые могут быть получены благодаря использованию услуг РИЭЛТОРА®. (с изменениями от 1/93)
Стандарт практики 1–5
РИЭЛТОРЫ® могут представлять продавца/арендодателя и покупателя/арендатора в одной и той же сделке только после полного раскрытия информации и с информированного согласия обеих сторон. (Принято 1/93)
Стандарт практики 1-6
РИЭЛТОРЫ® должны подавать предложения и встречные предложения объективно и как можно быстрее. (Принято 1/93, изменено 1/95)
Стандарт практики 1-7
Выступая в качестве листинговых брокеров, РИЭЛТОРЫ® должны продолжать представлять продавцу/арендодателю все предложения и встречные предложения до закрытия или исполнения договора аренды, если только продавец/арендодатель не отказался от этого обязательства в письменной форме.
По письменному запросу сотрудничающего брокера, который представляет предложение брокеру по листингу, брокер по листингу должен предоставить, как только это будет практически возможно, письменное подтверждение сотрудничающему брокеру о том, что предложение было отправлено продавцу/арендодателю, или письменное уведомление о том, что продавец/арендодатель отказался от обязательства представить предложение. РИЭЛТОРЫ® не обязаны продолжать продавать недвижимость после того, как предложение было принято продавцом/арендодателем. РИЭЛТОРЫ® должны рекомендовать продавцам/арендодателям получить консультацию юриста до принятия последующего предложения, за исключением случаев, когда принятие зависит от расторжения ранее существовавшего договора купли-продажи или аренды. (Изм. 1/20) Стандарт практики 1-8
РИЭЛТОРЫ®, действующие в качестве агентов или посредников покупателей/арендаторов, должны представлять покупателям/арендаторам все предложения и встречные предложения до их принятия, но не обязаны продолжать показывать недвижимость своим клиентам после того, как предложение было принято, если иное не согласовано в письменной форме.
По письменному запросу брокера по листингу, который направляет встречное предложение брокеру арендатора покупателя, брокер покупателя/арендатора должен предоставить, как только это станет практически возможным, письменное подтверждение брокеру по листингу о том, что встречное предложение было направлено покупателей/арендаторов, или письменное уведомление о том, что покупатели/арендаторы отказались от обязательства по представлению встречного предложения. РИЭЛТОРЫ®, выступая в качестве агентов или посредников покупателей/арендаторов, должны рекомендовать покупателям/арендаторам получить консультацию юриста, если возникает вопрос о том, был ли расторгнут ранее существовавший договор. (Принято 1/93, изменено 1/22) Стандарт практики 1-9
Обязанность РИЭЛТОРОВ® сохранять конфиденциальную информацию (согласно определению закона штата), предоставленную их клиентами в ходе любых агентских отношений или неагентские отношения, признанные законом, продолжаются после прекращения агентских отношений или любых неагентских отношений, признанных законом.
РИЭЛТОРЫ® не должны сознательно, во время или после прекращения профессиональных отношений со своими клиентами:1) разглашать конфиденциальную информацию клиентов; или
2) использовать конфиденциальную информацию клиентов в ущерб клиентам; или
3) использовать конфиденциальную информацию клиентов в интересах РИЭЛТОРА® или третьих лиц, за исключением случаев, когда:
а) клиент дает согласие после полного раскрытия; или
б) РИЭЛТОРЫ® требуются по решению суда; или
c) у клиента есть намерение совершить преступление и информация необходима для предотвращения преступления; или
d) необходимо защитить РИЭЛТОРА® или сотрудников или партнеров РИЭЛТОРА® от обвинения в неправомерном поведении.
Информация о скрытых дефектах материалов не считается конфиденциальной информацией в соответствии с настоящим Кодексом этики. (Принято 01/93, изменено 1/01)
Стандарт практики 1-10
РИЭЛТОРЫ® должны, в соответствии с положениями и условиями их лицензии на недвижимость и их соглашения об управлении недвижимостью, компетентно управлять собственностью клиентов с должным учетом прав, безопасности и здоровья арендаторов и других лиц, законно находящихся в помещении.
(Принято 1/95, изменено 1/00) Стандарт практики 1-11
РИЭЛТОРЫ®, нанятые для обслуживания или управления собственностью клиента, должны проявлять должную осмотрительность и прилагать разумные усилия для ее защиты от разумно предсказуемых непредвиденных обстоятельств. и потери. ( Принято 1/95 )
Стандарт практики 1-12
При заключении договоров о листинге РИЭЛТОРЫ® должны сообщать продавцам/арендодателям:
1) политику компании РИЭЛТОРА® в отношении сотрудничества и суммы( s) любой компенсации, которая будет предложена субагентам, агентам покупателя/арендатора и/или брокерам, действующим в юридически признанных неагентских качествах;
2) тот факт, что агенты покупателя/арендатора или брокеры, даже если им компенсируют посредники листинга, или продавцы/арендодатели, могут представлять интересы покупателей/арендаторов; и
3) любой потенциал для того, чтобы листинговые брокеры выступали в качестве раскрытых двойных агентов, например агентов покупателя/арендатора.
(Принято 1/93, перенумеровано 1/98, изменено 1/03) Стандарт практики 1-13
При заключении договоров покупателя/арендатора РИЭЛТОРЫ® должны сообщить потенциальным клиентам:
1) РИЭЛТОР политика компании ® в отношении сотрудничества;
2) размер возмещения, подлежащего выплате клиентом;
3) возможность получения дополнительной или компенсационной компенсации от других маклеров, продавца или арендодателя или других сторон;
4) любая возможность для представителя покупателя/арендатора выступать в качестве раскрытого двойного агента, например, листингового брокера, субагента, агента арендодателя и т. д.; и
5) возможность того, что продавцы или представители продавцов не могут рассматривать существование, условия предложений как конфиденциальные, если конфиденциальность не требуется в соответствии с законом, нормативным актом или любым соглашением о конфиденциальности между сторонами. (Принято 1/93, перенумеровано 1/98, изменено 1/06)
Стандарт практики 1-14
Вознаграждения за подготовку оценок или других оценок не должны зависеть от суммы оценки или оценки.
(Принято 02.01.) Стандарт практики 1-15
РИЭЛТОРЫ® в ответ на запросы покупателей или сотрудничающих с ними брокеров должны, с одобрения продавцов, раскрывать наличие предложений по недвижимости. В тех случаях, когда раскрытие разрешено, РИЭЛТОРЫ® должны также раскрыть, если их спросят, были ли предложения получены лицензиатом листинга, другим лицензиатом в листинговой фирме или сотрудничающим брокером. (Принято 03.01, изменено 09.01)
Стандарт практики 1-16
РИЭЛТОРЫ® не должны получать доступ или использовать, а также разрешать или позволять другим получать доступ или использовать зарегистрированную или управляемую собственность на других условиях. чем те, которые разрешены владельцем или продавцом. (Принято 1/12)
Статья 2
РИЭЛТОРЫ® должны избегать преувеличения, искажения или сокрытия соответствующих фактов, касающихся имущества или сделки. Однако РИЭЛТОРЫ® не обязаны обнаруживать скрытые дефекты в собственности, давать советы по вопросам, выходящим за рамки их лицензии на недвижимость, или раскрывать факты, которые являются конфиденциальными в рамках агентских или неагентских отношений, как это определено Закон штата.
(Изм. 1/00) Стандарт практики 2-1
РИЭЛТОРЫ® обязаны выявлять и раскрывать неблагоприятные факторы, разумно очевидные лицу, имеющему опыт в тех областях, которые требуются их органом по лицензированию недвижимости. Статья 2 не налагает на РИЭЛТОРА обязательство быть экспертом в других профессиональных или технических дисциплинах. (с изменениями от 1/96)
Стандарт практики 2-2
(перенумерован как Стандарт практики 1-12 1/98)
Стандарт практики 2-3
(перенумерован как Стандарт практики 1-13 1/98)
Стандарт практики 2-4
вознаграждение в каком-либо документе, если только это не указание явно номинального вознаграждения.
Стандарт практики 2-5
Факторы, определяемые как «несущественные» в соответствии с законом или нормативно-правовым актом или на которые в законе или нормативно-правовом акте прямо ссылаются как на не подлежащие раскрытию, считаются несущественными для целей статьи 2.
(Принято 01/93) Статья 3
РИЭЛТОРЫ® должны сотрудничать с другими брокерами, за исключением случаев, когда сотрудничество не отвечает интересам клиента. Обязательство сотрудничать не включает в себя обязательство делить комиссионные, сборы или иным образом компенсировать другого брокера. (Изм. 1/95)
Стандарт практики 3-1
РИЭЛТОРЫ®, выступая в качестве эксклюзивных агентов или посредников продавцов/арендодателей, устанавливают условия предложений о сотрудничестве. Если это прямо не указано в предложениях о сотрудничестве, сотрудничающие брокеры не могут предполагать, что предложение о сотрудничестве включает предложение компенсации. Условия компенсации, если таковые имеются, должны быть установлены сотрудничающими брокерами до начала принятия предложения о сотрудничестве. (Изм. 1/99)
Стандарт практики 3-2
Любые изменения в компенсации, предлагаемой за совместные услуги, должны быть сообщены другому РИЭЛТОРУ® до того, как РИЭЛТОР® представит предложение о покупке/аренде недвижимости .
После того, как РИЭЛТОР® подал предложение о покупке или аренде недвижимости, брокер по листингу не может пытаться в одностороннем порядке изменить предложенную компенсацию в отношении этой совместной сделки. (поправка от 14 января) Стандарт практики 3-3
Стандарт практики 3-2 не запрещает листинговому брокеру и сотрудничающему брокеру заключать соглашение об изменении совместной компенсации. (Принято 01/94)
Стандарт практики 3-4
РИЭЛТОРЫ®, выступающие в качестве листинговых брокеров, имеют прямое обязательство раскрывать информацию о существовании договоренностей о двойной или переменной ставке комиссионных (т. выплачивается, если причиной продажи/аренды является фирма листингового брокера, и другая сумма комиссионных выплачивается, если продажа/аренда осуществляется благодаря усилиям продавца/арендодателя или сотрудничающего с ним брокера). Брокер по листингу должен, как только это станет практически возможным, сообщить о существовании таких договоренностей потенциальным сотрудничающим брокерам и должен, в ответ на запросы сотрудничающих брокеров, раскрыть разницу, которая может привести к совместной сделке или к продаже/аренде, которая возникает в результате усилия продавца/арендодателя.
Если сотрудничающий брокер является представителем покупателя/арендатора, представитель покупателя/арендатора должен раскрыть такую информацию своему клиенту до того, как клиент сделает предложение о покупке или аренде. (Изм. 1/02) Стандарт практики 3-5
Субагенты обязаны незамедлительно раскрывать все относящиеся к делу факты агенту принципала до, а также после заключения договора купли-продажи или аренды. (Поправка от 1/93)
Стандарт практики 3-6
РИЭЛТОРЫ® должны сообщать о наличии принятых предложений, включая предложения с неурегулированными непредвиденными обстоятельствами, любому брокеру, желающему сотрудничать. (Принято 5/86, изменено 1/04)
Стандарт практики 3-7
При запросе информации от другого РИЭЛТОРА® относительно собственности в соответствии с соглашением об управлении или листинге, РИЭЛТОРЫ® должны раскрывать свой статус РИЭЛТОРА® и то, являются ли их интересы личными или от имени клиента, и, если от имени клиента, их отношения с клиентом.
( с поправками от 1/11 )Стандарт практики 3-8
РИЭЛТОРЫ® не должны искажать информацию о наличии доступа для показа или осмотра выставленного на продажу имущества. (Изм. от 11.11.87)
Стандарт практики 3-9
РИЭЛТОРЫ® не должны предоставлять доступ к зарегистрированной недвижимости на условиях, отличных от тех, которые установлены владельцем или листинговым брокером. (Принято 1/10)
Стандарт практики 3-10
Обязанность сотрудничать, установленная в статье 3, относится к обязательству делиться информацией о выставленном на продажу имуществе и предоставлять имущество другим брокерам для показа потенциальным покупателям /арендодателям, когда это отвечает интересам продавцов/арендодателей. (Принято 1/1)
Стандарт практики 3-11
РИЭЛТОРЫ® не вправе отказываться от сотрудничества на основании расы, цвета кожи, религии, пола, инвалидности, семейного положения, национального происхождения, сексуальной ориентации, или гендерной идентичности.
(Принято 20 января) Статья 4
РИЭЛТОРЫ® не должны приобретать долю, покупать или делать предложения от самих себя, любых членов своих ближайших родственников, своих фирм или любого их члена или любых организаций, в которых они иметь какую-либо долю собственности, какое-либо недвижимое имущество, не сообщая о своем истинном положении владельцу, его агенту или брокеру. При продаже имущества, которым они владеют или в котором они имеют какую-либо заинтересованность, РИЭЛТОРЫ® должны сообщить о своей собственности или заинтересованности в письменной форме покупателю или представителю покупателя. (Изм. 1/00)
Стандарт практики 4-1
Для защиты всех сторон раскрытие информации, требуемое Статьей 4, должно быть в письменной форме и предоставлено РИЭЛТОРАМИ® до подписания любого контракта. (Принято 02/86)
Статья 5
РИЭЛТОРЫ® не берут на себя обязательство предоставлять профессиональные услуги в отношении имущества или его стоимости, если у них есть настоящий или предполагаемый интерес, если такой интерес специально не раскрыт всем заинтересованным сторонам.
Статья 6
РИЭЛТОРЫ® не должны принимать никаких комиссий, скидок или прибыли от расходов, сделанных для их клиента, без ведома и согласия клиента.
Рекомендуя продукты или услуги в сфере недвижимости (например, страхование домовладельца, гарантийные программы, ипотечное финансирование, страхование титула и т. д.), РИЭЛТОРЫ® должны раскрывать клиенту или заказчику, которому делается рекомендация, любые финансовые выгоды или сборы, другие РИЭЛТОР® или фирма РИЭЛТОРА® может получить непосредственный результат такой рекомендации. (Изм. 1/99)
Стандарт практики 6-1
РИЭЛТОРЫ® не должны рекомендовать или предлагать клиенту или покупателю пользоваться услугами другой организации или хозяйствующего субъекта, в которых они имеют прямую заинтересованность, не раскрывая такой интерес на момент рекомендации или предложения. (Изм. от 5/88)
Статья 7
В сделке РИЭЛТОРЫ® не должны принимать компенсацию более чем от одной стороны, даже если это разрешено законом, без раскрытия информации всем сторонам и информированного согласия РИЭЛТОРА®’ s клиент или клиенты.
(Изм. 1/93) Статья 8
РИЭЛТОРЫ® должны хранить на специальном счете в соответствующем финансовом учреждении, отдельно от собственных средств, денежные средства, поступающие в их распоряжение в доверительное управление для других лиц, такие как условное депонирование, траст средства, деньги клиентов и другие подобные предметы.
Статья 9
РИЭЛТОРЫ®, для защиты всех сторон, должны по возможности гарантировать, что все соглашения, связанные со сделками с недвижимостью, включая, помимо прочего, соглашения о листинге и представительстве, договоры купли-продажи и аренды, четко и ясно изложены в письменной форме. понятным языком, выражающим конкретные условия, обязанности и обязательства сторон. Копия каждого соглашения должна быть предоставлена каждой стороне таких соглашений после их подписания или парафирования. (Изм. 04.01.)
Стандарт практики 9-1
Для защиты всех сторон РИЭЛТОРЫ® должны проявлять разумную осторожность, чтобы обеспечить актуальность документов, касающихся покупки, продажи или аренды недвижимости.
путем использования письменных расширений или поправок. (Изм. 1/93) Стандарт практики 9-2
При оказании помощи или содействии клиенту или покупателю в установлении договорных отношений (например, листинг и представительские соглашения, договоры купли-продажи, аренды и т. д.) в электронной форме, РИЭЛТОРЫ ® прилагает разумные усилия для разъяснения характера и раскрытия конкретных условий договорных отношений, которые устанавливаются до их согласования с договаривающейся стороной. (Принято 07.01.)
Обязанности перед обществом
Статья 10
РИЭЛТОРЫ® не должны отказывать в равных профессиональных услугах любому лицу по причинам расы, цвета кожи, религии, пола, инвалидности, семейного положения, национального происхождения, сексуальная ориентация или гендерная идентичность. РИЭЛТОРЫ® не должны быть участниками какого-либо плана или соглашения о дискриминации лица или лиц по признаку расы, цвета кожи, религии, пола, инвалидности, семейного положения, национального происхождения, сексуальной ориентации или гендерной идентичности.
(Изм. 1/14) РИЭЛТОРЫ® в своей практике работы с недвижимостью не должны дискриминировать какое-либо лицо или лица по признаку расы, цвета кожи, религии, пола, инвалидности, семейного положения, национального происхождения, сексуальной ориентации. или гендерной идентичности. (Изм. 1/14)
Стандарт практики 10-1
При продаже или аренде жилья РИЭЛТОРЫ® не должны добровольно предоставлять информацию о расовом, религиозном или этническом составе любого района, а также однако в любой деятельности, которая может привести к паническим продажам, РИЭЛТОРЫ® могут предоставлять другую демографическую информацию. (Принято 01/94, изменено 01/06)
Стандарт практики 10-2
Если РИЭЛТОРЫ не участвуют в продаже или аренде жилья, они могут предоставлять демографическую информацию, связанную с недвижимостью, сделкой или профессиональным назначением. стороне, если такая демографическая информация (а) рассматривается РИЭЛТОРОМ® как необходимая для оказания помощи или завершения, в соответствии со статьей 10, сделки с недвижимостью или профессионального задания и (б) получена или получена от признанный, надежный, независимый и беспристрастный источник.
Источник такой информации и любые добавления, исключения, модификации, толкования или другие изменения должны быть раскрыты с достаточной степенью детализации. (Принято 01/05, перенумеровано 1/06) Стандарт практики 10-3
РИЭЛТОРЫ® не должны печатать, демонстрировать или распространять какие-либо заявления или рекламу в отношении продажи или аренды недвижимости, которые указывают на какие-либо предпочтения, ограничения или дискриминация по признаку расы, цвета кожи, религии, пола, инвалидности, семейного положения, национального происхождения, сексуальной ориентации или гендерной идентичности. (принят 1/94, перенумерован 1/05 и 1/06, изменен 1/14)
Стандарт практики 10-4
Как используется в статье 10, «практика найма в сфере недвижимости» относится к работникам и независимым подрядчикам, предоставляющим услуги, связанные с недвижимостью, а также к административному и канцелярскому персоналу, непосредственно поддерживающему этих лиц.
(Принято 1/00, перенумеровано 1/05 и 1/06) Стандарт практики 10-5
РИЭЛТОРЫ® не должны использовать оскорбительные высказывания, разжигание ненависти, эпитеты или оскорбления на основе расы, цвета кожи, религии, пол, инвалидность, семейное положение, национальность, сексуальная ориентация или гендерная идентичность. (Принято и вступило в силу 13 ноября 2020 г.)
Статья 11
Услуги, которые РИЭЛТОРЫ® предоставляют своим клиентам и покупателям, должны соответствовать стандартам практики и компетентности, которые разумно ожидаются в конкретных областях недвижимости, в которых они работают. привлекать; в частности, брокерские услуги в сфере жилой недвижимости, управление недвижимостью, брокерские операции с коммерческой и промышленной недвижимостью, брокерские операции с землей, оценка недвижимости, консультирование по вопросам недвижимости, синдикация недвижимости, аукционы недвижимости и международная недвижимость.
РИЭЛТОРЫ® не должны брать на себя обязательство предоставлять специализированные профессиональные услуги в отношении типа собственности или услуги, которые находятся за пределами их компетенции, если они не привлекут помощь лица, компетентного в отношении таких видов собственности или услуги, или если факты полностью не подтверждены.
раскрывается клиенту. Любые лица, нанятые для оказания такой помощи, должны быть обозначены для клиента, и их вклад в задание должен быть изложен. (Изм. 1/10) Standard of Practice 11-1
Когда РИЭЛТОРЫ® готовят заключения о стоимости или цене недвижимого имущества, они должны:
1) знать тип оцениваемого имущества,
2) иметь доступ к информации и ресурсам, необходимым для сформулировать точное заключение и
3) ознакомиться с местностью, где находится объект недвижимости
, если об отсутствии какого-либо из них не сообщается стороне, запрашивающей заключение заранее.
Когда заключение о стоимости или цене готовится не в целях листинга или для оказания помощи потенциальному покупателю в формулировании предложения о покупке, мнение должно включать следующее, если сторона, запрашивающая заключение, не требует отчета определенного типа или иного набор данных:
1) идентификация предмета собственности
2) дата подготовки
3) установленная стоимость или цена
4) ограничивающие условия, включая заявления о целях(ях) и предполагаемом(ых) пользователе(ях)
5) любые существующие или предполагаемый интерес, в том числе возможность представления продавца/арендодателя или покупателей/арендаторов
6) основание для заключения, включая применимые рыночные данные
7) если заключение не является оценкой, заявление на этот счет
8) раскрытие информации о том, проводился ли и когда физический осмотр объекта недвижимости снаружи
9) раскрытие информации о том, проводился ли и когда физический осмотр интерьера имущества
10) раскрытие информации о наличии у РИЭЛТОРА каких-либо конфликтов интерес (Изм.
1/14) Стандарт практики 11-2
Обязательства Кодекса этики в отношении дисциплин, связанных с недвижимостью, кроме оценки, должны толковаться и применяться в соответствии со стандартами компетентности и практики, которые клиенты и общественность обоснованно требуют защиты своих прав и интересов, учитывая сложность сделки, доступность экспертной помощи, а также, если РИЭЛТОР® является агентом или субагентом, обязанности доверенного лица. (Принято 01/95)
Стандарт практики 11-3
Когда РИЭЛТОРЫ® предоставляют консультационные услуги клиентам, которые включают советы или консультации за плату (не за комиссию), такие консультации должны предоставляться объективно и гонорар не должен зависеть от существа совета или совета. Если в дополнение к консультационным услугам должны быть оказаны брокерские или транзакционные услуги, по предварительному согласованию между клиентом и РИЭЛТОРОМ® может быть выплачена отдельная компенсация. (Принято 01/96)
Стандарт практики 11-4
Компетенция, требуемая Статьей 11, относится к услугам, заключенным по контракту между РИЭЛТОРАМИ® и их клиентами или заказчиками; обязанности, прямо предусмотренные Кодексом этики; и обязанности, возложенные законом или постановлением.
(Принято 02.01) Статья 12
РИЭЛТОРЫ® должны быть честными и правдивыми в своих сообщениях о недвижимости и должны представлять правдивую картину в своих рекламных, маркетинговых и других представлениях. РИЭЛТОРЫ® должны обеспечить, чтобы их статус профессионалов в сфере недвижимости был очевиден в их рекламных, маркетинговых и других заявлениях, и чтобы получатели всех сообщений о недвижимости были уведомлены или были уведомлены о том, что эти сообщения исходят от специалиста по недвижимости. (Изм. 08.01.)
Стандарт практики 12-1
РИЭЛТОРЫ® не должны заявлять, что их брокерские услуги клиенту или покупателю бесплатны или доступны для их клиентов бесплатно, за исключением случаев, когда РИЭЛТОР® не получит финансовая компенсация из любого источника за эти услуги. (Изменен 22 января)
Стандарт практики 12-2
(Удален 20 января)
Стандарт практики 12-3
Предложение премий, призов, списка товаров или других стимулов , продажа, покупка или аренда сами по себе не являются неэтичными, даже если получение выгоды зависит от листинга, продажи, покупки или аренды через РИЭЛТОРА, делающего предложение.
Тем не менее, РИЭЛТОРЫ® должны проявлять осторожность и искренность в любой такой рекламе или других публичных или частных заявлениях, чтобы любая сторона, заинтересованная в получении или иной выгоде от предложения РИЭЛТОРА®, имела четкое, полное и заранее понимание всех условий. предложения. Предложение любых поощрений для ведения бизнеса регулируется ограничениями законодательства штата и этическими обязательствами, установленными любым применимым Стандартом практики. (Изм. 1/95) Стандарт практики 12-4
РИЭЛТОРЫ® не должны продавать/сдавать в аренду или рекламировать недвижимость без разрешения. Выступая в качестве листинговых брокеров или субагентов, РИЭЛТОРЫ® не должны указывать цену, отличную от той, которая была согласована с продавцом/арендодателем. (Изм. от 1/93)
Стандарт практики 12-5
РИЭЛТОРЫ® не должны рекламировать и разрешать любому лицу, нанятому ими или аффилированному с ними, рекламировать услуги в сфере недвижимости или выставленную на продажу собственность на любом носителе (например, в электронном виде, в печатной , радио, телевидение и т.
д.) без раскрытия названия фирмы этого РИЭЛТОРА® разумным и очевидным образом либо в рекламе, либо в электронной рекламе через ссылку на дисплей со всеми необходимыми раскрытиями. (Принято 11 ноября 1986 г., изменено 1 июня) Стандарт практики 12-6
РИЭЛТОРЫ®, рекламируя незарегистрированную недвижимость для продажи/аренды, в которой они имеют долю владения, должны раскрывать свой статус обоих владельцев. /арендодателей, а также в качестве РИЭЛТОРОВ® или лицензиатов в сфере недвижимости. (Изм. 1/93)
Стандарт практики 12-7
Только РИЭЛТОРЫ®, которые участвовали в сделке в качестве листингового брокера или сотрудничающего брокера (торгового брокера), могут заявлять, что «продали» недвижимость. Перед закрытием сотрудничающий брокер может размещать знак «Продано» только с согласия листингового брокера. (Изм. 1/96)
Стандарт практики 12-8
Обязательство представлять достоверную картину в представлениях общественности включает информацию, представленную, предоставленную или отображенную на веб-сайтах РИЭЛТОРОВ®.
РИЭЛТОРЫ® должны прилагать разумные усилия для обеспечения актуальности информации на своих веб-сайтах. Когда становится очевидным, что информация на веб-сайте РИЭЛТОРА® больше не является актуальной или точной, РИЭЛТОРЫ® должны незамедлительно предпринять корректирующие действия. (Принят 07.01) Standard of Practice 12-9
Веб-сайты фирм REALTOR® должны раскрывать название фирмы и состояние(я) лицензии разумным и очевидным образом.
Веб-сайты РИЭЛТОРОВ® и лицензиатов, не являющихся членами, аффилированных с фирмой РИЭЛТОРов®, должны раскрывать название фирмы и состояние(я) лицензии этого РИЭЛТОРА® или лицензиата, не являющегося членом, разумным и очевидным образом. (Принято 07.01)
Стандарт практики 12-10
Обязанность РИЭЛТОРОВ® представлять реальную картину в своей рекламе и представлениях для общественности включает интернет-контент, изображения, а также URL-адреса и доменные имена, которые они используют, и запрещает РИЭЛТОРАМ®:
1) участие в обманном или несанкционированном создании веб-сайтов брокеров по недвижимости;
2) манипулирование (например, представление контента, разработанного другими) листингом и другим контентом любым способом, который приводит к вводящим в заблуждение результатам;
3) ввод в заблуждение с использованием метатегов, ключевых слов или других средств/методов для направления, управления или перенаправления интернет-трафика; или
4) представление контента, разработанного другими лицами, без указания авторства или без разрешения; или
5) введение потребителей в заблуждение иным образом, включая использование вводящих в заблуждение изображений.
(Принято 07.01, изменено 18.01) Стандарт практики 12-11
РИЭЛТОРЫ®, намеревающиеся поделиться или продать информацию о потребителях, собранную через Интернет, должны раскрыть эту возможность разумным и очевидным образом. (Принято 07.01)
Стандарт практики 12-12
РИЭЛТОРЫ® не должны:
1) использовать URL-адреса или доменные имена, которые не соответствуют действительности, или
2) регистрировать URL-адреса или доменные имена который, если бы его использовали, представил бы меньше, чем истинная картина. (Принято 01.08)
Стандарт практики 12-13
Обязанность представлять правдивую картину в рекламе, маркетинге и презентациях позволяет РИЭЛТОРАМ® использовать и демонстрировать только профессиональные обозначения, сертификаты и другие полномочия, они имеют законное право. (Принято 01.08)
Статья 13
РИЭЛТОРЫ® не должны заниматься деятельностью, которая представляет собой несанкционированную юридическую практику, и должны рекомендовать привлечение адвоката, когда этого требуют интересы любой стороны сделки.
Статья 14
В случае предъявления обвинений в неэтичных действиях или в случае просьбы представить доказательства или сотрудничать каким-либо иным образом в ходе любого судебного разбирательства или расследования, связанного с профессиональными стандартами, РИЭЛТОРЫ® должны передать все относящиеся к делу факты в соответствующие суды Совета членов или аффилированного института. , общество или совет, в котором состоят члены, и не должны предпринимать никаких действий, чтобы нарушить или воспрепятствовать таким процессам. (Изм. от 1/99)
Стандарт практики 14-1
РИЭЛТОРЫ® не должны подвергаться дисциплинарным разбирательствам более чем в одном Совете РИЭЛТОРОВ® или аффилированном институте, обществе или совете, в котором они являются членами в отношении к предполагаемым нарушениям Кодекса этики, относящимся к той же сделке или событию. ( с поправками 1/95)
Стандарт практики 14-2
РИЭЛТОРЫ® не должны несанкционированно раскрывать или распространять утверждения, выводы или решения, выработанные в связи со слушанием или апелляцией по вопросам этики или в связи с арбитражным разбирательством.
слушания или процессуального пересмотра. (Изм. от 1/92) Стандарт практики 14-3
РИЭЛТОРЫ® не должны препятствовать расследованию Совета или разбирательствам по профессиональным стандартам, возбуждая или угрожая возбуждением исков за клевету, клевету или диффамацию против любой стороны профессионального разбирательства по стандартам или их свидетелей на основании подачи арбитражного запроса, жалобы на этические нормы или свидетельских показаний, данных перед любым трибуналом. (Принято 1 ноября 1987 г., изменено 1 ноября 1999 г.)
Стандарт практики 14-4
РИЭЛТОРЫ® не должны намеренно препятствовать следственным или дисциплинарным разбирательствам Совета путем подачи нескольких жалоб на нарушение этических норм, основанных на одном и том же событии или сделке. (Принято 11/88)
Обязанности РИЭЛТОРОВ®
Статья 15
РИЭЛТОРЫ® не должны сознательно или по неосторожности делать ложные или вводящие в заблуждение заявления о других специалистах по недвижимости, их бизнесе или их деловой практике.
(Поправка от 1/12) Стандарт практики 15-1
РИЭЛТОРЫ® не должны намеренно или по неосторожности подавать ложные или необоснованные жалобы на этические нормы. (Принято 1/00)
Стандарт практики 15-2
Обязательство воздерживаться от ложных или вводящих в заблуждение заявлений о других специалистах по недвижимости, их бизнесе и их деловой практике включает в себя обязанность не публиковать информацию сознательно или по неосторожности , повторять, ретранслировать или переиздавать ложные или вводящие в заблуждение заявления, сделанные другими. Эта обязанность применяется независимо от того, повторяются ли ложные или вводящие в заблуждение заявления лично, в письменной форме, с помощью технических средств (например, Интернета) или любых других средств. (Принято 07.01, изменено 12.01)
Стандарт практики 15-3
Обязательство воздерживаться от ложных или вводящих в заблуждение заявлений о других специалистах по недвижимости, их бизнесе и их деловой практике включает в себя обязанность публиковать разъяснения или удалять заявления, сделанные другими на электронных носителях, контролируемых РИЭЛТОРОМ®, как только РИЭЛТОР® узнает, что заявление является ложным или вводящим в заблуждение.
(Принято 10 января, изменено 12 января) Статья 16
РИЭЛТОРЫ® не должны заниматься какой-либо практикой или предпринимать какие-либо действия, несовместимые с соглашениями об эксклюзивном представительстве или эксклюзивных брокерских отношениях, которые другие РИЭЛТОРЫ® имеют с клиентами. (Изм. 1/04)
Стандарт практики 16-1
Статья 16 не предназначена для запрета агрессивной или новаторской деловой практики, которая во всем остальном является этичной, и не запрещает разногласий с другими РИЭЛТОРАМИ®, касающихся комиссионных, гонораров, компенсаций или другие формы оплаты или расходов. (Принято 1/93, изменено 1/95)
Стандарт практики 16-2
Статья 16 не запрещает РИЭЛТОРАМ делать общие объявления потенциальным клиентам с описанием их услуг и условий их доступности, даже если некоторые получатели могут заключили агентские соглашения или другие эксклюзивные отношения с другим РИЭЛТОРОМ®. Общий телефонный опрос, общая почтовая рассылка или рассылка, адресованные всем потенциальным клиентам в данной географической области или в данной профессии, бизнесе, клубе или организации, или другой классификации или группе, считаются «общими» для целей настоящего стандарта.
(Изм. 01/04) Статья 16 призвана признать неэтичными два основных типа домогательств:
Во-первых, телефонные или личные домогательства владельцев собственности, которые были идентифицированы знаком недвижимости, составлением нескольких списков или другая информационная служба, поскольку они исключительно перечислили свою собственность у другого РИЭЛТОРА® и
Во-вторых, почта или другие формы письменных предложений потенциальных клиентов, чьи свойства указаны исключительно у другого РИЭЛТОРА®, когда такие предложения не являются частью общей рассылки, а направлены конкретно владельцам недвижимости, идентифицированным посредством компиляции текущих списков, вывесок «для продажи» или «сдается в аренду», или других источников информации, требуемых Статьей 3 и правилами службы множественных списков, которые должны быть доступны другим РИЭЛТОРАМ® в соответствии с предложениями субагентства или сотрудничества. (Изм. 04.01)
Стандарт практики 16-3
Статья 16 не запрещает РИЭЛТОРАМ® связываться с клиентом другого брокера с целью предложения или заключения договора на предоставление другого тип услуги в сфере недвижимости, не связанный с типом услуги, предоставляемой в настоящее время (например, управление недвижимостью в отличие от брокерских услуг), или от предложения того же типа услуги в отношении недвижимости, на которую не распространяются эксклюзивные соглашения с другими брокерами.
Однако информация, полученная через Службу множественного листинга или любое другое предложение о сотрудничестве, не может быть использована для целевых клиентов других РИЭЛТОРОВ®, которым могут быть сделаны такие предложения об оказании услуг. (Изм. 1/04) Стандарт практики 16-4
РИЭЛТОРЫ® не должны запрашивать листинг, который в настоящее время котируется исключительно у другого брокера. Однако, если листинговый брокер по запросу РИЭЛТОРА® отказывается раскрыть дату истечения срока действия и характер такого листинга, то есть исключительное право на продажу, эксклюзивное агентство, открытый листинг или другую форму договорного соглашения между листингом. брокера и клиента, РИЭЛТОР® может связаться с владельцем, чтобы получить такую информацию, и может обсудить условия, на которых РИЭЛТОР® может принять участие в листинге в будущем или, в качестве альтернативы, может принять листинг, который вступит в силу после истечения срока действия любого существующего эксклюзивного листинга.
(с поправками от 1/94) Стандарт практики 16-5
РИЭЛТОРЫ® не должны запрашивать соглашения покупателя/арендатора от покупателей/арендаторов, на которых распространяются эксклюзивные соглашения покупателя/арендатора. Однако, если РИЭЛТОР® попросит брокера отказаться раскрыть дату истечения срока действия эксклюзивного соглашения покупателя/арендатора, РИЭЛТОР® может связаться с покупателем/арендодателем, чтобы получить такую информацию, и может обсудить условия, на которых РИЭЛТОР® может заключить договор. в будущее соглашение покупателя/арендатора или, в качестве альтернативы, может заключить соглашение покупателя/арендатора, вступающее в силу после истечения срока действия любого существующего эксклюзивного соглашения покупателя/арендатора. (Принято 1/94, изменено 1/98)
Стандарт практики 16-6
Когда клиент другого РИЭЛТОРА® связывается с РИЭЛТОРАМИ® по поводу создания эксклюзивных отношений для предоставления того же типа услуг, и РИЭЛТОРЫ® прямо или косвенно не инициировали такие обсуждения, они могут обсудить условия, на которых они могут заключить будущее соглашение, или, в качестве альтернативы, могут заключить соглашение, которое вступает в силу по истечении срока действия любого существующего эксклюзивного соглашения.
(Изм. 1/98) Стандарт практики 16-7
Тот факт, что потенциальный покупатель нанял РИЭЛТОРА® в качестве эксклюзивного представителя или эксклюзивного брокера в одной или нескольких прошлых сделках, не препятствует другим РИЭЛТОРАМ® искать таких будущий бизнес перспективы. (Изм. 1/04)
Стандарт практики 16-8
Тот факт, что эксклюзивное соглашение было заключено с РИЭЛТОРОМ®, не исключает и не запрещает любому другому РИЭЛТОРУ® заключать аналогичное соглашение после истечения срока его действия. предшествующего соглашения. (Изм. 1/98)
Стандарт практики 16-9
РИЭЛТОРЫ®, до заключения договора о представительстве, имеют прямое обязательство предпринять разумные договор на оказание подобных услуг в сфере недвижимости. (Изм. 04.01.)
Стандарт практики 16-10
РИЭЛТОРЫ®, выступающие в качестве представителей или брокеров покупателя или арендатора, должны сообщить об этих отношениях представителю или брокеру продавца/арендодателя при первом контакте и должны предоставить письменное подтверждение об этом раскрытии продавцу/представителю арендодателя или брокеру не позднее заключения договора купли-продажи или аренды.
(Изм. 04.01.) Стандарт практики 16-11
В отношении незарегистрированной недвижимости РИЭЛТОРЫ®, выступающие в качестве представителей или брокеров покупателя/арендатора, должны раскрыть эти отношения продавцу/арендодателю при первом контакте с этим покупателем/арендодателем и должен предоставить письменное подтверждение такого раскрытия информации продавцу/арендодателю не позднее заключения любого договора купли-продажи или аренды. (Изм. 1/04)
РИЭЛТОРЫ® должны запрашивать ожидаемую компенсацию у продавца/арендодателя при первом контакте. (Изм. 1/98)
Стандарт практики 16-12
РИЭЛТОРЫ®, действующие в качестве представителей или брокеров продавцов/арендодателей или субагентов листинговых брокеров, должны раскрывать информацию об этих отношениях покупателям/арендаторам в кратчайшие сроки и должны предоставить письменное подтверждение такого раскрытия информации покупателям/арендаторам не позднее заключения любого договора купли-продажи или аренды.
(Изм. 04.01.) Стандарт практики 16-13
Все сделки с недвижимостью, выставленной исключительно на продажу, или с покупателем/арендаторами, на которых распространяется эксклюзивное соглашение, должны осуществляться с представителем или брокером клиента, а не с клиентом, кроме как с согласия представителя клиента или брокера, или за исключением случаев, когда такие сделки инициированы клиентом.
Прежде чем предоставлять существенные услуги (такие как написание предложения о покупке или представление CMA) потенциальным клиентам, РИЭЛТОРЫ® должны узнать у потенциальных клиентов, являются ли они стороной какого-либо соглашения об исключительном представительстве. РИЭЛТОРЫ® не должны сознательно предоставлять существенные услуги, связанные с предполагаемой сделкой, потенциальным клиентам, которые являются сторонами соглашений об исключительном представительстве, кроме как с согласия эксклюзивных представителей потенциальных клиентов или по указанию потенциальных клиентов.
(Принято 1/93, изменено 1/04) Стандарт практики 16-14
РИЭЛТОРЫ® могут свободно вступать в договорные отношения или вести переговоры с продавцами/арендодателями, покупателями/арендаторами или другими лицами, которые не являются предметом эксклюзивного соглашения, но должны сознательно не обязывать их платить более одной комиссии, кроме как с их осознанного согласия. (Изм. 1/98)
Стандарт практики 16-15
В совместных сделках РИЭЛТОРЫ® должны компенсировать сотрудничающим РИЭЛТОРАМ® (главным брокерам) и не должны компенсировать или предлагать компенсацию, прямо или косвенно, любому из лицензиатов продаж наняты или связаны с другими РИЭЛТОРАМИ® без предварительного явного уведомления и согласия сотрудничающего брокера.
Standard of Practice 16-16
РИЭЛТОРЫ®, действующие в качестве субагентов или представителей покупателя/арендатора или брокеров, не должны использовать условия предложения о покупке/аренде, чтобы попытаться изменить предложение брокера по листингу о компенсации субагентам или покупателю.
/ представители арендатора или брокеры, а также не ставить представление оформленного предложения о покупке / аренде в зависимость от согласия брокера по листингу изменить предложение о компенсации. (с изменениями от 01.04.) Стандарт практики 16-17
РИЭЛТОРЫ®, действующие в качестве субагентов, представителей покупателя/арендатора или брокеров, не должны пытаться распространять предложение брокера по листингу о сотрудничестве и/или компенсацию другим брокерам без согласия брокера по листингу. (Изм. 1/04)
Стандарт практики 16-18
РИЭЛТОРЫ® не должны использовать информацию, полученную от листинговых брокеров через предложения о сотрудничестве, сделанные через несколько листинговых служб, или через другие предложения о сотрудничестве, чтобы направлять клиентов листинговых брокеров к других брокеров или для создания отношений покупателя/арендатора с клиентами листинговых брокеров, если такое использование не разрешено листинговыми брокерами.
(Изм. 1/02) Стандарт практики 16-19
Знаки, уведомляющие о продаже, аренде, аренде или обмене недвижимости, не должны размещаться на собственности без согласия продавца/арендодателя. (Поправка от 1/93)
Стандарт практики 16-20
РИЭЛТОРЫ®, до или после прекращения их отношений с их текущей фирмой, не должны побуждать клиентов своей нынешней фирмы к расторжению эксклюзивных договорных соглашений между клиентом и та фирма. Это не мешает РИЭЛТОРАМ® (принципалам) заключать соглашения со своими ассоциированными лицензиатами, регулирующие переуступку эксклюзивных соглашений. (Принято 1/98, изменено 1/10)
Статья 17
В случае договорных споров или особых внедоговорных споров, как определено в Стандарте практики 17-4, между РИЭЛТОРАМИ® (принципалами), связанными с разными фирмами , вытекающие из их отношений в качестве РИЭЛТОРОВ®, РИЭЛТОРЫ® должны урегулировать спор, если Правление требует, чтобы его члены выступили посредником.
Если спор не разрешается посредством посредничества или если посредничество не требуется, РИЭЛТОРЫ® должны передать спор в арбитраж в соответствии с политикой Совета, а не рассматривать этот вопрос в судебном порядке.В случае, если клиенты РИЭЛТОРОВ® желают выступить посредником или арбитром в договорных спорах, возникающих в связи со сделками с недвижимостью, РИЭЛТОРЫ® должны выступить посредником или арбитром в этих спорах в соответствии с политикой Совета, при условии, что клиенты соглашаются быть связанными любым вытекающим из них соглашение или награда.
Обязательство участвовать в посредничестве и арбитраже, предусмотренное настоящей статьей, включает в себя обязательство РИЭЛТОРОВ® (доверителей) требовать от своих фирм проведения посредничества и арбитража и быть связанными любым вытекающим из них соглашением или решением. (Изм. 1/12)
Стандарт практики 17-1
Подача иска и отказ от него РИЭЛТОРАМИ® в арбитражном вопросе представляет собой отказ в арбитраже.
(Принято 02/86) Стандарт практики 17-2
Статья 17 не требует от РИЭЛТОРОВ® посредничества в тех обстоятельствах, когда все стороны в споре уведомляют Правление в письменной форме о том, что они решили не прибегать к посредничеству через удобства. Тот факт, что все стороны отказываются от участия в посредничестве, не освобождает РИЭЛТОРОВ® от обязанности арбитража.
Статья 17 не требует, чтобы РИЭЛТОРЫ® обращались в арбитраж в тех случаях, когда все стороны в споре уведомляют Совет в письменной форме о том, что они предпочитают не обращаться в арбитраж перед Советом. (Изм. 1/12)
Стандарт практики 17-3
РИЭЛТОРЫ®, действуя исключительно в качестве принципалов в сделке с недвижимостью, не обязаны разрешать споры в арбитраже с другими РИЭЛТОРАМИ® при отсутствии специального письменного соглашения об обратном . (Принято 1/96)
Standard of Practice 17-4
Конкретные внедоговорные споры, подлежащие арбитражному разбирательству в соответствии со статьей 17, включают:
продажа или аренда.
В таких случаях истец может указать первого сотрудничающего брокера в качестве ответчика, и арбитраж может продолжаться без указания листингового брокера в качестве ответчика. Когда арбитраж происходит между двумя (или более) сотрудничающими брокерами и когда листинговый брокер не является стороной, сумма спора и сумма любого возможного вытекающего из него решения ограничивается суммой, уплаченной ответчику листинговым брокером, и любой суммой, зачисленной либо выплачивается стороне по сделке по указанию ответчика. В качестве альтернативы, если жалоба подана против листингового брокера, листинговый брокер может указать первого сотрудничающего брокера в качестве стороннего ответчика. В любом случае решение комиссии по слушанию относительно причины закупки является окончательным в отношении всех текущих или последующих требований сторон о компенсации, вытекающих из лежащей в основе совместной сделки. (Принято 01/97, Изменено 01/07) 2) Если представитель покупателя или арендатора получает компенсацию от продавца или арендодателя, а не от листингового брокера, и листинговый брокер, в результате, уменьшает комиссию задолженности продавца или арендодателя, и после таких действий другой сотрудничающий брокер заявляет, что он является причиной продажи или аренды.
В таких случаях истец может указать первого сотрудничающего брокера в качестве ответчика, и арбитраж может продолжаться без указания листингового брокера в качестве ответчика. Когда арбитраж происходит между двумя (или более) сотрудничающими брокерами и когда листинговый брокер не является стороной, сумма спора и сумма любого возможного итогового решения ограничивается суммой, уплаченной ответчику продавцом или арендодателем, и любой суммой. зачисляется или выплачивается стороне по сделке по указанию ответчика. В качестве альтернативы, если жалоба подана против листингового брокера, листинговый брокер может указать первого сотрудничающего брокера в качестве стороннего ответчика. В любом случае решение комиссии по слушанию относительно причины закупки является окончательным в отношении всех текущих или последующих требований сторон о компенсации, вытекающих из лежащей в основе совместной сделки. (Принято 01/97, изменено 01/07) 3) Когда представитель покупателя или арендатора получает компенсацию от покупателя или арендатора, и в результате брокер по листингу уменьшает комиссию, причитающуюся с продавца или арендодателя, и, после таких действий другой сотрудничающий брокер заявляет, что он является причиной продажи или аренды.
В таких случаях истец может указать первого сотрудничающего брокера в качестве ответчика, и арбитраж может продолжаться без указания листингового брокера в качестве ответчика. В качестве альтернативы, если жалоба подана против листингового брокера, листинговый брокер может указать первого сотрудничающего брокера в качестве стороннего ответчика. В любом случае решение комиссии, проводящей слушания, в отношении причины закупки является окончательным в отношении всех текущих или последующих требований сторон о компенсации, вытекающих из лежащей в основе совместной сделки. (Принято 01/97) 4) Когда два или более листинговых брокера заявляют о праве на компенсацию в соответствии с открытыми листингами с продавцом или арендодателем, который соглашается участвовать в арбитраже (или который ходатайствует об арбитраже) и который соглашается быть связанным решение. В тех случаях, когда один из брокеров по листингу получил компенсацию от продавца или арендодателя, другой брокер по листингу, как истец, может указать первого брокера по листингу в качестве ответчика, и между брокерами может начаться арбитраж.
(Принято 01/97) 5) Когда представитель покупателя или арендатора получает компенсацию от продавца или арендодателя, а не от брокера по листингу, и брокер по листингу, в результате, уменьшает комиссию, причитающуюся с продавца, или арендодателем и после таких действий заявляет, что является причиной продажи или аренды. В таких случаях арбитраж должен быть между брокером по листингу и представителем покупателя или арендатора, и сумма спора ограничивается суммой уменьшения комиссии, на которую согласился брокер по листингу. (Принят 01.05)
Стандарт практики 17-5
Обязанность арбитража, установленная статьей 17, распространяется на споры между РИЭЛТОРАМИ® (доверителями) в различных государствах в случаях, когда в отсутствие установленного межассоциативного арбитражного соглашения, РИЭЛТОР® (принципал), ходатайствующий об арбитраже, соглашается подчиняться юрисдикции, путешествовать, участвовать и быть связанным любым результирующим решением, вынесенным в арбитраже, проводимом ассоциацией ответчика(ов) РИЭЛТОРА®, в случаях, когда ответчик (s) Ассоциация REALTOR® определяет, что существует арбитражный вопрос.
(Принято 07.01.) Пояснительные записки
Читатель должен знать о следующих правилах, утвержденных Советом директоров Национальной ассоциации:
При предъявлении обвинения в предполагаемом нарушении Кодекса Этики со стороны РИЭЛТОРА®, обвинение должно рассматриваться как предполагаемое нарушение одной или нескольких статей Кодекса. Стандарты практики могут быть процитированы в поддержку обвинения.
Стандарты практики служат для разъяснения этических обязательств, налагаемых различными статьями, и дополняют, но не заменяют толкование случаев в Интерпретации Кодекса этики .
Время от времени утверждаются модификации существующих стандартов практики и дополнительные новые стандарты практики. Читателей предупреждают, что они должны использовать самые последние публикации.
СТАТЬЯ II. ЗАКОНОДАТЕЛЬНАЯ ОРГАНА
Конституция Делавэра
§ 1. Генеральная ассамблея обладает законодательной властью; сочинение.
Раздел 1. Законодательная власть в этом штате принадлежит Генеральной Ассамблее, которая состоит из Сената и Палаты представителей.
§ 2. Состав Палаты и Сената; срок полномочий; районы; выборы.
Раздел 2. Палата представителей состоит из тридцати пяти членов плюс такие дополнительные члены, которые должны быть предоставлены в соответствии с разделом 2А настоящей статьи, которые избираются на два года. Сенат состоит из двадцати одного члена, избираемых на четыре года.
Настоящим штат разделен на тридцать пять представительных округов. Должны быть такие дополнительные представительные округа, которые предусмотрены в соответствии с разделом 2А настоящей статьи. От каждого представительного округа квалифицированными избирателями избирается по одному представителю. Штат также настоящим разделяется на двадцать один сенаторский округ, от каждого из которых квалифицированными выборщиками избирается по одному сенатору. В округе Нью-Касл должно быть семь сенаторских округов, пронумерованных от одного до семи включительно; в графстве Кент семь сенаторских округов, пронумерованных от одного до семи включительно; а в графстве Сассекс – семь сенаторских округов от первого до семи включительно.
Представительными округами округа Нью-Касл являются и будут следующими:
Номер один. Вся эта часть города Уилмингтон, входящая в состав Второго и Четвертого округов, а также те части Шестого и Восьмого округов, соответственно, лежащие к югу от центральной линии Восьмой улицы и ограниченные ею.
Номер два. Вся та часть указанного города, входящая в Девятый район, и те части Шестого и Восьмого районов, соответственно, лежащие к северу от центральной линии Восьмой улицы и ограниченные ею.
Номер три. Вся эта часть указанного города, входящая в Седьмой район, и та часть Пятого района, лежащая к северу от прямой линии, включающей центральную линию Восьмой улицы, и ограниченная ею.
Номер четыре. Вся часть указанного города, входящая в состав Первого и Третьего районов, и та часть Пятого района, лежащая к югу от центральной линии Восьмой улицы и ограниченная ею, к востоку от центральной линии улицы Адамс и к западу от нее. и ограничен центральной линией Рыночной улицы.
Номер пять. Вся та часть указанного города, которая входит в Десятый, Одиннадцатый и Двенадцатый районы, а также та часть Пятого района, лежащая к югу от и ограниченная прямой линией, включая центральную линию Восьмой улицы, к западу от центральной линии и ограниченная ею. Адамса, и ограниченный на западе западной границей указанного города.
Номер шесть. Брендивайн Сотня.
Номер семь. Кристиана Сотня.
Номер восемь. Милл Крик Сотня.
Номер девять. Уайт Клэй Крик Сотня.
Номер десять. Новая замковая сотня.
Номер Одиннадцать. Пенкадер Сотня.
Номер двенадцать. Красная львиная сотня.
Номер тринадцать. Георгиевская сотня.
Номер четырнадцать. Аппоквиниминк Сотня.
Номер пятнадцать. Сотня черных дроздов.
Представительными округами в графстве Кент являются и будут следующие:
Номер Один. Дак Крик Сотня.
Номер два. Литтл-Крик-Сотня и первый избирательный округ Ист-Дувр-Сотни.
Номер три.
Кентон Сотня.Номер четыре. Западная Дуврская Сотня и вся та часть Восточной Дуврской Сотни, лежащая рядом с Западной Дуврской Сотней и отделенная от остальной части Восточной Дуврской Сотни следующими граничными линиями: начало в середине дороги общего пользования, ведущей от дороги Хорсхед к Кентону в точке пересечения улиц Кентон-Сотня и Восточная Дуврская Сотня, оттуда проходит по середине указанной дороги до дороги Конской Головы, оттуда проходит в западном направлении по середине указанной дороги Конской Головы на небольшом расстоянии до короткой дороги, ведущей от указанной дороги Конской Головы дорога к дороге из Дувра в Хэзлетвилль, известной как дорога Хэзлетвилля, затем проходит по середине указанной короткой дороги от дороги Хорсхед к указанной дороге Хэзлетвилля, а оттуда проходит в западном направлении по середине указанной дороги Хэзлетвилля. недалеко от дороги, ведущей оттуда в Вайоминг, а оттуда проходит по середине указанной дороги, ведущей от указанной дороги Хазлетвилл в Вайоминг, до точки пересечение Восточной Дуврской Сотни и Северной Сотни Мердеркилл.
Номер пять. Вся та часть Ист-Дуврской сотни, не включенная в округа номер два и четыре.
Номер шесть. Части Северной Сотни Мёрдеркилл, Южной Сотни Мёрдеркилл и Миспиллион Сотни включены в следующие границы: начиная с пересечения южной линии Южной Сотни Мёрдеркилл со штатом Мэриленд, затем вдоль линии раздела между Сотней Миспиллион и Южной Сотни Мёрдеркилл до дорога общего пользования, ведущая из Уайтлисбурга в Харрингтон, оттуда идущая в юго-восточном и восточном направлении вдоль середины указанной дороги общего пользования к дороге общего пользования, ведущая от Корнера Мастена в Вернон, в Уайтс-Черч или рядом с ней, а затем идущая в северо-восточном направлении по середине указанной дороги общего пользования. указанная дорога общего пользования, ведущая из Мастенс-Корнер в Вернон, недалеко от дороги общего пользования, ведущей оттуда в город Харрингтон, является продолжением дороги, ведущей из Уайтлисбурга в Харрингтон, а оттуда идет в юго-восточном направлении до пересечения с Вест-стрит в город Харрингтон, оттуда идущий в северном направлении по середине Уэст-стрит до середины Уолкотт-стрит в указанном городе Харрингтон, оттуда в восточном направлении вдоль середины указанной Уолкотт-стрит до середины Дорман-стрит в указанном городе Харрингтон, оттуда в северном направлении вдоль середины от Дорман-стрит до Браунс-Бранч, оттуда она идет в восточном направлении по ходу указанной ветки до Делавэрской железной дороги, оттуда идет в северном направлении вдоль указанной Делавэрской железной дороги до Бивер-Дам-Бранч в Саут-Мердеркилл-Сотни, оттуда следует по курсу указанной Бивер-Бранч Ответвление плотины в северо-западном направлении к дороге общего пользования, ведущей из Фелтона в Уайтлисбург, оттуда идет в северо-восточном направлении вдоль середины указанной дороги общего пользования из Фелтона в Уайтлисбург к дороге Совиного гнезда, оттуда проходит в северном направлении по середине упомянутая дорога Совиного Гнезда до пересечения Каугилл-роуд из Вудсайда в Петербург, оттуда идущая в северном направлении n вдоль середины упомянутой Каугилл-роуд до Рид-роуд, идущей от Вудсайда к школьному дому Дюпона, оттуда она идет в северо-западном направлении по середине упомянутой Рид-роуд к школьному дому Дюпона, а оттуда идет в северном направлении по середине улицы Дюпон.
дорога общего пользования, ведущая из Уиллоу-Гроув в Кэмден, недалеко от Стаббс-Корнер, оттуда проходит в западном, северо-западном и западном направлениях вдоль середины дороги общего пользования, ведущей от школы Дюпона к богадельне к Грейс-Корнер, а оттуда продолжается в виде прямая западная линия до южной пограничной линии Западной Дуврской Сотни, оттуда следует по южной пограничной линии Западной Дуврской Сотни в западном направлении до штата Мэриленд, оттуда проходит в южном направлении вдоль восточной пограничной линии до штата Мэриленд до место начала.Номер семь. Вся та часть Северной Сотни Убийц Убийц, не входящая в округ номер шесть.
Номер восемь. Вся та часть Южной Сотни Убийц Убийц, не входящая в округ номер шесть.
Номер девять. Вся та часть Миспиллион Сотни, не вошедшая в округ номер шесть.
Номер десять. Милфордская сотня.
Представительные округа в округе Сассекс являются и будут следующими:
Номер один. Сидар-Крик Сотня.
Номер два. Вся та часть Нантикок-Сотни, которая лежит к северу и западу от Грэвелли-Бранч, начиная с точки, где упомянутая Грэвелли-Бранч пересекает разделительную линию между Джорджтауном и Нантикок-Сотнями, и тянется на юго-запад к тому, что раньше было известно как Старая мельница Реста, отсюда вдоль указанного ответвления к тому, что раньше называлось Коллинз-Миллс, до его устья, находящегося у истока пруда Миддлфорд-Милл; вместе с North West Fork Hundred.
Номер три. Вся та часть Нантикокской Сотни, которая лежит к югу и востоку от указанной Гравли-Бранч, начиная с точки, где указанная Гравийная ветвь пересекает разделительную линию между Нантикокской и Джорджтаунской сотнями, и проходит в юго-западном направлении к тому, что ранее было известно как Старая мельница Реста, оттуда вдоль указанного ответвления к месту, ранее известному как Коллинз-Миллс, до его устья у истока пруда Миддлфорд-Милл; вместе с Сифордской сотней.
Номер четыре. Брод-Крик Сотня.
Номер пять.
Литтл Крик Сотня.Номер шесть. Дагсборо и Гамборо Сотни.
Номер семь. Балтиморская сотня.
Номер восемь. Индийская речная сотня.
Номер девять. Джорджтаунская сотня.
Номер десять. Бродкилн, Льюис и Рехобот Сотни.
Сенаторские округа округа Нью-Касл являются и будут следующими:
Номер один. Вся эта часть города Уилмингтон, лежащая к северу от прямой линии и ограниченная ею, включая центральную линию Восьмой улицы, идущую от реки Делавэр до западной границы указанного города.
Номер два. Вся та часть указанного города, лежащая к югу от указанной выше прямой линии и ограниченная ею, включая центральную линию Восьмой улицы.
Номер три. Brandywine Hundred вместе со всей той частью Christiana Hundred, лежащей к северу от центральной линии Lancaster Turnpike и ограниченной ею.
Номер четыре. Милк-Крик-Сотня вместе со всей той частью Кристиана-Сотни, лежащей к югу от центральной линии Ланкастерской магистрали и ограниченной ею.
Номер пять. Сотня Уайт-Клэй-Крик, Сотня Красного Льва и Сотня Нью-Касл.
Номер шесть. Пенкадерская сотня и сотня Святого Георгия.
Номер семь. Аппоквиниминковая сотня и сотня дроздов.
Сенаторские округа округа Кент являются и будут следующими:
Номер Один. Первый и второй представительные округа.
Номер два. Третий и четвертый представительные округа.
Номер три. Пятый и седьмой представительные округа.
Номер четыре. Шестой и девятый представительные округа.
Номер пять. Восьмой и десятый представительные округа.
Номер Шесть, на свободе. Первый, второй, пятый, седьмой и восьмой представительные округа.
Номер семь, на свободе. Третий, четвертый, шестой, девятый и десятый представительные округа.
Сенаторские округа округа Сассекс являются и будут следующими:
Номер один. Первый и второй представительные округа.
Номер два. Третий и четвертый представительные округа.
Номер три.
Пятый и шестой представительные округа.Номер четыре. Седьмой и восьмой представительные округа.
Номер пять. Девятый и десятый представительные округа.
Номер Шесть, на свободе. Первый, второй, третий, четвертый и пятый представительные округа.
Номер семь, на свободе. Шестой, седьмой, восьмой, девятый и десятый представительные округа.
Первые сенаторы, избранные от шестых сенаторских округов графств Кент и Сассекс, избираются только на двухлетний срок, после чего их преемники избираются на полный четырехлетний срок.
Вся территория, которая в дальнейшем будет присоединена к городу Уилмингтон и включена в его состав, станет частью Представительных округов в округе Нью-Касл, как указано ниже:
Все, лежащие к востоку от прямой линии, включая центральную линию Маркет-стрит, Восьмая улица, поскольку в настоящее время существуют указанные две улицы, и южнее прямой линии, включая центральную линию Восьмой улицы, поскольку она существует в настоящее время, должны стать частью Представительного округа номер один.
Все расположено к северу от прямой линии, включая центральную линию Восьмой улицы, как она существует сейчас, простираясь от северо-восточной стороны Брендивайн-Крик до реки Делавэр или к северу от Брендивайн-Крик, к западу от точки пересечения указанная прямая линия с северо-восточной стороной указанного ручья станет частью Представительного округа номер два.
Все, что расположено к северу от прямой линии, включая центральную линию Восьмой улицы, как она существует сейчас, к югу от Брендивайн-Крик и к западу от центральной линии Маркет-стрит, как она существует сейчас, станет частью Представительства Район номер три.
Все, что находится между прямой линией, включающей центральную линию улицы Маркет, продолжающейся на юг, и прямой линией, включающей центральную линию улицы Вашингтон, продолжающейся на юг, должно стать частью Представительного округа номер четыре.
Все, что расположено к югу от прямой линии, включая центральную линию Восьмой улицы, как она существует в настоящее время, и к западу от прямой линии, включая центральную линию улицы Вашингтон, как она существует в настоящее время, должны стать частью номера представительного округа.
пять.В случае любого изменения линии границы между этим штатом и штатом Пенсильвания любой из указанных сенаторских и представительных округов в округе Нью-Касл, затронутый этим, должен соответствовать любой новой линии границы между указанными штатами.
Вся территория, которая в дальнейшем будет присоединена к городу Уилмингтон и включена в его состав, станет частью сенаторских округов округа Нью-Касл следующим образом: река Делавэр на запад, станет частью Сенаторского округа номер один.
Все, что расположено к югу от прямой линии, включая центральную линию Восьмой улицы, простирающейся от реки Делавэр на запад, должно стать частью Сенаторского округа номер два.
Всякий раз, когда путем расширения границ города Уилмингтон территория, входящая в состав любого представительного или сенаторского округа, как это установлено настоящим, будет включена в пределы указанного города, такой представительный или сенаторский округ впоследствии будет состоять из остатка из них, не включенные в указанные пределы.
Несколько представительных и сенаторских округов в штате, за исключением случаев, предусмотренных настоящим документом, продолжают ограничиваться, описываться и определяться линиями сотен, округов, избирательных округов, дорог общего пользования, железных дорог и других границ, упомянутых в настоящем документе, как такие же сейчас установлены и расположены.
§ 2А. Дополнительные представительные районы.
Раздел 2А. В дополнение к существующим 35 представительным округам, указанным в Разделе 2 настоящей статьи, должны быть дополнительные представительные округа, указанные ниже.
Каждый существующий представительный округ, указанный в Разделе 2 настоящей статьи, с населением, проживающим в нем более 15 000 человек, согласно последней официальной федеральной десятилетней переписи населения, имеет право на одного дополнительного представителя на каждые дополнительные 15 000 человек населения или большую часть из них, проживающих в округе.
Если какой-либо представительный округ, как указано в разделе 2 настоящей статьи, имеет право на более чем одного представителя, он должен быть подразделен на новые представительные округа для каждого дополнительного представителя, на которого он имеет право, из которых выбирается его квалифицированные выборщики, представитель.
После каждой официальной федеральной переписи населения, проводимой раз в десять лет, новые репрезентативные округа, созданные в соответствии с настоящим разделом, упраздняются, а репрезентативные округа, указанные в разделе 2 настоящей статьи, снова переделываются, как указано в настоящем документе.
Разделение представительных округов, как указано в настоящем документе, осуществляется Комиссией по изменению избирательного округа, состоящей из губернатора в качестве председателя и председателя штата от двух политических партий, получивших наибольшее количество голосов на пост губернатора на предыдущих выборах. Губернатор в качестве советников губернатора. Перераспределение округов и перераспределение Комиссией, как указано в настоящем документе, должно осуществляться в соответствии со следующими критериями: каждый новый представительный округ должен, насколько это возможно, формироваться из смежной территории; должны быть как можно ближе по численности населения к другим новым округам, создаваемым в пределах существующего представительного округа; должны быть ограничены древними границами, основными дорогами, ручьями или другими естественными границами; и не создаваться таким образом, чтобы оказывать неправомерное предпочтение какому-либо лицу или политической партии.
В течение 120 календарных дней после официального отчета Президенту Соединенных Штатов о каждой десятилетней переписи (или в течение 120 календарных дней после вступления в силу настоящей поправки) Губернатор от имени Комиссии должен подать заявление Государственному секретарю план перераспределения и перераспределения, как предусмотрено в настоящем документе. Сразу же после подачи заявления губернатор издает указ о перераспределении округов и перераспределении. Секретарь штата распорядится опубликовать такое прокламацию в двух газетах, распространяемых в штате в течение двух недель подряд, в течение 20 дней после выпуска прокламации. Постановление вступает в силу в течение 30 дней после его опубликования.
Любой квалифицированный избиратель может обратиться в Верховный суд с требованием обязать губернатора посредством мандамуса или иным образом выполнить обязанности по перераспределению избирательных округов и перераспределению или исправить любую ошибку в перераспределении избирательных округов и перераспределении.
Заявление о принуждении Губернатора к выполнению обязанностей по перераспределению округов и перераспределению должно быть подано в течение тридцати дней после истечения 120 дней, отведенных Комиссии на подачу своего плана, если такой план не подан вовремя. Заявление о принудительном исправлении любой ошибки в перераспределении округов и перераспределении должно быть подано в течение тридцати дней после провозглашения. Первоначальная юрисдикция в этих вопросах принадлежит Верховному суду. По апелляции дело должно быть рассмотрено Верховным судом в соответствии с законом и фактами.§ 2Б. Делегаты Конституционного собрания.
Раздел 2Б. Количество делегатов и метод избрания делегатов на Конституционный Конвент, как это предусмотрено в Разделе 2 Статьи 16, не должны зависеть от добавления Представителей или Представительных округов в соответствии с Разделом 2А настоящей Статьи. Представительные округа, которые избирают делегатов на Конституционное собрание, указаны в разделе 2 настоящей статьи.
§ 3. Квалификация членов.
Раздел 3. Сенатором не может быть лицо, не достигшее возраста двадцати семи лет и являвшееся гражданином и жителем штата в течение трех лет, предшествующих дню его или ее избрания, и в последний год этого назначать жителя сенаторского округа, в котором он или она должны быть избраны, за исключением случаев, когда он или она отсутствовал по государственным делам Соединенных Штатов или этого штата. Ни одно лицо не может быть Представителем, если оно не достигло возраста двадцати четырех лет и было гражданином и жителем штата в течение трех лет, предшествующих дню его или ее избрания, и в последний год этого срока являлось его жителем. представительного округа, в котором он или она должны быть избраны, за исключением случаев, когда он или она отсутствовал по государственным делам Соединенных Штатов или этого штата.
§ 4. Время и частота сеансов.
Раздел 4. Генеральная ассамблея созывается во второй вторник января каждого календарного года, если иное не созвано Губернатором или по взаимному созыву председателей обеих Палат.
Генеральная ассамблея может продолжать свои сессии каждый календарный год до тех пор, пока, по ее мнению, этого требуют общественные интересы; однако каждая сессия не должна продолжаться дольше последнего дня июня, если сессия не будет отозвана губернатором или по взаимному созыву председателей обеих палат.
§ 5. Место встречи.
Раздел 5. Генеральная ассамблея собирается и заседает в Дувре, столице штата; при условии, однако, что в периоды чрезвычайной ситуации, вызванной вражеским нападением, терроризмом, болезнью, несчастным случаем или другим стихийным или антропогенным бедствием, Генеральная Ассамблея может временно собираться и заседать в другом месте.
§ 6. Вакансии; срок пребывания в должности лиц, избранных для заполнения.
Раздел 6. Всякий раз, когда в какой-либо из Палат Генеральной Ассамблеи возникает вакансия по причине неизбрания, неправомочности, смерти, отставки или по иным причинам, председательствующий в Палате, в которой при наличии вакансии или в случае необходимости иным способом, предусмотренным законом; и лицо, выбранное впоследствии для заполнения такой вакансии, остается в должности до конца срока.
И всякий раз, когда в какой-либо из палат появляется такая вакансия, а Генеральная Ассамблея не заседает, губернатор имеет право издать указ о выборах для заполнения такой вакансии, который должен быть оформлен как приказ, изданный председательствующим должностным лицом любая из палат в случае вакансии, и лицо, выбранное в связи с этим для заполнения такой вакансии, остается в должности до конца срока.§ 7. Временный президент, спикер и другие должностные лица; отсутствие председательствующих.
Раздел 7. Сенат на первой ежегодной сессии каждой новой Генеральной Ассамблеи избирает одного из своих членов временным президентом, который председательствует в отсутствие вице-губернатора, или в случае, если последний становится губернатором, или пока он или она продолжает исполнять обязанности губернатора по причине инвалидности губернатора. Сенат также избирает других своих должностных лиц и в отсутствие лейтенант-губернатора и своего временного президента может время от времени, в зависимости от обстоятельств, назначать одного из своих членов председательствовать.
Палата представителей на такой первой ежегодной сессии избирает одного из своих членов спикером, а также выбирает других своих должностных лиц, а в отсутствие спикера может время от времени, в зависимости от обстоятельств, назначать одного из своих членов председательствовать.§ 8 Каждая палата как судья выборов и квалификации ее членов; кворум; отсрочки; убедительная посещаемость.
Раздел 8. Каждая Палата является судьей выборов, результатов и квалификации своих членов; и большинство всех членов, избранных в каждую Палату, составляют кворум для ведения дел; но меньшее число может откладывать заседания со дня на день и будет иметь право принудить к явке отсутствующих членов таким образом и под такими наказаниями, которые будут сочтены целесообразными.
§ 9. Правила; наказание и исключение членов; объем полномочий.
Раздел 9. Каждая палата может устанавливать правила своей работы, наказывать любого из своих членов за хулиганство и с согласия двух третей всех избранных в нее членов исключать члена, а также имеет все другие полномочия, необходимые для ветвь Законодательного собрания свободного и независимого государства.
§ 10. Журналы; публикация; ввод да и нет; принятие законопроектов и постановлений.
Раздел 10. Каждая палата ведет журнал своих заседаний и публикует его сразу же после каждой сессии, за исключением тех частей, которые могут потребовать соблюдения секретности. Имена членов, голосующих за и против любого законопроекта или совместного решения, за исключением случаев переноса заседания, при окончательном голосовании заносятся в журнал; и да и нет членов по любому вопросу, по желанию любого члена, должны быть внесены в журнал. Никакой законопроект или совместная резолюция, кроме как в отношении отсрочки, не могут быть приняты ни одной из палат, если окончательное голосование не было проведено по принципу «за» и «против» или без согласия большинства всех членов, избранных в каждую палату.
§ 11. Доступность каждой палаты и комитетов полного состава.
Раздел 11.
Двери каждой палаты и комитетов полного состава должны быть открыты, за исключением тех случаев, когда речь идет о деле, которое следует держать в секрете.§ 12. Согласие каждой палаты на отсрочку.
Раздел 12. Ни одна из палат не может без согласия другой переносить заседание более чем на три дня или в любое другое место, кроме того, в котором заседают две палаты.
§ 13. Иммунитет от ареста и допроса выступлений.
Раздел 13. Сенаторы и представители во всех случаях, за исключением государственной измены, тяжкого преступления или нарушения общественного порядка, пользуются привилегией от ареста во время их присутствия на сессии их соответствующих палат, а также при входе и возвращении из одной и той же палаты; и для любой речи или дебатов ни в одной из палат они не должны подвергаться сомнению в каком-либо другом месте.
§ 14.
Сотрудничество по совместительству или заинтересованность в контракте с армией или флотом.Раздел 14. Ни один сенатор или представитель в течение срока, на который он или она должны быть избраны, не может быть назначен на какую-либо гражданскую должность в этом штате, которая будет создана или вознаграждение которой будет увеличено в течение такого срока. время. Ни член Конгресса, ни любое лицо, занимающее какую-либо должность в этом штате или Соединенных Штатах, за исключением должностных лиц, обычно назначаемых судами соответственно, адвокатов и офицеров милиции, не занимающих дисквалификационных должностей, во время его или ее пребывание в Конгрессе или на посту сенатора или представителя; и ни одно лицо, заинтересованное в каком-либо контракте с армией или флотом, не может быть сенатором или представителем.
§ 15. Вознаграждение, расходы и пособия членов.
Раздел 15. Председатель Сената и члены Генеральной Ассамблеи получают годовой оклад и ежегодную компенсацию расходов на транспорт и другие необходимые и надлежащие цели, которые Генеральная Ассамблея устанавливает законом.
Средства, выделяемые по настоящему Договору, выплачиваются из Государственного казначейства.§ 16. Ограничение законопроектов и постановлений одной темой; выражение в названии; исключение.
Статья 16. Никакой законопроект или совместное решение, за исключением законопроектов об ассигновании денег на общественные нужды, не должны охватывать более одного предмета, что должно быть выражено в его заголовке.
§ 17. Лотереи и другие азартные игры.
Раздел 17. В этом штате запрещены все формы азартных игр, за исключением следующих:
(a) Лотереи под контролем штата с целью сбора средств,
(b) Лотереи (кроме игровых автоматов, рулетки, игры в кости и баккара) при условии, что каждая из них спонсируется и проводится в соответствии с ограничениями Раздела 17B компаниями, организациями или обществами, которые существуют не менее 2 лет; при условии, однако, что ни одно лицо, не достигшее 18-летнего возраста, не может участвовать ни в какой лотерее (где призом являются деньги), иначе разрешенной статьей
(c) Пари или пари с использованием пари-мутуэля автоматы или тотализаторы на скачках, проводимых на ипподромах в штате или за его пределами, при условии, что такие ставки или пари могут проводиться только:
(1) на территории любого ипподрома, имеющего лицензию в соответствии с законодательством штата на проведение скачек, или
(2) в ограде любого ипподрома, имеющего лицензию в соответствии с законодательством штата на получение и прием пари или пари на одновременную трансляцию скачек по электронному телевидению.
(d) Игры в бинго, проводимые в соответствии с ограничениями Раздела 17A.
Генеральная ассамблея должна обеспечить соблюдение настоящего Раздела посредством соответствующего законодательства.
§ 17А. игры в бинго; организации, уполномоченные на проведение; вынесение на референдум; районы; регулирование; штрафы.
Раздел 17А. Игра в Бинго считается законной, если она спонсируется и проводится Добровольными пожарными компаниями, организациями ветеранов, религиозными или благотворительными организациями или братскими обществами при условии, что чистые поступления или прибыль от проведения или управления такими играми в Бинго вышеупомянутыми компаниями, Организации или общества используются исключительно для продвижения или достижения целей таких компаний, организаций или обществ, и при условии, что деятельность вышеупомянутых компаний, организаций или обществ соответствует положениям раздела 170.
Налогового кодекса США и правил, обнародованных в соответствии с ним министром финансов США.1. Генеральная ассамблея должна предусмотреть в соответствии с законом представление для голосования правомочных избирателей нескольких округов штата или любого из них, упомянутых в подпункте 2 статьи 17А настоящей статьи, на всеобщих выборах, проводимых в 1958 г., вопрос о том, следует ли лицензировать или запрещать игру в «Бинго» в ее пределах; и в каждом округе, в котором большинство голосов против лицензии, ни одна организация, упомянутая в Разделе 17А, не может после этого спонсировать или разрешать игру «Бинго» в указанном округе до тех пор, пока при последующем представлении такого вопроса не будет получено большинство голосов. быть отлитым в указанном районе для получения лицензии. Всякий раз, когда большинство всех членов, избранных в каждую палату Генеральной Ассамблеи квалифицированными выборщиками в любом округе, указанном в подпункте 2 раздела 17А настоящей статьи, требует вынесения вопроса о лицензии или отсутствии лицензии на голосование квалифицированных избирателей в указанном округе, Генеральная ассамблея должна обеспечить представление такого вопроса правомочным избирателям в таком округе на следующих после этого всеобщих выборах.
2. В соответствии с положениями настоящей статьи графство Сассекс включает один округ, графство Кент включает один округ, город Уилмингтон, как его корпоративные границы в настоящее время являются или могут быть расширены в будущем, один округ и оставшуюся часть Округ Нью-Касл, один округ.
3. Генеральная ассамблея принимает необходимые законы для выполнения и обеспечения соблюдения положений настоящей статьи, принимает законы, регулирующие игру «Бинго» в соответствии с ограничениями, установленными настоящей статьей, и может налагать такие санкции, которые могут быть необходимы для обеспечения соблюдения тех же положений. .
§ 17Б. лотереи, не находящиеся под контролем государства; организации, уполномоченные на проведение; вынесение на референдум; районы; регулирование; штрафы.
Раздел 17В. Лотереи, не находящиеся под контролем государства, являются законными, если их спонсируют и проводят добровольные пожарные команды, организации ветеранов, религиозные или благотворительные организации или братские общества при условии, что указанные компания, организация или общество существуют не менее 2 лет и при условии, что чистая поступления или прибыль, возникающие в результате проведения или управления такими лотереями вышеупомянутыми компаниями, организациями или обществами, используются исключительно для продвижения или достижения целей таких компаний, организаций или обществ, и при условии, что вышеупомянутые компании, организации или общества действуют таким образом, чтобы соответствовать § 170 Налогового кодекса Соединенных Штатов и правилам, обнародованным в соответствии с ним министром финансов Соединенных Штатов.
1. Генеральная ассамблея должна предусмотреть в соответствии с законом представление для голосования правомочных избирателей нескольких округов штата или любого из них, упомянутых в пункте 2 настоящей статьи, на всеобщих выборах, состоявшихся в 1984 г., вопрос о том, следует ли лицензировать или запрещать проведение лотерей, не находящихся под контролем государства, в его пределах; и в каждом округе, в котором большинство голосов против лицензии, ни одна организация, упомянутая в этом разделе, не может после этого спонсировать или разрешать лотереи, не находящиеся под контролем штата, в пределах указанного округа до тех пор, пока при последующем представлении такого вопроса не будет получено большинство голосов. литье в указанном районе для получения лицензии. Всякий раз, когда большинство всех членов, избранных в каждую палату Генеральной Ассамблеи квалифицированными избирателями в любом округе, указанном в пункте 2 настоящей статьи, требует вынесения вопроса о лицензии или отсутствии лицензии на голосование квалифицированных избирателей в указанном округа, Генеральная Ассамблея должна обеспечить представление такого вопроса правомочным избирателям в таком округе на следующих после этого всеобщих выборах.
2. В соответствии с этой статьей графство Сассекс включает 1 округ, графство Кент включает 1 округ, город Уилмингтон, как его корпоративные границы в настоящее время являются или могут быть расширены в будущем, 1 округ и оставшуюся часть округа Нью-Касл. , 1 микрорайон.
3. Генеральная Ассамблея должна принять всеобъемлющее законодательство, предусматривающее лицензирование всех организаций, проводящих и регулирующих проведение лотерей в соответствии с настоящим разделом, и может предусмотреть такие санкции, которые могут быть необходимы для обеспечения соблюдения такого законодательства.
§ 18. Развод или алименты.
Раздел 18. Развод не разрешается, а алименты не допускаются, кроме как по решению суда, как это предусмотрено общим и единообразным законом.
§ 19. Местные или специальные законы, касающиеся заборов, домашнего скота, канав, школьных округов, а также дорог, шоссе, улиц и т.
д.Раздел 19. Генеральная Ассамблея не может принимать какие-либо местные или специальные законы, касающиеся заборов; бродячий скот; канавы; создание или изменение границ школьных округов; или планировка, открытие, изменение, техническое обслуживание или отпуск, полностью или частично любой дороги, шоссе, улицы, переулка или переулка; при условии, однако, что Генеральная ассамблея может двумя третями голосов всех членов, избранных в каждую палату, принимать законы, касающиеся планировки, открытия, изменения или содержания любой дороги или шоссе, образующих непрерывную дорогу или шоссе, протянувшееся по крайней мере через часть из трех округов штата.
Никакие дороги, шоссе или улицы, предназначенные для общественного пользования и содержащиеся за государственный счет, не должны строиться иначе как в соответствии со стандартами, принятыми агентством, ответственным за строительство, реконструкцию или содержание таких дорог, шоссе или улиц. Любая дорога или улица, построенная исключительно для частного пользования, может содержаться за государственный счет только после того, как она будет построена или реконструирована в соответствии с нормами, установленными органом, на который возложена обязанность по содержанию таких дорог или улиц.
§ 20. Раскрытие личной или частной заинтересованности законодателя в любой рассматриваемой мере.
Раздел 20. Любой член Генеральной ассамблеи, имеющий личный или частный интерес в какой-либо мере или законопроекте, находящемся на рассмотрении Генеральной ассамблеи, должен сообщить об этом факте Палате, членом которой он или она является, и не должен голосовать по этому поводу.
§ 21. Осуждение за преступление как запрет на занятие государственной должности.
Раздел 21. Ни одно лицо, которое должно быть осуждено за растрату государственных денег, взяточничество, лжесвидетельство или другое позорящее преступление, не имеет права занимать место в любой из Палат Генеральной Ассамблеи или быть способным занимать какую-либо трастовую, почетную должность. или прибыль в соответствии с этим государством.
§ 22. Подкуп должностных лиц исполнительной, судебной или законодательной власти.
Раздел 22. Каждое лицо, которое дает, предлагает или обещает, прямо или косвенно, любые деньги, свидетельства, привилегии, личную выгоду или ценные вещи любому исполнительному или судебному должностному лицу этого штата или любому члену любой Палаты Генеральная ассамблея с целью оказания на него или на нее влияния при выполнении любых его или ее официальных или общественных обязанностей считается виновным во взяточничестве и подлежит наказанию в порядке, предусмотренном законом.
§ 23. Статуты как публичные законы, если не заявлено иное.
Статья 23. Каждый закон является публичным законом, если иное не указано в самом законе.
§ 24.
Расчеты Государственного казначея; лишение права занимать законодательные должности до урегулирования.Статья 24. Государственный казначей ежегодно проводит расчеты с Генеральной Ассамблеей или ее совместным комитетом, который назначается на каждую 90-дневную сессию. Ни одно лицо, занимавшее должность государственного казначея, не имеет права занимать место ни в одной из палат Генеральной Ассамблеи до тех пор, пока он или она не произведет окончательный расчет по своим счетам в качестве казначея и не погасит причитающийся остаток, если таковой имеется. на нем.
§ 25. Законы, разрешающие постановления о зонировании и использовании земли.
Раздел 25. Генеральная ассамблея может принимать законы, в соответствии с которыми муниципалитеты и графство Сассекс, графство Кент и графство Ньюкасл могут принимать постановления о зонировании, законы или правила, ограничивающие и ограничивающие определенные районы и регулирующие в них здания и сооружения.
в соответствии с их конструкцией и характером и степенью их использования, а также использованием земли в таких районах для других целей, кроме сельскохозяйственных; и осуществление таких полномочий считается относящимся к полицейской власти государства.Центр данных по альтернативным видам топлива: Сравнение свойств топлива
Химическая структура [1] C 4 по C 12 и этанол ≤ до 10% С 8 по С 25 н/д Метиловые эфиры C 12 – C 22 жирных кислот CH 3 CH 2 OH CH 4 (большинство), C 2 H 6 и инертные газы CH 4 то же, что CNG с инертными газами C 3 H 8 (большинство) и C 4 H 10 (меньшинство) Н 2 CH 3 ОН Топливный материал (сырье) Сырая нефть Сырая нефть Природный газ, уголь, атомная энергетика, энергия ветра, гидроэнергия, солнечная энергия и небольшой процент геотермальной энергии и биомассы Жиры и масла из таких источников, как соевые бобы, отработанное кулинарное масло, животные жиры и рапс Кукуруза, зерно или сельскохозяйственные отходы (целлюлоза) Подземные запасы и возобновляемый биогаз Подземные резервы и возобновляемый биогаз Побочный продукт переработки нефти или природного газа Природный газ, метанол и электролиз воды Природный газ, уголь или древесная биомасса Бензиновый или дизельный эквивалент в галлонах (GGE или DGE) 1 галлон = 1,00 GGE
1 галлон = 0,88 DGE1 галлон = 1,12 GGE
1 галлон = 1,00 DGE1 кВтч = 0,030 GGE
1 кВтч = 0,027 DGEB100
1 галлон = 1,05 GGE
1 галлон = 0,93 DGEB20
1 галлон = 1,11 GGE
1 галлон = 0,99 DGE1 галлон = 0,67 GGE
1 галлон = 0,59 DGE1 фунт = 0,18 GGE
1 фунт = 0,16 DGE1 фунт = 0,19 GGE
1 фунт = 0,17 DGE1 галлон = 0,74 GGE
1 галлон = 0,66 DGE1 фунт = 0,45 GGE
1 фунт = 0,40 DGE1 кг = 1 GGE
1 кг = 0,9 DGE1 галлон = 0,50 GGE
1 галлон = 0,45 DGEСравнение энергии [2] 1 галлон бензина содержит 97–100 % энергии 1 ГГЭ. Стандартное топливо — 90 % бензина, 10 % этанола.1 галлон дизельного топлива содержит 113% энергии 1 ГГЭ из-за более высокой плотности энергии дизельного топлива. Типичная батарея размером с галлон бензина (0,134 фута 3 ), при использовании на транспорте может хранить 15,3% энергии в 1 ГГЭ. [6][7] 1 галлон B100 содержит 93% энергии 1 DGE, а 1 галлон B20 содержит 99% энергии 1 DGE из-за более низкой плотности энергии биодизеля. 1 галлон E85 содержит 73–83 % энергии 1 GGE. 1 галлон E100 содержит 67% энергии 1 GGE. Этанол смешивают с примесью для смешения оксигенатов (компонент бензина). [5] 5,66 фунта или 123,57 фута 3 CNG имеет ту же энергию, что и 1 GGE, а 6,37 фунта или 139,30 фута 3 CNG имеет ту же энергию, что и 1 DGE. [3][4](б)5,37 фунта СПГ имеют ту же энергию, что и 1 ГГЭ, а 6,06 фунта СПГ имеют ту же энергию, что и 1 ДГЭ. (а) 1 галлон пропана имеет 73% энергии в 1 ГГЭ из-за меньшей плотности энергии пропана. 2,2 фунта. (1 кг) H 2 имеет ту же энергию, что и 1 GGE. 1 галлон метанола содержит 50% энергии 1 ГГЭ. Содержание энергии (низшая теплотворная способность) 112 114–116 090 БТЕ/галлон (в) 128 488 БТЕ/гал (в) 3 414 БТЕ/кВтч B100
119 550 БТЕ/галB20
126 700 БТЕ/гал (c)76 330 БТЕ/гал для E100 (c) 20 160 БТЕ/фунт [3](б) 21 240 БТЕ/фунт (а) 84 250 БТЕ/гал (в) 51 585 БТЕ/фунт (c) 33,3 кВтч/кг
57 250 БТЕ/гал (в) Содержание энергии (высшая теплотворная способность) 120 388–124 340 БТЕ/гал (в) 138 490 БТЕ/гал (в) 3 414 БТЕ/кВтч 127 960 БТЕ/гал для B100 (c) 84 530 БТЕ/галлон для E100 (c) 22 453 БТЕ/фунт [1](c) 23 726 БТЕ/фунт (в) 91 420 БТЕ/гал (в) 61 013 БТЕ/фунт (в) 65 200 БТЕ/гал (в) Физическое состояние Жидкость Жидкость Электричество Жидкость Жидкость Сжатый газ (легче воздуха) Криогенная жидкость (легче воздуха в виде газа) Жидкость под давлением (тяжелее воздуха в виде газа) Сжатый газ (легче воздуха) или жидкость Жидкость Цетановое число Н/Д 40–55 (г) н/д 48–65 (г) 0–54 (д) н/д н/д н/д н/д н/д Октановое число насоса 84–93 (е) Н/Д Н/Д н/д 110 (и) 120+ (ч) 120+ (в) 105 (г) 130+ (г) 112 (и) Температура вспышки -45°F (к) 165°F (к) н/д от 212° до 338°F (г) 55°F (к) -300°F (к) -306°F (к) от -100° до -150°F (j) н/д 54°F (к) Температура самовоспламенения 495°F (к) ~600°F (к) Н/Д ~300°F (г) 793°F (к) 1004°F (к) 1004°F (k) от 850° до 950°F (j) от 1050° до 1080°F (j) 897°F (к) Проблемы с обслуживанием Смазывающая способность улучшена по сравнению с обычным дизельным топливом с низким содержанием серы. Дополнительную информацию о техническом обслуживании см. в Руководстве по обращению с биодизельным топливом и его использованию — пятое издание. (г)Могут потребоваться специальные смазочные материалы. Практика очень похожа, если не идентична, на операции с обычным топливом. Резервуары высокого давления требуют периодической проверки и сертификации. СПГ хранится в криогенных резервуарах с определенным временем выдержки до сброса давления. Автомобиль следует эксплуатировать по графику, чтобы поддерживать более низкое давление в баке. Когда водород используется в топливных элементах, обслуживание должно быть минимальным. Необходимо использовать специальные смазочные материалы Резервуары высокого давления требуют периодической проверки и сертификации.в соответствии с указаниями поставщика, а также запасные части, совместимые с M85. Может вызвать серьезное повреждение органов тела, если человек проглотит его, вдохнет или попадет на кожу. Влияние на энергетическую безопасность Изготовлено с использованием масла. На транспорт приходится примерно 30% общих потребностей США в энергии и 70% потребления нефти. (л) г. Изготовлено с использованием масла. На транспорт приходится примерно 30% общих потребностей США в энергии и 70% потребления нефти. (л) Электроэнергия производится внутри страны из самых разных источников, в том числе с помощью угольных электростанций и возобновляемых источников, что делает ее универсальным топливом. Биодизель отечественного производства, возобновляемый и снижает потребление нефти на 95% на протяжении всего жизненного цикла. (м) Этанол отечественного производства. E85 снижает потребление бензина в течение всего жизненного цикла на 70 %, а E10 снижает потребление бензина на 6,3 %. (н) CNG производится внутри страны из природного газа и возобновляемого биогаза. Соединенные Штаты обладают огромными запасами природного газа. СПГ производится внутри страны из природного газа и возобновляемого биогаза. Соединенные Штаты обладают огромными запасами природного газа. Приблизительно половина сжиженного нефтяного газа в США производится из нефти, но нефть не импортируется специально для производства сжиженного нефтяного газа. Водород производится внутри страны и может производиться из возобновляемых источников. Метанол производится внутри страны, иногда из возобновляемых ресурсов. Ядерные и цитоплазматические включения хантингтина отличаются биохимическим составом, интерактомными и ультраструктурными свойствами ген хантингтина (HTT)
1,2,3,4 . Расширенные повторы транслируются в длинный патогенный полиглутаминовый (polyQ) тракт (> 36 повторов), что делает белок Huntingtin (Htt) более склонным к агрегации 5 . Длина повтора CAG обратно пропорциональна возрасту начала заболевания, при этом ювенильная форма связана с длиной повтора polyQ 75 или более 6,7 . Накопление Htt-позитивных интранейрональных агрегатов и включений в коре и стриатуме посмертного мозга больных БХ и в животных и клеточных моделях БХ 2 привели к гипотезе о том, что агрегация Htt играет центральную роль в патогенезе БХ. Однако наше понимание ультраструктурного и биохимического состава включений Htt и молекулярных механизмов, управляющих их образованием, клиренсом и токсичностью, остается неполным. Устранение этого пробела в знаниях имеет решающее значение для понимания молекулярных и клеточных механизмов, лежащих в основе БХ, и для разработки эффективных методов лечения и стратегий модификации этого заболевания.Хотя в нескольких исследованиях изучались специфические клеточные механизмы, связанные с агрегацией Htt, в современных моделях часто отсутствует подробная характеристика включений как на биохимическом, так и на структурном уровне. Кроме того, в то время как в некоторых из предыдущих исследований изучалось влияние длины повтора полиQ на ультраструктурные свойства мутантного Htt, нет сообщений о роли первых 17 N-концевых аминокислот (Nt17) Htt, которые регулируют многие аспекты агрегационные и клеточные свойства белков Htt 8,9,10,11 – на организацию и ультраструктурные свойства цитоплазматических и ядерных включений Htt.
Наконец, подавляющее большинство предыдущих исследований использовали мутантные конструкции Htt, слитые либо с метками на основе пептидов, либо с крупными флуоресцентными белками, такими как GFP, которые могли вмешиваться в клеточные и агрегационные свойства Htt 12,13,14,15 .Здесь мы использовали клеточные и нейрональные модели образования цитоплазматических и ядерных включений Htt, чтобы понять, как модификации последовательности влияют на окончательные ультраструктурные и биохимические свойства цитоплазматических и ядерных включений Htt и их влияние на клеточные органеллы и функции. Клеточные модели, используемые в этом исследовании, основаны на сверхэкспрессии N-концевых фрагментов мутантного Htt, включающих область Exon 1 (Httex1), которая содержит экспансию polyQ. Было показано, что неполный сплайсинг HTT, приводящий к экспрессии белка Httex1, происходит в мозгу пациентов с БХ , 16, , а белок Httex1 был ранее описан как ключевой компонент внутриклеточных включений, обнаруженных в посмертном мозге HD , 16,17,18, .
Более того, экспрессии патогенного Httex1 (полиQ-тракт > 43Q) достаточно, чтобы индуцировать признаки БГ, включая образование агрегатов и токсичность у мышей 19,20,21 , дрозофил 22 , C. elegans 23 и клеток культурные модели 11,12,15,24,25,26,27,28,29,30,31 . Исследования in vitro и на клетках также показали, что Httex1 агрегирует в зависимости от длины и концентрации повторов polyQ 32,33,34 . Поэтому модели на основе мутантного Httex1 полезны для изучения патогенеза БХ, поскольку они воспроизводят различные аспекты агрегации Htt и сыграли важную роль в улучшении нашего понимания последовательности, молекулярных и структурных детерминант агрегации Htt и образования включений 10,26 ,27,28,29,35,36 .Мы применили комбинацию коррелятивной световой и электронной микроскопии (CLEM) и подходов, основанных на протеомике, для исследования структурных и биохимических свойств цитоплазматических и ядерных включений Httex1 в HEK 29.
3 клетки и первичные корковые нейроны. Мы также исследовали роль домена Nt17 и длины поли-Q-тракта в модулировании состава, структурных и токсических свойств мутантных включений Httex1. Наконец, учитывая, что большое количество опубликованных клеточных исследований механизмов агрегации Httex1 и образования включений основано на конструкциях, в которых Httex1 слит с GFP 9,11,12,37,38,39,40 , мы также впервые сравнили состав, ультраструктурные свойства и токсичность включений, образованных нативными (без тегов) и меченными GFP мутантными белками Httex1.Наши результаты показывают, что включения Htt состоят из сложной смеси агрегированного мутантного Httex1, различных клеточных белков и мембранных органелл, включая эндосомальную систему. Мы показываем, что функциональные и ультраструктурные свойства включений Httex1 по-разному изменяются модификациями последовательности и взаимодействиями с липидами и клеточными органеллами. Мы показываем, что цитоплазматические и ядерные включения Htt имеют разный состав и ультраструктурные свойства, что предполагает разные механизмы агрегации, образования включений и токсичности.
Эти наблюдения показали, что различия в клеточной среде и интерактоме могут влиять на механизмы агрегации, структурные и биохимические свойства включений и их относительный вклад в токсичность, вызванную мутантным Htt. Наконец, мы показываем, что включения, продуцируемые мутантным Httex1 72Q-GFP, демонстрируют поразительные различия с точки зрения организации, ультраструктурных свойств, белкового состава и их влияния на митохондриальные функции по сравнению с включениями, образованными мутантным Httex1 72Q без меток. В целом, наши результаты способствуют лучшему пониманию последовательности, молекулярных и клеточных детерминант агрегации мутантного Httex1, образования включений и токсичности. Они также подчеркивают критическую важность дальнейших исследований для изучения роли, которую липиды, органеллы и интерактом Htt играют в формировании и созревании патологии Htt, а также индуцированной Htt токсичности при БХ.Результаты
Цитоплазматические включения Httex1 демонстрируют отличительную морфологию ядра и оболочки и состоят из высокоорганизованных фибрилл, цитоплазматических белков и мембранных структур
, в котором было показано, что сверхэкспрессия мутантного Httex1 с повторами полиQ > 39 приводит к надежному и воспроизводимому образованию цитоплазматических включений Htt 15,29 .
В этой модели ядерные включения также наблюдаются в ~15% клеток, трансфицированных Httex1 72Q 41 , что дает возможность исследовать и сравнивать ультраструктурные особенности цитоплазматических и ядерных включений Htt в идентичных условиях. Эта модельная система широко используется для исследования молекулярных, клеточных и фармакологических модуляторов агрегации Htt и образования включений 11,12,15,24,25,26,27,28,29,30,31,42,43 , хотя очень часто используются конструкции Htt с дополнительными неестественными последовательностями, такими как теги Myc, FLAG и GFP. Предыдущие исследования показали, что наличие таких меток существенно изменяет агрегационные свойства N-концевых фрагментов мутантного Htt 9.1441 44,45,46 . Поэтому в этой работе мы решили сохранить нативную последовательность белка и исследовали агрегацию мутантного Httex1 и образование включений в отсутствие дополнительных последовательностей или меток. Безметочный Httex1 72Q был сверхэкспрессирован в клетках HEK 293 (клетки HEK), и были оценены морфология и структурные свойства включений Httex1. Несмотря на образование и обилие включений, образованных Httex1 72Q, мы не наблюдали явной токсичности в клетках НЕК. Инициация апоптотических событий была очевидна только через 96 ч, на что указывает активация каспазы 3 без потери целостности плазматической мембраны 41 .Затем мы оценили морфологию включений Httex1 72Q с помощью иммуноцитохимии (ICC) с использованием нескольких антител против разных эпитопов вдоль последовательности Httex1 (рис. 1a и S1). Интересно, что все антитела проявляли сильную иммунореактивность к периферии включений Httex1 72Q, и ни одно из этих антител не метило сердцевину этих включений (рис. 1b и S2a). Это наблюдение предполагает либо отсутствие Htt в центре включений, либо плохую доступность используемых антител к ядру включений, возможно, из-за высокой компактности агрегатов Htt в ядре по сравнению с периферией включений. Кроме того, мы наблюдали высокую колокализацию филаментозного актина (визуализируемую фаллоидином) с включениями Httex1 (рис.
Рис. 1: Конфокальная микроскопия и CLEM выявили кольцеобразную структуру включений Httex1 72Q в клетках НЕК. 1b, белые стрелки), что указывает на возможное участие белков цитоскелета в формировании включений Htt.a Картирование эпитопов антител Httex1, использованных в этом исследовании. b Конфокальная визуализация включений Httex1 72Q, образовавшихся через 48 ч после трансфекции в клетках НЕК. Все антитела Htt проявляли сильную иммунореактивность к периферии включений. Ядро контрастировали DAPI, а F-актин – фаллоидином. Белые стрелки указывают на колокализацию F-актина с кольцеобразной структурой включений Httex1. Масштабная линейка = 20 мкм и 10 мкм. c , d CLEM включений Httex1 72Q. c Конфокальная визуализация включений Httex1 72Q, образовавшихся через 48 ч после трансфекции в клетках HEK. Выбранный участок (белый квадрат) исследовали с помощью ЭМ ( d ). Ортогональная проекция (Orth.
Pr.), Масштабные линейки = 5 мкм. d Присутствие фрагментов мембран и везикул указано белыми стрелками. Бинарное изображение (вставка) показывает ультраструктуру ядра и оболочки включения Httex1 72Q. Масштабная линейка = 500 нм. e Репрезентативные ЭМ-изображения включения Httex1 72Q. Большее увеличение (белый квадрат) показано на правой панели. Штриховые линии ограничивают агрегат и ядро включения. Интернализованные мембранные структуры и митохондрии обозначены оранжевой и зеленой стрелками соответственно. Масштабная линейка = 1 мкм и 500 нм. f 3D-модель клеточного включения Httex1 72Q и окружающих органелл (вид сверху). Оболочка тела включения Httex1 (IB) представлена фиолетовым цветом, ядро — голубым. Мембраны ЭР показаны зеленым цветом, мембранные структуры внутри включения – белым, ядро - синим, митохондрии – желтым, а расположенные стопкой цистерны ЭР обозначены красной стрелкой. Масштабная линейка = 2 мкм. г Репрезентативные ЭМ-изображения включения Httex1 39Q, образовавшегося через 48 часов после трансфекции в клетках НЕК. Белый квадрат обозначает область, показанную на правой панели при большем увеличении. Пунктирные линии ограничивают включение. Масштабная линейка = 1 мкм и 500 нм. h 3D-модель Httex1 39Q IB показана фиолетовым цветом, окружена митохондриями (желтый), структурами ЭР (зеленый), цистернами ЭР, расположенными друг над другом (красные стрелки), мембранные структуры внутри включения показаны белым, а ядро синим . Масштабная линейка = 2 мкм.Увеличить
Чтобы лучше понять структурные и организационные особенности включений Httex1, образующихся в клетках, мы обратились к электронной микроскопии (ЭМ). Сначала мы использовали корреляционный подход для анализа ультраструктуры включений с помощью ЭМ и их субклеточного окружения, наблюдаемого с помощью CLEM (рис. 1c, d). Положительные включения Httex1 72Q были иммуноокрашены и визуализированы с помощью конфокальной микроскопии (рис. 1c), а затем подвергнуты серийным срезам для анализа с помощью ЭМ (рис. 1d). ЭМ-микрофотографии включений Httex1 72Q выявили удивительно сложную морфологию, характеризующуюся галоподобной структурой с плотным ядром и сильно окрашенной внешней оболочкой.
Внешний слой включений содержал фибриллярные структуры, которые казались плотно упакованными и расходились от ядра включения. ЭМ-плотные цитоплазматические структуры были обнаружены как в ядре, так и на периферии и могли отражать включение или секвестрацию разрушенных органелл внутри включения 12,47,48 (рис. 1г, белые стрелки). Галоморфология включений Httex1 была одинаковой для многих включений, изображенных на изображениях (дополнительный рисунок 3a).Для дальнейшей характеристики структурных свойств и распределения Httex1 внутри включения мы использовали криофиксацию посредством замораживания под высоким давлением, которое сохраняет клеточную ультраструктуру в нативном состоянии. ЭМ-изображение выявило излучающие фибриллы на периферии и более плотно организованные и уложенные друг на друга фибриллы в ядре (дополнительная рис. 4a). Электронно-плотные мембранные структуры наблюдались в центре и более на периферии. Эти результаты позволяют предположить, что и сердцевина, и оболочка содержат агрегированные фибриллярные формы мутантного Htt, но с отчетливой структурной организацией.
Структурная организация включений Httex1 72Q в сочетании с первичной локализацией Htt-антител на их периферии позволяет предположить, что фибриллы Htt во внешнем слое этих включений служат активными центрами для рекрутирования растворимого Htt, роста фибрилл Htt и взаимодействия Htt с другими белками и клеточными органеллами.Чтобы определить, связано ли образование цитоплазматических включений с взаимодействием или рекрутированием мембранных структур, таких как ЭР и митохондрии, мы визуализировали включение-положительные клетки с помощью ЭМ в условиях, которые сохраняют внутренние мембраны клеточных органелл, т.е. в отсутствие детергентов обычно используется в процедуре ICC. ЭМ-изображения показали, что ядро и периферия включения Httex1 72Q содержат множество мелких мембранных структур (рис. 1д, желтые стрелки). Сеть ER и митохондрии присутствовали на периферии этих включений, указывая на то, что эти области могут действовать как активные сайты для рекрутирования и взаимодействия растворимого Htt с другими белками и клеточными компартментами во время образования и созревания включений.
Большинство митохондрий, окружающих цитоплазматические включения Httex1 72Q, имели поврежденные или заметно уменьшенное количество крист (рис. 1e, зеленые стрелки). Затем мы создали 3D-модель включений и органелл в их окрестностях (рис. 1e, f и дополнительный фильм 1). Трехмерная модель подтвердила, что включения формируются в переполненной области с ЭР и митохондриями вокруг нее. Трехмерная модель также предполагает, что электронно-плотные мембранные структуры, рекрутированные внутри включений Httex1, в основном состоят из эндомембран и везикул (обозначены белым цветом), что согласуется с предыдущим отчетом 9.1441 12 . Интересно, что ER принял специфическую «розетоподобную» или «уложенную стопкой цистерн» морфологию (выделено красной стрелкой) на периферии некоторых включений Httex1 72Q. Наконец, следует отметить, что митохондрии (отмечены желтым цветом, рис. 1е) были обнаружены не внутри включения, а скорее на периферии включений. Интересно, что, несмотря на их близость к ядру, включение не нарушило целостность ядерной мембраны (дополнительный рисунок 3a).Затем мы попытались идентифицировать эндомембранные компартменты внутри включений Httex1 72Q, используя панель антител или красителей, маркирующих внутриклеточные компартменты. Митохондриальные (Tom 20 и Mitotracker) и ER (BiP / Grp78) маркеры были сильно обнаружены рядом с цитоплазматическими включениями Httex1 72Q (дополнительная рис. 5a–c). Маркер потока аутофагии, p62, был обогащен на периферии включений Httex1 72Q (дополнительная рис. 5d). Маркеры образования агресом, такие как виментин и HDAC6, также были обогащены на периферии и в непосредственной близости от включений Httex1 72Q (дополнительная рис. 5e, f). Более того, когда белки цитоскелета, такие как актин и тубулин, были сверхэкспрессированы (слиты с RFP), они наблюдались в основном на периферии включений Httex1 72Q (дополнительная рис. 5g, h). Это наблюдение согласуется с колокализацией F-актина (окрашенного фаллоидином) с включениями Httex1-72Q (рис. 1b, белые стрелки). Ни один из маркеров органелл не был обнаружен внутри ядра этих включений (дополнительная рис.
5), хотя ЭМ-изображения ясно показали наличие мембранных структур в центре (рис. 1e). Это подтверждает плохую доступность антител и красителей для окрашивания ядра включений Httex1, вероятно, из-за их компактности, и подчеркивает важность использования ЭМ для расшифровки ультраструктурных свойств и состава патологических включений.В целом наши данные свидетельствуют о том, что агрегаты Httex1 могут образовываться на поверхности мембран, как это было предложено Suoponki и коллегами 49 , что может способствовать ранней агрегации, а также возмущению мембранных структур и их рекрутированию в цитоплазматические включения Httex1.
Цитоплазматические и ядерные включения Httex1 в клетках НЕК демонстрируют различные ультраструктурные свойства
Как цитоплазматические, так и ядерные включения наблюдались в мозге пациентов с БХ и в моделях трансгенных мышей с БГ 50,51,52 . Поэтому мы сравнили структурные и организационные свойства включений Httex1 72Q в цитоплазме (~85%) и ядре (~15%) трансфицированных клеток НЕК, содержащих включения 41 .
Рис. 2: Ядерные включения, образованные Httex1 72Q, не демонстрируют классической организации ядра и оболочки, наблюдаемой для цитозольных включений. Используя конфокальную микроскопию на основе иммунофлуоресценции, мы не наблюдали существенных различий в размере или общей морфологии между включениями Httex1 72Q в ядре и околоядерной области (рис. 2а). Однако ЭМ ясно показала, что ядерные включения, образованные Httex1 72Q, были обогащены фибриллярными структурами (рис. 2b), но не демонстрировали классическую организацию ядра и оболочки и не содержали мембранных структур, захваченных внутри цитоплазматического включения (рис. 1d). , д). Это говорит о том, что внутриклеточная среда является ключевым фактором, определяющим структурную и молекулярную сложность включений, и что образование ядерных и цитоплазматических включений происходит с помощью различных механизмов.a Репрезентативные конфокальные изображения ядерных включений Httex1 72Q через 48 ч после трансфекции.
Экспрессию Httex1 (серый цвет) определяли с использованием специфического первичного антитела против N-концевой части Htt (аминокислоты 1-17; 2B7 или Ab109115). Ядро контрастно окрашивали DAPI (синий), а фаллоидин (красный) использовали для окрашивания актина F. Ядерные включения Httex1 обозначены желтыми стрелками. Масштабные полосы = 10 мкм. b Электронная микрофотография репрезентативного ядерного включения Httex1 72Q. Белый квадрат указывает на область, показанную при большем увеличении на правой панели. Ядро выделено синим цветом, а двойные стрелки указывают расстояние между ядерным включением и ядерной оболочкой. Взаимодействие ядерного включения с ядерной оболочкой не наблюдалось. Масштабные полосы = 2 мкм (левая панель) и 500 нм (правая панель).Изображение в полный размер
Длина домена polyQ является еще одним ключевым фактором, определяющим формирование включений
Мы и другие ранее показали, что длина повтора polyQ сильно влияет на конформацию и свойства агрегации Httex1, при этом более высокий polyQ способствует образованию более компактного домена polyQ и ускоряет агрегацию Htt in vitro и in vivo 34,45,53 .
Однако зависимость полиQ от образования включений Htt в основном оценивалась в основном с помощью ICC, а в контексте Htt, слитого с флуоресцентными белками, полиQ повторяется намного дольше патогенного порога (64Q-9).7Q 12 ; 64Q-150Q 30 ; 43Q-97Q 14 ). Поэтому затем мы исследовали, влияет ли длина повтора polyQ также на организацию и ультраструктурные свойства цитоплазматических включений Htt в клетках HEK. С этой целью мы исследовали уровень агрегации и структурную организацию включений Httex1, несущих полиQ разной длины (16Q, 39Q против 72Q). В соответствии с предыдущими данными нашей группы, в клетках НЕК, сверхэкспрессирующих Httex1 16Q или 39, не наблюдалось значительной гибели клеток.Q строится даже через 96 часов 41 . Однако клетки, экспрессирующие Httex1 72Q, подвергались апоптозу через 96 часов. Как и ожидалось, никаких включений не образовывалось при сверхэкспрессии Httex1 16Q даже через 72 часа после трансфекции (дополнительная рис. 6). Включения Httex1 39Q обнаруживались преимущественно в цитоплазме клеток НЕК во все исследованные моменты времени (24–72 ч), хотя и в меньшем количестве, чем в условиях Httex1 72Q: Httex1 39Q (16%) против Httex1 72Q (38%). ) трансфицированных клеток 41 .Темная структура оболочки, отграничивающая ядро от периферии включений Httex1 72Q (рис. 1г, д), отсутствовала у включений Httex1 39Q (рис. 1ж). Кроме того, включения Httex1 39Q оказались менее плотными по сравнению с включениями Httex1 72Q. Эти наблюдения были одинаковыми для всех восьми включений, изображенных для каждого состояния (дополнительная рис. 3b).
Подобно тому, что мы наблюдали для Httex1 72Q, трехмерная реконструкция включений Httex1 39Q четко показала изменение организации ER, а также локализацию митохондрий вблизи включений (рис. 1h и дополнительный фильм 2). Электронно-плотные мембранные структуры, обнаруженные внутри включений, были идентифицированы как эндомембраны и везикулы. Httex1 39Клетки, экспрессирующие Q, также содержали специфические ER-цистерны на периферии включений (рис.
1h, красные стрелки). В целом, наши данные показывают, что экспансия полиQ играет решающую роль в определении окончательной архитектуры и ультраструктурных свойств включений Httex1.Удаление домена Nt17 уменьшает агрегацию мутантного Httex1, но не влияет на организацию и ультраструктурные свойства включений
Домен Nt17 функционирует как сигнал ядерного экспорта (NES), и было показано, что он играет важную роль в регуляции внутриклеточной локализация Htt, а также кинетика его агрегации и степень образования включений 36,54 . Поэтому мы стремились оценить роль Nt17 в регуляции ультраструктурных свойств включений Htt. С этой целью мы создали мутанты Httex1 39Q и 72Q, лишенные всего домена Nt17 (∆Nt17), и сравнили структурные свойства включений, образованных этими мутантами, со свойствами, образованными Httex1 39Q и Httex1 72Q. Количественная конфокальная микроскопия выявила сильное уменьшение количества включений (уменьшение ~50%) клеток, трансфицированных Httex1 ∆Nt17 72Q, по сравнению с Httex1 72Q 41 .
Удивительно, но включения, образованные Httex1 ∆Nt17 72Q (дополнительный рис. 7a, b), демонстрировали архитектуру и организацию (центральное ядро и периферийная оболочка), аналогичные тем, которые образованы Httex1 72Q (рис. 1d, e и дополнительный рисунок 8a). Кроме того, подобно Httex1 39Q, цитоплазматические включения Httex1 ∆Nt17 39Q не имеют архитектуры ядра и оболочки. Эти наблюдения указывают на то, что домен Nt17, хотя и играет решающую роль в регуляции кинетики и ранних событий агрегации Htt, не влияет на морфологию или структурную организацию цитоплазматических и ядерных включений Htt. Это удивительно, учитывая, что исследования in vitro и на клетках последовательно показали, что домен Nt17 играет важную роль в регуляции кинетики и структурных свойств агрегации Htt 11,41 и взаимодействие Htt с липидами и мембранами 8,9,55,56,57 . В недавнем исследовании с использованием твердотельной спектроскопии ядерного магнитного резонанса (ssNMR) Boatz et al. показали, что домен Nt17 и часть PRD (спирали PPII) скрыты в сердцевине фибрилл, в то время как другая часть PRD (случайная спираль) остается динамичной, доступной и регулирует мультифиламентные сборки in vitro 58 . Вместе эти наблюдения могут объяснить, почему делеция домена Nt17 не препятствует организации включения, и позволяют предположить, что взаимодействия между фибриллами и др. белками и/или органеллами, скорее всего, опосредуются гибким доменом PRD.Нейтральные липиды включаются в клеточные включения Httex1 в зависимости от длины полиQ
Хотя несколько исследований показали дисфункцию метаболизма холестерина в различных клеточных и животных моделях HD 59,60 , роль липидов в формировании включений Htt и липидный состав клеточных включений гентингтина остается неизвестным 61,62 . Чтобы лучше понять роль липидов в формировании и структурной организации включений Httex1, мы затем оценили их присутствие с помощью флуоресцентных зондов, нацеленных на разные классы липидов. Мы не наблюдали привлечение церамида, эфира холестерина или фосфолипидов во включения мутантного Httex1 (дополнительная рис. 9).а-в). Интересно, что нейтральные липиды не были обнаружены в центре включений Httex1 39Q (дополнительный рисунок 9d, белые стрелки), но были обогащены включениями Httex1 72Q (дополнительный рисунок 9e).
Это может способствовать зависящим от длины polyQ различиям в ультраструктурных свойствах включений Httex1 (рис. 1e, g). Нейтральные липиды также были обнаружены в ядерных включениях (дополнительный рисунок 9e, желтые стрелки). Хотя было показано, что домен Nt17 действует как липид- и мембраносвязывающий домен, нейтральные липиды также были обнаружены во включениях Httex1 ΔNt17 72Q, хотя и не внутри Httex1 ΔNt17 39.Включения Q (дополнительный рис. 10а). Эти результаты демонстрируют, что полиQ-зависимые взаимодействия между Htt и нейтральными липидами играют важную роль в агрегации Httex1 и образовании как ядерных, так и цитоплазматических включений. Интересно, что эти взаимодействия не зависят от домена Nt17. Остается неизвестным, происходят ли взаимодействия мутантного Httex1 с липидами до агрегации и помогают инициировать их олигомеризацию или представляют собой поздние события, связанные с созреванием включений.Количественная протеомика показывает, что образование включений Httex1 72Q включает активное рекрутирование и секвестрацию белков и органелл
растворимые фракции клеток НЕК, сверхэкспрессирующих Httex1 72Q или Httex1 16Q (см.
подробности эксперимента, дополнительные рис. 11a, c). Вестерн-блот-анализ фракции мочевины подтвердил наличие Httex1 72Q во включениях, в то время как непатогенный Httex1 16Q был обнаружен только в растворимой фракции (дополнительная рис. 11a).Как и ожидалось, белки не были значительно обогащены нерастворимой фракцией клеток НЕК, экспрессирующих Httex1 16Q, по сравнению с клетками, экспрессирующими GFP (дополнительная рис. 11d). Напротив, 377 белков были значительно обогащены нерастворимой фракцией клеток НЕК, сверхэкспрессирующих Httex1 72Q, по сравнению с клетками, сверхэкспрессирующими Httex1 16Q (рис. 3а). Среди этих белков мы идентифицировали эндогенный белок HTT (дополнительная рис. 11e). Это указывает на то, что процесс агрегации, запускаемый сверхэкспрессией Httex1, также приводит к рекрутированию эндогенного белка HTT.
Рис. 3: Протеомный анализ содержания обогащенного белка, обнаруженного в нерастворимой фракции НЕК, экспрессирующей Httex1 72Q, показывает пути агрегации белка и реакции на стресс. Белки, растворимые в мочевине, из HEK, экспрессирующих Httex1 72Q или Httex1 16Q из 3 независимых экспериментов, экстрагировали через 48 ч после трансфекции и анализировали с помощью ЖХ-МС/МС. a Идентифицированные белки были нанесены на график с использованием вулканического графика. Черные линии представляют собой порог значимости при частоте ложных открытий (FDR) < 0,05 и S0, равном 0,5, которые использовались для последующего анализа. b , c Белки, значительно обогащенные Httex1 72Q (красные точки в правой части графика вулкана), были классифицированы по клеточному компоненту ( b ) или биологическим процессам ( c ) с использованием термина Gene Ontology (GO). и обогащение DAVID (-log 10 ( p -значение) > 1). Исходные данные предоставляются в виде файла исходных данных.
Изображение полного размера
Классификация по клеточному компоненту (CC, рис. 3Bb с использованием Gene Ontology (GO) показала, что 55% белков, обогащенных нерастворимыми фракциями клеток HEK, содержащими включения Httex1 72Q, входили в состав цитоплазматического компартмента, причем 24% этих белков принадлежат эндомембранной системе, включая эндолизосомальный аппарат (покрытые клатрином эндоцитозные везикулы, ранние эндосомы, эндосомы, рециклирующие эндосомы, экзосомы и аутофагосомы), везикулы, участвующие в транспорте Гольджи-ER, и мембраны из Сеть Гольджи и транс-Гольджи.
Эти данные согласуются с данными ЭМ, показывающими, что включения Httex1 72Q состоят из небольших мембранных структур и везикул. Примерно 14% белков, обогащенных нерастворимой фракцией, были классифицированы как относящиеся к цитоскелетному компартменту. , с наиболее преобладающим актиновым цитоскелетом, что согласуется с нашими конфокальными результатами (рис. 1b). белков в растворимой фракции мочевины подтверждает, что митохондрии не изолированы внутри включений, а, скорее, накапливаются на периферии, как показано на нашей ЭМ-изображении. Остальные цитоплазматические белки, секвестрированные в нерастворимой фракции клеток HEK, трансфицированных Httex1 72Q, были в основном из перинуклеарной области или из макромолекулярных белковых комплексов, таких как убиквитин-протеасомная система (UPS), тела процессинга мРНК и стрессовые гранулы. . Интересно, что нерастворимая фракция клеток HEK, трансфицированных Httex1 72Q, также была значительно обогащена белками ядерного компартмента (~45%). Среди ядерных белков ~71,6% принадлежало нуклеоплазме. Белки из ядрышка, ядерных телец, ядерной оболочки и ядерной поры также были значительно обогащены.Классификация обогащенных белков по биологической функции показала, что наиболее обогащенные термины для биологического процесса GO (рис. 3c) и сигнальные пути с использованием анализа путей изобретательности (дополнительная рис. 12) были связаны с белками, участвующими в протеасомная убиквитинзависимая деградация белков. Белки из системы UPS 63 , включая убиквитиновую часть, убиквитинлигазы E3 (например, ITCH, RNF34, TRIM32), субъединицы 26S протеасомы и деубиквитиназы сформировали основной кластер дифференциально обогащенных белков во включениях Httex1 72Q. Эти результаты согласуются с предыдущими исследованиями, показывающими, что включения Htt, сформированные у пациентов с БХ, или в нескольких клеточных и in vivo моделях секвестрируют несколько ключевых компонентов системы UPS, включая протеасомы 26S 64 (ADRM1 65 ), деубиквитиназы 66 (например, NEDD4 и USP5) и убиквитинлигазы Е3 (например, ITCH, TRAF6, UBE3A, UHRF2 и Parkin) и индуцируют нарушение ИБП 67,441 ,69 .
Кроме того, наш анализ показал, что среди белков, значительно повышающих или понижающих уровень регуляции в Httex1 72Q по сравнению с Httex1 16Q, 42 белка являются известными Htt-интеркторами, связанными с UPS, идентифицированными из базы данных HDinHD (дополнительные данные 1).Белки, участвующие в образовании аутофагосом (оптинеурин 70 ), созревание (убиквилины 71 ) и процесс слияния аутофагосом-лизосом 72 (TOLLIP-взаимодействующие белки и белки, участвующие в эндолизосомном переносе) также были обогащены внутри включений Httex1 72Q. И TOLLIP 73,74,75 , и оптинейрин 76,77,78,79 имеют решающее значение для эффективного удаления агрегатов белка полиQ 80 и, в частности, для деградации агрегатов Htt. Истощение TOLLIP в клетках HeLa увеличивает индуцированную GFP-Htt-103Q токсичность 73 , в то время как нокдаун оптинейрина способствует агрегации Htt.
Кроме того, несколько шаперонов из семейств Hsp70 и DnaJ/HSp40 (например, DNAJB6, DNAJB2 и другие белки из семейств Hsp40 и Hsp70), а также убиквилин-2, обнаруженные ранее обогащенными включениями Htt 30, 81,82 , также были обогащены включениями Httex1 72Q.
Этот кластер белков был связан с сигнальным путем BAG2 (дополнительный рисунок 12), одним из главных канонических сигнальных путей, регулирующих взаимодействие между шаперонами из семейства Hsp70/Hsc70 и убиквитином. Интересно, что недавно было показано, что комплекс Hsp70 вместе с семейством шаперонов Hsp40/110 образует комплекс дезагрегазы, который может напрямую связываться с агрегатами Htt 83 . После дезагрегации Ubiquilin-2 взаимодействует с Htt и перемещает дезагрегированные виды к протеасомам, чтобы способствовать их полной деградации 84 . Обогащение сети шаперонов дезагрегации во включениях Httex1 72Q указывает на то, что они были активно рекрутированы, но не смогли очистить агрегаты Httex1 72Q. Наш протеомный анализ также выявил обогащение нескольких биологических процессов и сигнальных путей, связанных с белками, связывающими РНК, факторами транскрипции, сплайсингом РНК, процессингом и стабильностью мРНК, а также белками, связывающими хроматин и нуклеотиды (рис. 3с и дополнительная рис. 12). ). Нарушение регуляции путей транскрипционных генов было зарегистрировано у нескольких животных и клеточных моделей HD 85,86,87 а также в посмертных тканях БГ и клетках периферической крови БГ 88,89,90 .Вместе наши протеомные данные и данные CLEM обеспечивают убедительные доказательства того, что образование включения Httex1 72Q включает активное рекрутирование и секвестрацию клеточных белков, липидов и органелл. Это также свидетельствует о том, что секвестрация регуляторов транскрипции и истощение ключевых белков аутофаголизосомного, UPS и шаперонного путей из-за их секвестрации внутри патологических включений Httex1 вносят основной вклад в клеточную дисфункцию и нейродегенерацию при БХ, как сообщалось в мозге БХ человека. ткань 91 .
Образование цитоплазматических включений Httex1 72Q индуцирует фрагментацию митохондрий, усиливает митохондриальное дыхание и приводит к ремоделированию сайта выхода ER
Заметное накопление поврежденных митохондрий на периферии включений Httex1 72Q побудило нас исследовать, как образование включений Htt влияет на функции митохондрий.
Рис. 4: Формирование включений Httex1 72Q вызывает митохондриальные изменения и уменьшение количества сайтов выхода ER. Количественная оценка длины митохондрий по ЭМ-микрофотографиям выявила более короткий митохондриальный профиль, связанный с включениями Httex1 72Q, по сравнению с клетками НЕК, трансфицированными пустым вектором (рис. 4а, б). Сходные уровни белка внешней митохондриальной мембраны VDAC1 предполагают, что эта митохондриальная фрагментация не была связана с уменьшением плотности митохондрий (дополнительная рис. 13a). Мы предположили, что фрагментация и рекрутирование митохондрий во включения Httex1 могут быть связаны с дыхательной дисфункцией. Поэтому мы провели респирометрию с высоким разрешением на клетках, трансфицированных Httex1 16Q и 72Q, в течение 48 часов. Агрегаты были обнаружены только в клетках, трансфицированных мутантным Httex1 72Q (дополнительная рис. 13b). Мы оценили различные состояния дыхания митохондрий с помощью респирометрии высокого разрешения (дополнительная рис. 13c-d). Трансфекция Httex1 72Q приводила к значительно более высокому митохондриальному дыханию, чем Httex1 16Q (дополнительная рис. 4c). Вместе наши результаты демонстрируют явное влияние образования включений Httex1 72Q на митохондриальную морфологию и функцию.a Электронные микрофотографии митохондрий в клетках НЕК со сверхэкспрессией пустого вектора (EV) или Httex1 72Q. На вставках показаны увеличенные изображения митохондрий, обнаруженных на периферии включений Httex1 72Q или в контроле EV. Масштабные полосы = 1 мкм. b Измерение длины митохондрий с помощью FIJI показывает значительное уменьшение размера митохондриального профиля, расположенного вблизи включений. Непарный 9Был проведен тест 0012 t (двусторонний), в результате чего значение p составило 0,0006. Был проведен непарный t-критерий (двусторонний), в результате чего значение p составило 0,0006. c Клетки HEK из 4 независимых экспериментов осторожно отделяли для респирометрии высокого разрешения (HRR) через 48 часов после трансфекции указанными конструкциями.
HRR проводили в дыхательных средах. После измерения обычного дыхания клетки пермеабилизировали дигитонином, а затем индуцировали различные состояния дыхания с использованием протокола титрования субстрат-разобщающий-ингибитор (SUIT). Обычное дыхание, НАДН-зависимое или сложное 1-связанное дыхание после добавления АДФ (состояние OXPHOS) (NP), НАДН- и сукцинатное или сложное 1- и 2-связанное дыхание в состоянии OXPHOS (NSP) и в оценивали мощность несвязанной электрон-транспортной системы (ETS) (NSE), а также сукцинат-зависимое или сложное 2-связанное дыхание в состоянии ETS (SE). Графики представляют собой среднее значение ± SD для 4 независимых экспериментов. Двухфакторный дисперсионный анализ показывает значительную взаимосвязь между состояниями дыхания и длиной повтора полиQ ( p -значение = 0,004). d Репрезентативные конфокальные изображения клеток HeLa, трансфицированных Httex1 16Q, 39Q или 72Q или EV. Клетки фиксировали через 48 ч после трансфекции и иммуноокрашивали. Httex1 был обнаружен с помощью антитела MAB5492 Htt (серый), а сайты выхода ER (ERES) были обнаружены с помощью Sec13 (красный). Масштабные полосы = 20 мкм. e , f Количество ERES ( e ) и размер ( f ) Количественные оценки по конфокальной визуализации были выполнены с использованием FIJI. На графиках представлено среднее ± ± стандартное отклонение 3 независимых экспериментов, представленное в процентах относительно контроля Httex1 16Q. Был проведен однофакторный дисперсионный анализ с последующим апостериорным тестом честной значимой разницы Тьюки [HSD] (9).0012 p – значения < 0,0001 для панелей e и f). * P < 0,05, ** P < 0,005, *** P < 0,001 для множественных сравнений.Изображение в натуральную величину
Наш ЭМ-анализ и 3D-реконструкция дополнительно выявили наличие цистерн ЭР на периферии включений Httex1. Сложенные цистерны ER обычно образуются при увеличении концентрации специфических резидентных белков или в условиях стресса 92 .
Изменения, которые мы наблюдали в организации ER, вместе с обогащением белков, связанных с переносом ER-Golgi внутри включений, побудили нас исследовать, были ли нарушены функции ER при образовании включений Httex1.Основной функцией ER является биогенез носителей COPII, которые переносят белки и липиды в дистальные компартменты. Носители COPII образуются в местах выхода ER (ERES), которые представляют собой свободные от рибосом домены шероховатого ER. Чтобы определить, мешает ли образование включений Httex1 гомеостазу ERES, мы использовали конфокальную визуализацию и количественно определили количество ERES, специально помеченных белком Sec13 компонента COPII в клетках HeLa, экспрессирующих Httex1 72Q, Httex1 39Q или Httex1 16Q (рис. . 4г). Наши данные показывают, что количество ERES было значительно снижено (~ 20%) только в клетках, содержащих включения Httex1 72Q (рис. 4д). Затем мы количественно оценили размер ERES, используя тот же конвейер визуализации (рис. 4f). Сверхэкспрессия Httex1 16Q вызывала уменьшение размера ERES на 20% по сравнению с пустым вектором (EV).
Однако снижение стало гораздо более значительным в клетках, несущих Httex1-39.Q или Httex1 72Q с уменьшением на ~40% по сравнению с пустым вектором и на ~20% по сравнению с Httex1 16Q. Эти результаты показывают, что образование включений Httex1 препятствует образованию и слиянию ERES. Ремоделирование ERES в клетках было описано прежде всего как адаптивный ответ на уровень синтеза белка ER с количеством ERES, пропорциональным нагрузке груза. Интересно, что наши результаты протеомики (рис. 3а) показали, что белок TGF (трансформирующий фактор роста), который играет центральную роль в биогенезе и организации ERES, секвестрирован во включениях мутантного Httex1 93 . В соответствии с нашими результатами ранее было показано, что истощение TGF индуцирует резкое уменьшение количества сайтов ERES 94 . Альтернативно, уменьшение количества сайтов ERES может отражать ранние признаки клеточной уязвимости и токсичности, вызванные присутствием включений Httex1 в клетках.Экспрессия Httex1 72Q в первичных нейронах коры приводит к образованию плотных и нитевидных ядерных включений
Затем мы исследовали детерминанты последовательности агрегации Htt и образования включений в первичных корковых нейронах с течением времени.
Рис. 5: Анализ конфокальной микроскопии и классификация включений Httex1, образующихся в нейронах, выявили их морфологическую гетерогенность. Как и ожидалось, ни одна из конструкций Httex1 16Q (Httex1 16Q и ΔNt17 Httex1 16Q), сверхэкспрессированных в нейронах, не индуцировала образование агрегатов до 14 дней после трансдукции (D14). Напротив, сверхэкспрессия Httex1 72Q индуцировала образование круглых ядерных включений почти в 100% трансдуцированных нейронов уже на D3 (рис. 5a, b и S14). Менее 1% нейронов имели цитоплазматические включения либо в виде точек, либо кольцевидной морфологии. Хотя никаких изменений в субклеточном распределении включений не наблюдалось до D14, мы наблюдали значительные изменения размера и формы ядерных включений с течением времени (рис. 5c). На D3 большинство ядерных включений были обнаружены в виде мелких (<1 мкм) ядерных точек (94%), и лишь немногие проявлялись в виде крупных (~3 – 4 мкм) включений (рис. 5г). Соотношение мелких и крупных ядерных включений немного сместилось со временем от 50:50 на Д7 до 43:57 на Д14 (рис. 5г). a Экспрессия Httex1 была обнаружена с помощью окрашивания ICC в сочетании с конфокальной визуализацией в первичных корковых нейронах через 3 (D3), 7 (D7) и 14 (D14) дни после лентивирусной трансдукции. Мутанты Httex1 были обнаружены с помощью MAB549.2 антитела. Ядро контрастировали с помощью DAPI (синий), а MAP2 использовали для визуализации нейронов (красный). Масштабные полосы = 20 мкм. b Количественная оценка количества нейронов, содержащих включения Httex1, с течением времени на основе изображений. На графиках представлено среднее значение ± стандартное отклонение для трех независимых экспериментов. c Первичные нейроны классифицировали по обнаружению Httex1 как диффузные или по морфологии обнаруженных агрегатов Httex1: (1) мелкие ядерные точки; (2) по крайней мере одно крупное ядерное включение и (3) цитоплазматическое включение. Кроме того, был создан подкласс для нейронов, содержащих крупное ядерное включение, связанное с ядерной конденсацией. Масштабная линейка = 20 мкм.
d Количественная оценка и классификация различных морфологий включений Httex1 на основе изображений на основе панели c . Минимум n = 145 клеток на условие было исследовано в течение 3 независимых экспериментов. Данные представлены в виде средних значений + /– SD. Статистический анализ: был проведен однофакторный дисперсионный анализ с последующим апостериорным тестом Тьюки [HSD]. p – значения < 0,0001 для верхней и нижней панелей и * P < 0,05, ** P < 0,005, *** P < 0,001 для множественных сравнений. e Количественная оценка нейронов, содержащих большое ядерное включение с ядерной конденсацией, на основе изображений. Минимум n = 145 клеток на условие было исследовано в течение 3 независимых экспериментов. Данные представлены в виде средних значений + /– SD. Статистический анализ: двухфакторный дисперсионный анализ не выявил значительного взаимодействия ( p -значение = 0,0598), но значимые выборки фактора строки ( p -значение = 0,0076) и фактор времени столбца ( p -значение = 0,0002) с * P < 0,05, ** P < 0,005, *** P < 0,001 для множественных сравнений.Изображение с полным размером
Затем мы охарактеризовали ультраструктурные свойства ядерных включений Httex1 72Q с помощью CLEM. На D7 эти включения выглядели как плотные и примерно круглые агрегаты без отчетливой структурной организации ядра и оболочки (рис. 6a и S15a). В этих тонких срезах внутриядерные включения Httex1 72Q казались темнее окружающей нуклеоплазмы и структурно отличались от ядрышка (рис. 6а и контрольные дополнительные рис. 15в, г). Из-за высокой плотности этих агрегатов было сложно определить, состоят ли они из нитевидных структур. Однако с помощью электронной томографии (ЭТ) мы смогли визуализировать и подтвердить наличие нитевидных структур внутри этих ядерных включений (рис. 6b и дополнительный фильм 3). Сегментированные филаменты, по-видимому, не были тесно уложены параллельно друг другу, а скорее были организованы в виде сети извилистых филаментов. Ни ЭМ, ни ЭТ изображения не выявили наличия мембраноподобных, органеллоподобных или везикулоподобных структур внутри или на периферии этих включений.
Рис. 6: Ультраструктурный анализ ядерных включений Httex1 72Q, образованных в первичных нейронах коры, показывает зернистые и нитевидные структуры с помощью CLEM и томографии. Хотя удаление домена Nt17 (рис. 6c) не изменило ультраструктурные свойства или состав включений, оно значительно ускорило образование крупных агрегатов. Уже в D3 ~ 60% нейронов, сверхэкспрессирующих ΔNt17 Httex1 72Q, уже содержали большие ядерные включения по сравнению только с ~ 6% для Httex1 72Q (рис. 5d). На 14-й день почти все агрегаты, образовавшиеся в гиперэкспрессирующих нейронах ΔNt17 Httex1 72Q, превратились в крупные ядерные включения (~90%) (рис. 5d) по сравнению с 57% для Httex1 72Q.a Репрезентативные электронные микрофотографии включений Httex1 72Q, образовавшихся через 7 дней после лентивирусной трансдукции в первичных нейронах коры головного мозга мыши. Синие пунктирные линии обозначают включения, а красные стрелки — ядрышки. Масштабные полосы = 500 нм для ЭМ-изображений и 50 мкм для флуоресцентного изображения (вставка).
b Электронная микрофотография и соответствующая томограмма (выделенная область красного цвета) нейронного включения Httex1 72Q на панели a. Сегментация томограммы выявляет наличие множества нитей. c Репрезентативные электронные микрофотографии включений ΔNt17 Httex1 72Q, образовавшихся через семь дней после лентивирусной трансдукции в первичных нейронах коры головного мозга мыши. Оранжевые пунктирные линии обозначают включения, а красные стрелки обозначают ядрышки. Масштабные полосы = 500 нм для ЭМ-изображений и 50 мкм для флуоресцентного изображения (вставка).Изображение в полный размер
Наконец, среди мелких агрегатов, рассеянных по всему ядру, несколько наблюдалось вблизи ядерной мембраны, которая часто выглядела поврежденной и разорванной (рис. S15a). Наши данные согласуются с предыдущими исследованиями ЭМ у пациентов с HD 95 и моделей HD мышей 19,96 , демонстрирующих ядерные ультраструктурные изменения, включая измененную форму ядерной мембраны, ядерную инвагинацию и повышенную плотность ядерных пор в нейронах, несущих включения Httex1.
В соответствии с этими наблюдениями, в ядрах, содержащих включения Httex1 72Q, со временем наблюдалась повышенная ядерная конденсация (рис. 5e), что указывает на повышенную токсичность для нейронов, что согласуется с предыдущими отчетами у пациентов с БХ 97 и мыши модели HD 96 . Интересно, что, несмотря на наличие крупных включений уже в день D3 в нейронах со сверхэкспрессией ΔNt17 Httex1 72Q, мы не наблюдали более раннего начала гибели клеток или более высокого уровня токсичности с течением времени в этих нейронах по сравнению с нейронами со сверхэкспрессией Httex1 72Q (рис. . 5д). Это свидетельствует об отсутствии корреляции между уровнем гибели клеток и размером включений, при этом крупные включения менее токсичны.Эти результаты демонстрируют образование конденсированных фибриллярных агрегатов мутантного Httex1 внутри внутриядерных включений в нейронах и позволяют предположить, что процесс их формирования и созревания напрямую связан с дисфункцией нейронов и гибелью клеток.
Добавление GFP к С-концевой части Httex1 индуцирует дифференциальную структурную организацию и токсические свойства включений Httex1 агрегационные свойства мутантного Httex1, слитого с GFP (Httex1 72Q-GFP и Httex1 39Q-GFP). Сначала мы оценили морфологию цитоплазматических включений Httex1 72Q-GFP в клетках HEK с помощью ICC с использованием панели антител Httex1 (рис. 7a, b и S16a). Конфокальная визуализация выявила диффузный сигнал GFP во включениях. Напротив, антитела Htt метили в основном самую внешнюю область включений Httex1 72Q-GFP, но также слабо окрашивали их центры (рис. 7b). Напротив, ядра включений без меток не были помечены всеми протестированными антителами Htt. Таким образом, мы предположили, что наличие метки GFP приводит к образованию менее компактных включений Httex1. Хотя было обнаружено, что Actin-F колокализуется с включениями Httex1 без меток [(Fig. 1b, Httex1 72Q), 39Q (дополнительный рисунок 9d, Httex1 39Q) и ΔNt17 Httex1 72Q (дополнительный рисунок 10a, ΔNt17 Httex1 72Q)] специфического обогащения актином-F в ядре или на периферии включений Httex1 72Q-GFP обнаружено не было.
Кроме того, актиновые филаменты были обнаружены исключительно на периферии цитоплазматических включений без метки Httex1 72Q и 39Q (+/- ΔNt17) (рис. 1b и S10A). Эти результаты указывают на потенциальную роль актина в формировании или созревании включений Httex1. В соответствии с этими наблюдениями, актиновый цитоскелет был одним из наиболее нерегулируемых путей согласно протеомному анализу, проведенному в мозге людей с БХ 98 , подчеркивая его важность в развитии патологии ГБ. Рис. 7: Добавление GFP к С-концевой части Httex1 индуцирует дифференциальную структурную организацию, обнаруженную с помощью ICC и CLEM. a Картирование эпитопов антител Httex1. b Включения Httex1 72Q-GFP, образовавшиеся через 48 ч после трансфекции в клетках НЕК, были обнаружены с помощью окрашивания ICC в сочетании с конфокальной визуализацией. Все антитела Htt показали сильную иммунореактивность к периферии включений Httex1 и умеренную иммунореактивность к сердцевине.
Ядро контрастировали DAPI (синий), а F-актин – фаллоидином (красный). Масштабные полосы = 20 мкм (левые панели) и 10 мкм (средняя и правая панели). c , d Через 48 ч после трансфекции клетки HEK фиксировали, выполняли ICC против Httex1 и визуализировали с помощью конфокальной микроскопии ( c ). Масштабные полосы = 5 мкм. Затем выбранную область клеток (белый квадрат) исследовали с помощью ЭМ ( d ). Бинарное изображение (вставка) было создано из электронной микрофотографии с использованием медианной фильтрации и порога интенсивности Оцу, что позволяет лучше различать морфологию включений. Масштабные полосы = 500 нм. e Репрезентативные конфокальные изображения ядерных включений Httex1 72Q-GFP, образовавшихся в клетках HEK через 48 ч после трансфекции. Экспрессия Httex1 (серый цвет) была обнаружена с использованием специфического антитела против N-концевой части Htt (аминокислоты 1–17; 2B7 или Ab109).115), а GFP (зеленый) визуализируется непосредственно в соответствующем канале. Ядро докрашивали DAPI (синий), а фаллоидин (красный) использовали для окрашивания филаментного актина. Ядерные включения Httex1 обозначены желтыми стрелками. Масштабные полосы = 10 мкм. f Электронная микрофотография репрезентативного включения Httex1 72Q-GFP. Белый квадрат указывает на область, показанную при большем увеличении на правой панели. Ядро выделено синим цветом. Масштабные полосы = 2 мкм (левая панель) и 500 нм (правая панель).Изображение полного размера
Далее мы провели более глубокий анализ клеточных включений Httex1 72Q-GFP, сформированных в HEK с помощью CLEM (рис. 7c, d). Включения Httex1 72Q-GFP были организованы в виде очень плотной сети фибрилл, которые были более гомогенно окрашены (рис. 7d и S16c) и не демонстрировали архитектуру ядра и оболочки, характерную для включения Httex1 72Q без метки (рис. . 1d и S16d). Более внимательное изучение включений показало, что они состоят из плотно упакованных фибрилл, которые демонстрировали поразительное сходство с фибриллярными агрегатами, образованными мутантными белками Httex1 в бесклеточной системе (рис.
7d и S16b). Структурный анализ Httex1 72Q-GFP с помощью замораживания под высоким давлением (дополнительная рис. 4b) также выявил плотно упакованные фибриллы, исходящие из включений. Центр включений не был хорошо разрешен, но можно было четко наблюдать более толстые фибриллы, расходящиеся по периферии с увеличенным расстоянием между ними по сравнению с включением Httex1 72 без метки (дополнительная рис. 4a). Часть включений Httex1 72Q-GFP имела перинуклеарную локализацию. В некоторых случаях скопление фибрилл вблизи ядерной мембраны приводит к явному искажению ядра, но без разрушения мембраны (рис. 7d и S17a). В целом, мы не наблюдали существенных различий в диаметре и расстоянии от ядра для всех включений Httex1 (+/- GFP), визуализированных в клетках HEK (дополнительная рис. 18).Затем мы исследовали ультраструктурные свойства включений Httex1 72Q-GFP, присутствующих в ядре клеток HEK (рис. 7e, f). Никаких существенных различий с точки зрения организации между ядерными и цитоплазматическими включениями Httex1 72Q-GFP не наблюдалось как при обнаружении GFP, так и при окрашивании антителами (рис.
7e). Ядерные включения Httex1 72Q-GFP также были обогащены фибриллярными структурами. Трехмерная реконструкция, полученная из серии электронных микрофотографий, выявила гораздо меньше мембранных структур как в Httex1, так и в 39Включения Q-GFP и Httex1 72Q-GFP (дополнительная рис. 19) по сравнению с включениями Httex1 без меток (рис. 1f, h). Более того, в соответствии с наблюдениями ЭМ, во включениях Httex1 39Q-GFP и Httex1 72Q-GFP не было обнаружено нейтральных липидов (дополнительная рис. 10b-c). Интересно, что ни длина повтора polyQ, ни присутствие или удаление домена Nt17, по-видимому, существенно не изменяют размер, морфологию или структурные свойства включений, образованных мутантными белками Httex1, слитыми с GFP (дополнительная рис. 20). Это отличается от того, что мы наблюдали для клеток, экспрессирующих tag-free Httex1, в которых увеличение длины полиQ приводило к включениям с отличной морфологией и особенностями организации. Эти данные свидетельствуют о том, что добавление GFP значительно изменяет механизм агрегации Httex1 и образования включений.Чтобы получить еще более четкое представление об этих различиях, мы затем выполнили ЭМ в условиях без моющих средств, чтобы сохранить внутренние мембраны и структуры включений. Мы заметили, что длина повтора polyQ не влияла на размер включений Httex1 39Q-GFP и 72Q-GFP (дополнительная рис. 18b) или их общую архитектуру в клетках HEK. Включения Httex1 39Q-GFP состоят из излучающих фибрилл и, таким образом, похожи на включения Httex1 72Q-GFP, хотя и немного менее плотные. Трехмерная реконструкция включений показала наличие ER и митохондрий на их периферии, но несколько мембранных структур были интернализованы (дополнительная рис. 19).b желтая стрелка) по сравнению с включениями Httex1 39Q и Httex1 72Q без тегов. Аналогичный анализ клеточных включений Httex1 ΔNt17 39Q-GFP и Httex1 ΔNt17 72Q-GFP (рис. S20a–d) также не выявил влияния делеции Nt17 на ультраструктуру включений или их взаимодействие с окружающими органеллами. Эти результаты подтверждают, что механизмы агрегации и образования включений мутантного Httex1 значительно изменяются при добавлении GFP.
Мы также оценили, как присутствие метки GFP может влиять на кинетику агрегации, морфологию, субклеточную локализацию и токсичность ядерных включений в первичных нейронах коры (рис. 8 и S21). Во-первых, мы наблюдали, что присутствие метки GFP замедляет скорость агрегации Httex1 72Q в отличие от Httex1 72Q без метки, агрегация значительно замедляется, о чем свидетельствует отсутствие ядерных или цитоплазматических агрегатов или включений на D3 ( Рис. 8а, б). Однако в день 7 почти все белки Httex1 72Q-GFP появлялись в виде небольших ядерных точек (~ 35%) или крупных ядерных включений (~ 60%) (рис. 8c). Доля мелких точек по сравнению с крупными включениями не менялась с течением времени вплоть до дня 14 (рис. 8в). Интересно, что присутствие метки GFP не влияло на субклеточную локализацию, морфологию и распределение по размерам включений, наблюдаемых с помощью конфокальной визуализации. Однако агрегаты Httex1 72Q-GFP показали значительно меньшую токсичность по сравнению с Httex1 72Q без метки в нейронах (дополнительная рис.
Рис. 8: Метка GFP изменяет кинетические, ультраструктурные свойства, содержание белка и токсичность включений Httex1 72Q в нейронах. 21d). Даже на 14-й день конденсация хроматина в этих нейронах не превышала 10% (дополнительная рис. 21d), тогда как ~ 50% нейронов, сверхэкспрессирующих конструкцию Httex1 72Q без метки, уже были мертвы (рис. 5e). Кроме того, анализ гибели клеток TUNEL выявил резкое увеличение (~ 60%) фрагментации ДНК в нейронах коры, несущих внутриядерные включения Httex1 72Q, по сравнению с только 40% в нейронах, экспрессирующих Httex1 72Q-GFP (рис. 8d).a Httex1 72Q-GFP непосредственно визуализировали с помощью GFP (зеленый) через 3 (D3), 7 (D7) и 14 (D14) дни после трансдукции лентивируса. Ядра контрастировали DAPI, а нейроны окрашивали антителами MAP2. Масштабная линейка = 20 мкм. b Количественная оценка количества трансдуцированных нейронов, несущих включения Httex1 72Q-GFP, на основе изображений с течением времени.
c Морфологическая и субклеточная классификация локализации (см. рис. 5c) включений 72Q-GFP, образованных в трансдуцированных нейронах. n = 219 клеток исследовали в 3 независимых экспериментах. d Анализ гибели клеток TUNEL в нейронах коры, трансдуцированных Httex1. b , c , d Графики представляют собой среднее ± ± стандартное отклонение трех независимых экспериментов. b , d Был проведен односторонний ANOVA с последующим апостериорным тестом HSD. * P < 0,05, ** P < 0,005, *** P < 0,001 для множественных сравнений. # P < 0,05 (72Q по сравнению с 72Q-GFP). c Двухфакторный дисперсионный анализ с повторными измерениями не выявил существенных различий. e Репрезентативные ЭМ-изображения включений Httex1 72Q-GFP, образовавшихся в нейронах коры головного мозга после трансдукции D7. Зеленые пунктирные линии представляют включения Httex1. Масштабные полосы = 500 нм и 50 мкм (вставка). f-i Белки, растворимые в мочевине, экстрагированные из нейронов коры, экспрессирующих Httex1 72Q или Httex1 16Q ( f ) или Httex1 72Q-GFP или GFP ( h ) на D7 после трансдукции анализировали с использованием ЖХ-МС/МС. n = 3 независимых эксперимента. Идентифицированные белки были нанесены на график с использованием вулканических графиков (f, Httex1 72Q по сравнению с Httex1 16Q; h, Httex1 72Q-GFP по сравнению с GFP). Пунктирные линии представляют частоту ложных открытий <0,05, и был назначен порог абсолютного логарифмического 2-кратного изменения значимости, равный 1. Активированные белки представлены красным цветом, подавленные - синим, а несущественные белки - серым. г Онлайн-платформа STRING подчеркивает связь между активированными белками Httex1 72Q и белками, связанными с путями деградации (красный кружок). i Диаграмма Венна, сравнение белков, обогащенных Httex1 72Q, по сравнению с Httex1 16Q (синий) и Httex1 72Q-GFP по сравнению с Httex1 16Q (зеленый). 24 белка были уникальными для Httex1 72Q по сравнению с Httex1 16Q (44,4%), 17 уникальными для Httex1 72Q-GFP по сравнению с Httex1 16Q (31,5%) и 13 (24,1%) общими для обоих коагрегированных белков. Исходные данные предоставляются в виде файла исходных данных.Полноразмерное изображение
Затем мы исследовали, как присутствие метки GFP может влиять на ультраструктурные свойства этих включений нейронов (рис. 8e и S22). Ядерные включения, образованные в присутствии метки GFP, имели круглую форму, но демонстрировали меньшую плотность темного окрашивания по сравнению с включениями, образованными немеченым белком Httex1 72Q (рис. 6а). Из-за их более низкой плотности можно было обнаружить присутствие нитевидных структур, организованных в виде запутанного расположения по всему включению, меченному GFP. Наконец, эти включения были локализованы по всему ядру, но всегда удалены от ядерной мембраны (дополнительная рис. 22).
В целом, наши результаты показывают, что добавление GFP значительно замедляет инициацию фибрилляции мутантного Httex1 и образование включений в ядре, но ускоряет созревание однажды сформированных агрегатов, что приводит к преимущественно большим включениям с пониженной токсичностью в нейронах.
Отсутствие физического взаимодействия между включениями Httex1 72Q-GFP и ядерной оболочкой и/или отсутствие агрегатов небольших точек в нейронах, экспрессирующих Httex1-72Q-GFP, может объяснить их пониженную токсичность (рис. 8d, e).Ядерные и цитоплазматические включения Httex1 72Q и Httex1 72Q-GFP демонстрируют разный состав протеома. содержание ядерных включений, образованных в первичных нейронах (дополнительная рис. 23). Протеомный анализ показал, что 23 белка были значительно обогащены включениями Httex1 72Q по сравнению с Httex1 16Q (рис. 8f). Эти белки были проанализированы с помощью ассоциации белок-белок с использованием онлайн-платформы STRING (рис. 8g) и классифицированы с использованием анализа терминов GO по клеточному компоненту, молекулярной функции и биологическому процессу (дополнительный рисунок 24). Идентифицированные белки включают факторы транскрипции PCBD2 и Mlf2, белок, связывающий РНК/ДНК (GNL3), ДНК-зависимую РНК-полимеразу (Polr2c), гистон-лизин-деметилазу (No66), эксцизионную репарацию нуклеотидов генома ДНК (Rad23B) и белки, связанные с сегрегация хромосом (ARL8B и Tubb2a).
В соответствии с нашими результатами, субъединицы ARL8B, Tubb2A, Polr2, белки Mlf2, GNL3 и Rad23B также были обнаружены в нерастворимых фракциях мозга мышей Q175 HD 99 и Tubb2a у мышей R6/2 100 ядерные включения. Интересно, что уровень экспрессии генов этих ядерных белков также был значительно повышен в посмертном мозге HD (PCBD2) , 101, и у пациентов с симптомами HD (Mlf2, GNL3, Polr2 и ARL8B) , 102, . В целом это указывает на то, что потеря ключевых ядерных белков из-за их секвестрации патологическим включением Htt компенсируется в нейронах повышением уровня экспрессии генов. Однако этого кажется недостаточным, чтобы компенсировать потерю их биологических функций, о чем свидетельствует высокий уровень ядерных изменений, наблюдаемых в нашей нейронной модели HD, включая конденсацию хроматина, фрагментацию ядра и потерю целостности ядерной оболочки (рис. 6).В дополнение к ядерным белкам, STRING-анализ (рис. 8g) вместе с классификацией клеточных компонентов GO (дополнительный рис.
24a), молекулярные функции (дополнительный рис. 24B) и аннотации биологических процессов (дополнительный рис. . 24c) выявили обогащение кластера белков [убиквилин (ubqln) 1, 2 и 4; Rps27a, Rad23B и Nedd8, красный кружок, рис. 8g], связанные с механизмом деградации белка, включая пути UPS, аутофагии и ERAD. В соответствии с нашими выводами, все эти белки также были обнаружены в нерастворимых фракциях мозга мышей линии Q175 HD, в то время как члены семейства убикилинов (Ubqln 1, 2 или 4) 103,104,105 также были секвестрированы в ядерных включениях мышей R6/2 100 и в ядерных включениях Httex1, образованных в предшественнике нейронов крысы PC12 106 . Интересно, что ранее было показано, что белки ubiquilin перемещаются из цитоплазмы в ядро или имеют повышенный уровень экспрессии в нуклеоплазме одновременно с ранней стадией образования нейрофибриллярных клубков в посмертных тканях мозга AD , 107, . Более того, Nedd8, изначально показанный для колокализации с убиквитином и протеасомными компонентами в цитоплазматических включениях 108,109 обнаруженный при нескольких нейродегенеративных заболеваниях, недавно было показано, что он способствует агрегации ядерных белков в качестве защитного механизма от протеотоксичности 110 . Кроме того, наш анализ выявил 5 белков, известных как интеракторы Htt (на основе базы данных HDinHD), среди белков, значительно отличающихся между Httex1 72Q и Httex1 16Q в первичных нейронах (дополнительные данные 1).Наконец, мы также оценили, как присутствие метки GFP может влиять на содержание белка во внутриядерных включениях нейронов (рис. 8h). Подобно агрегатам Httex1 72Q, ~45% белков, обогащенных нерастворимой фракцией агрегатов Httex1 72Q-GFP, были связаны с ядерными биологическими процессами и функциями, включая факторы транскрипции (Mlf2 106 , nfyc-1, TCERG1, Med15), белки, связывающие ДНК-хроматин (ZBED5, N6AMT1 и Actl6b), белок эксцизионной репарации нуклеотидов генома ДНК (Rad23B) и ДНК/РНК-связывающий белок FUS. Кроме того, агрегаты Httex1 72Q-GFP также были обогащены белками из ядерных телец промиелоцитарного лейкоза (Sumo 1 и 2), известных как ядерные безмембранные компартменты, участвующие в поддержании генома, таком как репарация ДНК, реакция на повреждение ДНК, гомеостаз теломер, а также связаны с сигнальными путями апоптоза (дополнительная рис.
25). Кроме того, белки убиквитин-протеасомной системы (Bag5, Fus, Rad23b, Sgta, Sumo1, Sumo2, Ubqln1, Ubqln2, Ubqln4), ERAD (Sgta, Ubqln1, Ubqln2), аутофагии (Ubqln1, Ubqln2, Ubqln4) и сумоилирования (Sumo1, Sumo2) пути также были сильно обогащены нерастворимой фракцией агрегатов Httex1 72Q-GFP (дополнительная рис. 25a, b). Интересно, что протеом агрегатов Httex1 72Q-GFP, образованных в нашей модели нейронов HD, разделял 40% белков, также обнаруженных обогащенными нерастворимой фракцией ядерных включений Httex1 74Q-GFP, образованных в клеточной линии предшественников нейронов крысы PC12 106 . Это подчеркивает сильное сходство между путями и белками, участвующими в агрегации Httex1 и образовании включений в нейрональных клетках грызунов.Затем мы сравнили белки, идентифицированные между включениями Httex1 72Q без метки и Httex1 72Q-GFP (рис. 8i). 24 белка были уникальными для Httex1 72Q (44,4%), 17 уникальных для Httex1 72Q-GFP (31,5%) и только 13 (24,1%) были общими для обоих типов включений.
Большинство уникальных белков, обнаруженных во включениях Httex1 72Q без метки или с меткой GFP, были связаны с ядерными функциями, убиквитин-протеасомной системой, ERAD и путями аутофагии. Более того, среди 13 распространенных белков большинство было связано с механизмом деградации белков (например, убиквилины 1, 2 и 4, а также белки Rad23b и Nedd8).Хотя включения Httex1 72Q и Httex1 72Q-GFP без меток имеют несколько общих белков, связанных с механизмом деградации, более 75% коагрегирующих белков различаются, что может объяснить резкие различия в нейронной токсичности, связанные с каждым типом этих включений.
Поскольку большинство включений, образующихся в нейронах, являются ядерными, мы использовали клетки HEK для дальнейшей оценки того, как метка GFP влияет на протеом цитоплазматических включений Httex1 (дополнительная рис. 11a). Анализ участка вулкана показал 492 белков, значительно обогащенных нерастворимой фракцией клеток НЕК, сверхэкспрессирующих Httex1 72Q-GFP, по сравнению с теми, которые экспрессируют GFP (рис.
Рис. 9: Протеомный анализ Httex1 72Q-GFP во фракции, растворимой в мочевине, выявил 55% различий по сравнению с Httex1 72Q без метки для белков, коагрегированных с Httex1 в клетках НЕК. 9а). Мы наблюдали значительное обогащение эндогенным белком HTT, предполагая, что, как и в случае с Httex1 72Q без метки, мутантные включения Httex1-GFP могут рекрутировать эндогенный белок HTT (дополнительная рис. 11e).a Идентифицированные белки были проиллюстрированы с помощью вулканического графика для сравнения уровней белков, идентифицированных в растворимой в мочевине фракции через 48 часов после трансфекции Httex1 72Q-GFP или GFP в клетках НЕК, в 3 независимых экспериментах. Среднее различие (Log2 (Fold-Change) по оси X) между растворимой в мочевине фракцией клеток HEK, сверхэкспрессирующих Httex1 72Q-GFP или GFP, было построено в зависимости от значимости (Log10 ( p -значение) по оси Y ( Т -Тест)). Для последующих анализов были назначены частота ложных открытий (FDR) 0,05 и порог значимости S0 = 0,5.
b Классификация клеточных компартментов белков, значительно обогащенных растворимой в мочевине фракцией клеток НЕК, сверхэкспрессирующих Httex1 72Q-GFP, по сравнению с клетками, экспрессирующими GFP. Анализы обогащения Gene Ontology (GO) были определены с помощью анализа DAVID (значительно для p значений <0,05 с поправкой на множественное тестирование на основе Бенджамини и Хохберга). c Чтобы сравнить результаты, полученные с Httex1 72Q-GFP, с Httex1 72Q без метки, мы представили белки, обогащенные Httex1 72Q, по сравнению с Httex1 16Q по сравнению с белками, обогащенными Httex1 72Q-GFP, по сравнению с Httex1 16Q, используя диаграмму Венна. . Всего было обнаружено сходство 256 белков (44,6%) между двумя состояниями, 198 белков (34,5%) были уникальными для обогащения Httex1 72-GFP, а 120 белков (20,9%) были для обогащения Httex1 72Q. d Киотская энциклопедия генов и геномов (KEGG) Анализ путей коагрегированных белков с Httex1 72Q и Httex1 72Q-GFP. На тепловой карте представлены важные пути (KEGG), обогащенные растворимыми фракциями мочевины, извлеченными из диаграммы Венна ( c ): коагрегированные белки, уникальные для Httex1 72Q, уникальные для Httex1 72Q-GFP и общие для обоих состояний, все по сравнению к управлению Httex1 16Q. Исходные данные предоставляются в виде файла исходных данных.Изображение в полный размер
Компоненты цитоплазмы, обогащенные нерастворимой фракцией Httex1 72Q-GFP, были аналогичны компонентам, обнаруженным с Httex1-72Q без метки, и были частью эндомембранной системы (~ 50%, светло-голубой Рис. 9b) , цитоскелет, околоядерная область, UPS, тела процессинга мРНК и стрессовые гранулы (рис. 9b, темно-синий). Однако, в отличие от нерастворимых фракций Httex1 72Q без меток, в которых не было обнаружено обогащения митохондриальным белком, 10% белков, обогащенных нерастворимой фракцией Httex1 72Q-GFP, были связаны с компартментом митохондрий (фиолетовый рисунок 9).б). Ядерные белки, обнаруженные в нерастворимой фракции Httex1 72Q-GFP, принадлежат к тем же ядерным компартментам, что и идентифицированные ранее в нерастворимой фракции Httex1 72Q (рис. 9b, серый).
Анализы биологического процесса (дополнительная рис. 26) и канонического пути (дополнительная рис. 27) показали, что UPS и механизм шаперонов были наиболее обогащенными терминами, как ранее наблюдалось в нерастворимой фракции без меток.
Это указывает на то, что процесс агрегации самого Httex1, независимо от присутствия метки GFP, приводит к секвестрации ключевых клеточных механизмов, ответственных за фолдинг и деградацию белков. Это может поставить под угрозу способность клетки предотвращать накопление или удаление агрегатов Httex1. Хотя и в разных пропорциях, большинство биологических процессов и путей, значительно обогащенных нерастворимой фракцией Httex1 72Q, таких как воспаление, транскрипция, передача сигналов HD и гибель клеток, также были обнаружены в нерастворимой фракции Httex1 72Q-GFP.Идентифицировав белки, значительно обогащенные включениями 72Q как без метки, так и с меткой GFP, мы затем определили, какие белки были уникальными для каждого типа включения. С этой целью мы использовали диаграмму Венна для сравнения списков белков, значительно обогащенных в нашем анализе участков вулкана [(Httex1 72Q по сравнению с нерастворимыми фракциями Httex1 16Q) по сравнению с (Httex1 72Q-GFP по сравнению с нерастворимыми фракциями Httex1 16Q)] (рис.
. 3a и S11c). На рисунке 9c показано, что ~ 45% (256 белков) были обнаружены как во включениях Httex1 72Q без метки, так и во включениях с меткой GFP. Всего 120 белков (20,9%) были уникальными для нерастворимой фракции Httex1 72Q, а 198 белков (34,5%) были уникальными для Httex1 72Q-GFP. В целом, мы обнаружили 55% различных белков среди белков, которые коагрегируются или секвестрируются во включениях Httex1 72Q по сравнению с включениями 72Q-GFP, оба по сравнению с Httex1 16Q. Мы использовали анализ путей Киотской энциклопедии генов и геномов (KEGG) для классификации списка белков во включениях Httex1 (рис. 9d). Протеасома, процессинг белков в эндоплазматическом ретикулуме и эндоцитоз были одними из наиболее обогащенных путей общих коагрегированных белков. Эти 3 термина также были значительно обогащены для Httex1 72Q-GFP, что указывает на участие других уникальных белков, обогащенных этими путями по сравнению с Httex1 72Q. Классификация KEGG выявила два отдельных кластера для Httex1 72Q и Httex1 72Q-GFP, а также дополнительные различия (рис. 9).г). Было обнаружено, что белки, связанные с инфекциями и путями воспаления (например, инфекция простого герпеса, передача сигналов TNF и передача сигналов Toll-подобных рецепторов), являются уникальными для Httex1 72Q, тогда как белки, связанные с метаболизмом, были специально обогащены для Httex1 72Q-GFP. Наш протеомный анализ подчеркивает, что добавление флуоресцентной метки, такой как GFP, значительно изменяет не только механизм образования включений Htt, но также интерактом Htt и, следовательно, биохимический состав включений.Включения Httex1 72Q вызывают более сильную дыхательную дисфункцию и более сильное снижение числа ERES по сравнению с Httex1 72Q-GFP в клетках HEK -связанные функции. Во-первых, мы измерили митохондриальное дыхание в различных состояниях дыхания в клетках НЕК (дополнительная рис. 13c, d). По сравнению с нашими выводами с мутантом Httex1 72Q без меток (рис. 4c), усиление митохондриального дыхания было значительным, но менее выраженным для Httex1 72Q-GFP, чем для Httex1 16Q-GFP (дополнительный рисунок 13e).
Учитывая роль митохондрий как основного источника активных форм кислорода (АФК), с последствиями АФК как при нейродегенеративных заболеваниях, так и при клеточных защитных сигнальных каскадах 111,112 , мы подозревали, что на продукцию митохондрий АФК может влиять митохондриальная фрагментация, наблюдаемая в областях, близких к свободному от метки Httex1 72Q и отсутствующая для Httex1 72Q-GFP (дополнительная рис. 28a, b). Чтобы проверить эту гипотезу, мы использовали амплексный красный анализ для измерения производства митохондриальных АФК (супероксид и перекись водорода) одновременно с митохондриальным дыханием (дополнительная рис. 13d). Мы не наблюдали существенных различий в продукции митохондриальных АФК между Httex1 72Q или 16Q без метки или с меткой GFP (дополнительная рис. 28c).Фрагментация митохондрий 113 и воспаление 114 являются отличительными чертами Htt-индуцированной нейродегенерации. Недавние исследования сообщили о главной роли гомеостаза митохондриального деления и слияния в HD, а также выявили специфическую фрагментацию митохондрий, индуцированную мутантным Htt, с помощью электронной микроскопии в нейронах STHdh 115,116 .
Интересно, что только сверхэкспрессия Httex1 72Q без метки приводит к фрагментации митохондрий и характеризуется сильным накоплением белков, связанных с воспалением, в агрегатах. Мы предполагаем, что наблюдаемая гиперактивация митохондриального дыхания (в отсутствие значительного увеличения продукции АФК) является адаптацией клеток, несущих включения, для выработки достаточного количества энергии за счет окислительного фосфорилирования для увеличения потребности в энергии для развертывания или очистки агрегирующих белков. . Аналогичная гиперактивация митохондриального дыхания недавно была обнаружена для агрегатов альфа-синуклеина 117 . Эти авторы предположили, что гиперреспирация может представлять собой патогенное событие, предшествующее патологии альфа-синуклеина. Мы думаем, что это также может быть в случае патологии, индуцированной немеченым Httex1 72Q. Однако, основываясь на нашем обширном протеомном анализе включений, образованных Httex1 72Q или Httex1 72Q-GFP, и сравнивая функциональность органелл в нашей клеточной модели, мы можем предложить более сложную гипотезу. В состоянии без метки Httex1 72Q митохондриальная гиперактивность совпадала как с митохондриальной фрагментацией, так и с обнаружением большего количества белков, связанных с воспалением, в агрегатах по сравнению с состоянием GFP. Мы предполагаем, что митохондриальная фрагментация изначально может быть защитной, поскольку она увеличивает митохондриальную подвижность, необходимую для процессов восстановления клеток 118 . Повышение регуляции митохондриального дыхания может быть разумным следствием повышенных потребностей в АТФ для митохондриального и белкового транспорта, связанного с формированием агрегации и протеостатическими процессами, включая АТФ-требующие шапероновые, протеасомные и аутофагические процессы.Наконец, мы также исследовали, мешают ли включения Httex1-GFP гомеостазу ERES, сравнив размер и количество ERES в клетках HeLa, сверхэкспрессирующих Httex1 16Q-GFP, с клетками, экспрессирующими мутанты Httex1, меченные GFP 39Q или 72Q (дополнительный рис. 29a).
Мы обнаружили, что клетки, содержащие включения Httex1 72Q-GFP, вызывали снижение числа ERES (дополнительная рис. 29b), хотя и незначительное, по сравнению с Httex1 72Q, которое показало снижение на 20% (рис. 4e). Уменьшение размера ERES было значительным для клеток, экспрессирующих Httex1 72Q-GFP, но не Httex1 39Q-GFP, по сравнению с Httex1 16Q-GFP (дополнительная фигура 29c). Таким образом, влияние включений Httex1-GFP на ERES присутствует, но менее выражено по сравнению с наблюдаемым в клетках, содержащих включения Httex1 без метки. Эти наблюдения согласуются с недавними исследованиями крио-ЭТ, предполагающими снижение динамики ЭР вблизи включений Httex1-GFP 12 .В целом, наши результаты показали, что ответы клеточных органелл и адаптация к образованию включений отличались для включений Httex1 72Q-GFP, чем для включений Httex1 72Q, что согласуется с GFP-зависимыми изменениями, наблюдаемыми в протеоме, и ультраструктурными свойствами Httex1 цитоплазматические включения.
Обсуждение
Показано, что патологические включения при нейродегенеративных заболеваниях имеют сложную организацию и состав. Например, недавно было продемонстрировано, что тельца Леви (ТЛ), выделенные из головного мозга больных болезнью Паркинсона, или ЛТ-подобные включения в первичных нейронах состоят не только из нитевидных и агрегированных форм альфа-синуклеина, но и из сложной среды липидов, белков цитоскелета , и другие белки и мембранные органеллы, включая митохондрии и аутофагосомы 119 120 121 122 . Исследования на культурах нейронов также показали, что рекрутирование липидов и мембранных органелл во время образования и созревания LB способствует дисфункциям органелл и приводит к синаптической дисфункции и нейродегенерации. В соответствии с этими выводами наши данные ЭМ вместе с 3D-реконструкциями выявили присутствие фрагментов мембран и везикул, захваченных ядром включений Httex1. В соответствии с этими наблюдениями наш протеомный анализ показал, что 24% белков, обогащенных фракцией включений, принадлежат эндолизосомным компартментам, аппарату Гольджи и транс-сети Гольджи (рис.
3). Кроме того, митохондрии и ЭР были обнаружены на периферии включений, как сообщалось ранее на клеточных моделях 15 и ткани человека 95 123 . Мы предположили, что секвестрация ключевых функциональных белков вместе с липидами, эндомембранами и органеллами внутри включений Httex1 может вызвать нарушение клеточного гомеостаза. В соответствии с этой гипотезой наши электронные микрофотографии показали, что митохондрии, связанные с включениями Httex1 72Q, были фрагментированы и часто имели дезорганизованные или обедненные кристы (рис. 4а). Эти изменения в митохондриальной морфологии были связаны с нарушением регуляции митохондриального дыхания (рис. 4c), что согласуется с предыдущими исследованиями, демонстрирующими, что мутантные агрегаты Htt взаимодействуют непосредственно с наружными митохондриальными мембранами (в клетках STHdh) 124 и вызывают фрагментацию митохондрий (в первичных нейронах) 125 . Дефекты митохондриального дыхания также наблюдались в головном мозге пациентов с БХ, особенно дефекты комплексов II и IV дыхательной цепи могут нарушать окислительное фосфорилирование 126,127,128 .ЭР на периферии включений Httex1 72Q также были затронуты на структурном уровне, о чем свидетельствует их морфологическая реорганизация в розетки или «стопочные цистерны», и на функциональном уровне, о чем свидетельствует нарушение регуляции гомеостаза ERES ( Рис. 4г-е). Формирование розеток ЭР указывает на накопление белков в гладком ЭР 9.1441 92 и, как полагают, является результатом дефектов низкоаффинного связывания и экспорта, которые могут быть вызваны развернутыми белками Htt, но не обязательно связаны со стрессом ER. Kegel и коллеги 129 также сообщили о присутствии цистерн ER рядом с агрегатами, образованными экспрессией большого фрагмента Htt, слитого с меткой FLAG в клональных клетках полосатого тела. Формирование включений Httex1, по-видимому, приводит к реорганизации сети ER на их периферии, но не приводит к секвестрации белков ER, как показывает количественный протеомный анализ, который не выявил значительных белков ER, захваченных внутри включений.
Наличие рибосом и деформация мембраны ЭР ранее были обнаружены вблизи периферии включений Httex1 с помощью крио-ЭТ 9.1441 12 и было связано с сильным снижением динамики ЭР. В том же исследовании 12 периферия включений была иммунореактивна к нескольким компонентам механизма ER-ассоциированной деградации (ERAD) (например, Erlin-2, Sel1L). Эти наблюдения согласуются с нашими данными ICC, показывающими присутствие шаперона ER BIP на внешней периферии, но не внутри включений (дополнительная рис. 5). Мы также наблюдали модуляцию ERES, которую можно объяснить секвестрацией специфических белков, необходимых для их слияния 130 . В соответствии с этими наблюдениями наш протеомный анализ показал обогащение белками, специфичными для переноса ER-Golgi. Модуляция ERES также может быть объяснена снижением биосинтетической способности этого компартмента, т.к. ERES, как было показано, адаптируются к количеству секреторной нагрузки, которой они подвергаются 131 . Рекрутирование и возмущение сети ER может способствовать цитотоксичности во время образования включения, но само по себе, по-видимому, недостаточно, чтобы вызвать явную токсичность. В соответствии с этой гипотезой, ERAD и Ca 2+ сигнальные пути, как известно, не регулируются в различных клеточных и животных моделях с использованием различных фрагментов Htt 132,133,134,135,136,137 . В целом, мы показали, что образование и созревание цитоплазматических включений Httex1 включает сложное взаимодействие между агрегатами Httex1 и органеллами.Хотя предыдущие исследования предполагали образование кольцеобразных ядерных включений, эта архитектура определялась периферическим окрашиванием ядерных белков, а не периферическим окрашиванием Htt, как это наблюдается в случае цитоплазматических включений (рис. 1). Наша глубокая характеристика включений Httex1 72Q на уровне ультраструктуры выявила отчетливые различия в организации и архитектуре между цитоплазматическими и ядерными включениями Htt.
Цитоплазматические включения Httex1 72Q демонстрируют плотную морфологию ядра и оболочки и состоят из высокоорганизованных фибриллярных агрегатов как в ядре, так и на периферии. Напротив, ядерные включения Htt как в клетках HEK, так и в первичных нейронах коры проявляли менее сложные ультраструктурные свойства, выглядели как гомогенные структуры без морфологии ядра и периферии и были лишены эндомембран или везикул (рис. 2 и 6a). Эти наблюдения согласуются с большинством ядерных включений Htt, обнаруженных в моделях HD мышей, которые не отображают организацию ядра и оболочки 19,32 .Наши новые данные о включении Htt в ядерные нейроны показали, что экспрессия мутанта Httex1 без метки в первичных нейронах приводит к образованию грануло-филаментных ядерных включений, подобно предыдущим ультраструктурным исследованиям, проведенным в мозге пациентов с БХ и в моделях БХ in vivo, включая Мыши R6/2 19,32 и трансгенные крысы 138 . Однако в большинстве этих исследований ультраструктурные свойства филаментов были менее очевидны по сравнению с хорошо разрешенной филаментной структурой, которую мы и другие 12 показали с помощью электронной томографии.
Например, Тагава и его коллеги также наблюдали ядерные включения Httex1 без меток, но их данные не выявили того уровня ультраструктурных деталей и понимания, который дает наше исследование 139 .Наши результаты также предполагают, что внутриклеточная среда и взаимодействия с липидами и эндомембранами или везикулами являются ключевыми детерминантами структурной и молекулярной сложности включений и что различные механизмы управляют образованием и созреванием ядерных и цитоплазматических включений. Такие дифференциальные характеристики между цитоплазматическими и ядерными включениями могут объяснить дифференциальную клеточную дисфункцию и токсичность, связанные с этими двумя типами включений Htt 29 140 141 142 143 . Кроме того, дифференциальная токсичность, наблюдаемая для ядерных включений Htt в первичных нейронах по сравнению с клетками HEK 293, указывает на токсичность Htt, зависящую от типа клеток, которая может возникать из-за различных механизмов агрегации Htt или взаимодействия с клеточными белками или различий в устойчивости различных клеток на основе например, на отчетливую толерантность к окислительному стрессу и иммунные функции/воспаление.
Мы предполагаем, что формирование ядра включений управляется преимущественно межмолекулярными взаимодействиями с участием домена polyQ и может быть инициировано быстрыми событиями, потенциально вызванными фазовым разделением, как недавно описано несколькими группами 14,39,144,145 . Быстрое формирование ядра агрегата не позволяет регулировать рекрутирование других белков и органелл (рис. 1, 2 и 10а), о чем свидетельствует тот факт, что большинство антител против белков, обнаруженных во включении Htt, окрашивают периферию, а не ядро включений 48 . Домен Nt17, скорее всего, играет ключевую роль в начальных событиях олигомеризации и, возможно, в упаковке или латеральной ассоциации фибрилл. Как только это ядро плотных фибрилл сформировано, фибриллы на периферии продолжают расти за счет рекрутирования эндогенных растворимых белков (рис. 3 и 10а). Поскольку эта фаза роста более медленная, она позволяет фибриллам взаимодействовать и/или привлекать другие белки во включения.
Рис. 10: Формирование включений Хантингтина обусловлено главным образом повторным доменом полиQ и включает активное рекрутирование и секвестрацию липидов, белков и мембранных органелл. В соответствии с этой гипотезой Мацумото и его коллеги показали, что транскрипционные факторы TATA-связывающий белок (TBP) и CREB-связывающий белок (CBP), содержащие экспансию polyQ, присутствовали только на периферии включений, а не в сердцевине 9.1441 47 . Интересно, что хотя длина повтора polyQ, по-видимому, не оказывала существенного влияния на плотность фибрилл в сердцевине включений, периферическая организация фибрилл Htt и формирование внешней оболочки показали сильную зависимость от длины повтора polyQ (рис. 4 и 10а). Эта модель подтверждается исследованиями Hazeki et al. и Kim et al., продемонстрировав, что нерастворимое в детергенте ядро клеточных включений Httex1 представляет собой скелетную структуру включений, в то время как активная поверхность динамически взаимодействует с видами Htt и процессируемыми белками 30,38 . В целом, наши результаты показывают, что образование включений Htt происходит в две основные фазы: во-первых, формирование ядра, обусловленное главным образом повторным доменом polyQ, а затем рост включений с добавлением фибрилл Htt и рекрутирование другие белки и органеллы. Вторая фаза, по-видимому, обусловлена взаимодействиями, включающими как домены polyQ, так и домены PRD, и включает активное привлечение и секвестрацию липидов, белков и мембранных органелл (рис. 10a). Однако мы не можем исключить присутствие захваченных олигомеров в ядре или на поверхности вблизи растущих фибрилл на периферии 146 . Ранее Qin et al., используя FLAG-меченый фрагмент Htt1-969 (100Q), продемонстрировали, что виды Htt в сердцевине включений обладают высокой устойчивостью к протеазе и что олигомерные формы Htt накапливаются с цитоплазматическими белками и везикулами в периферия включений 48 . В нашем исследовании мы продемонстрировали, что цитоплазматические и ядерные включения Htt обладают различными ультраструктурными свойствами, на которые по-разному влияет размер повторов полиQ. a Мы разделили формирование и организацию цитоплазматических включений Httex1 на различные элементы: (1) хотя домен Nt17 может играть главную роль на ранних стадиях олигомеризации, мы показали, что фибрилизация управляется повторами polyQ; (2) Ранее было показано, что инициирование образования включений происходит по механизму фазового перехода 14,31 и может включать в себя секвестрацию белков и близлежащих органелл прямо или косвенно; (3) Во время роста включения многие клеточные белки, эндомембраны и липиды рекрутируются и секвестрируются внутри цитоплазматических включений; (4) Зрелые цитоплазматические включения Httex1, образующиеся в клетках, демонстрируют структурную организацию ядра и оболочки, если длина полиQ относительно велика (72Q), но не близка к патогенному порогу (39Q), даже если включения все еще формируются во второй кейс. Образование цитоплазматических включений приводит к накоплению митохондрий и сети ЭР на периферии; (5) образование включения Httex1 приводит к адаптации ER-митохондриальной сети и дыхательной дисфункции, а также значительной токсичности при длительной инкубации; (6) Ядерные включения не имеют отчетливой организации ядра/оболочки, аналогичной цитоплазматическим включениям, но они все еще обнаруживаются в виде кольца, поскольку они непроницаемы для антител.
Включения Httex1 без ядерной метки также обогащены нейтральными липидами. b Изображение показывает различную структурную организацию и клеточное влияние Httex1 72Q (слева) по сравнению с Httex1 72Q-GFP (справа). Расположение и упаковка фибрилл Httex1 различны в зависимости от присутствия GFP, а также взаимодействия, рекрутирования и возмущений с окружающими органеллами.Изображение в полный размер
Насколько нам известно, это первое исследование, в котором непосредственно сравниваются включения, образованные мутантными нативными белками Httex1 без меток, с мутантным Httex1, меченным GFP, на структурном, интерактомном и биохимическом уровнях.
Ультраструктура включений Httex1 72Q-GFP выявила лучистые фибриллы с однородным окрашиванием и отсутствием морфологии ядра и оболочки. В соответствии с нашими выводами предыдущие исследования показали, что цитоплазматические агрегаты Httex1 72Q-GFP являются фибриллярными 14,147 и напоминают другие агрегаты Httex1-GFP в клетках COS 148 , в первичных полосатых нейронах 149 , а также агрегаты, образованные мутантным Httex1 объединен с тегом FLAG в ячейках HEK 15 .
В недавнем исследовании криоэлектронной томографии Bäuerlein et al. исследовали ультраструктурные свойства мутантного Httex1 в отсутствие и в присутствии GFP и предположили, что присутствие GFP существенно не изменяет организацию цитоплазматических включений в HeLa и нейронах 12 . Подобно нашим выводам, они показали, что присутствие метки GFP 1) вызывает снижение плотности фибрилл на 50% и увеличение жесткости фибрилл на 25%; 2) приводит к увеличению расстояния между фибриллами во включениях Httex1-GFP; а также; 3) не влияет на общий размер включений. Однако, когда дело доходит до ультраструктурных свойств включений мутанта Httex1 без метки, мы наблюдаем очень разные результаты, поскольку они не наблюдали образования включений с морфологией ядра и оболочки. Изучение последовательности используемых ими конструкций выявило Myc-метку на N-концевой стороне их белков Httex1, тогда как используемые нами конструкции содержали только нативную последовательность мутантного Httex1. Хотя мы показали, что удаление домена Nt17 не влияет на способность Htt без метки образовывать включения с организацией ядра и оболочки, слияние дополнительных аминокислот на N-конце все еще может значительно изменить кинетику, олигомеризацию и путь агрегации белка Httex1. В дополнение к различиям на ультраструктурном уровне наши исследования показали рекрутирование мембранных органелл и везикул в виде нарушенных структур внутри включений как в ядре, так и в оболочке, а также в виде интактной ЭР-митохондриальной сети на периферии бесметочного включения. Менее интернализованные структуры наблюдались во включениях Httex1-GFP, что согласуется с выводами Bäuerlein et al. 12 . Еще одно важное различие между двумя исследованиями заключается в том, что Bäuerlein et al. не наблюдали никакого влияния длины полиQ на размер или ультраструктурные свойства включений, образованных мутантным Httex1-GFP в диапазоне повторов полиQ от 64Q до 97Q. Хотя присутствие GFP может объяснить различия между двумя результатами, также возможно, что эти различия связаны с тем, что авторы сравнивали повторы полиQ, которые далеко за пределами патогенного порога. Мы считаем, что в этом диапазоне polyQ разница в агрегационных свойствах между 64 и 97Q будет небольшим по сравнению с тем, что можно было бы наблюдать при сравнении мутантного Httex1 с повторами polyQ, близкими к патогенному порогу (39-43Q). Поэтому в нашем исследовании мы сравнили ультраструктурные свойства включений Httex1, содержащих 39 или 72Q повторов. В наших экспериментах цитоплазматические включения, образованные мутантным Httex1-39Q, оказались менее организованными и не имели стабильного расположения ядра и оболочки, как в случае Httex1-72Q. Наконец, в первичных нейронах использование Httex1 72Q-GFP резко изменило кинетику образования включений и долю малых и крупных ядерных агрегатов по сравнению с Httex1 72Q без метки. Более того, нейрональные ядерные включения Httex1 72Q-GFP были меньше и менее плотными по сравнению с включениями Httex1 72Q без метки, демонстрировали отличный состав протеома и меньше взаимодействовали с ядерной мембраной. Эти различия могут объяснить пониженную токсичность включений Httex1 72Q-GFP по сравнению с включениями Httex1 72Q без метки.В целом наши данные свидетельствуют о том, что процессы агрегации Httex1 без метки и с меткой GFP различаются (рис. 10b и S30), что подчеркивает потенциальные ограничения использования GFP для исследования молекулярных, биохимических и клеточных детерминант Htt.
образование включений и механизмы токсичности.Таким образом, наши интегративные визуализирующие и протеомные исследования демонстрируют, что процесс образования цитоплазматических включений Htt происходит по крайней мере в две фазы и включает активное рекрутирование липидов, белков и органелл. На организацию и ультраструктурные свойства этих включений большое влияние оказывает длина повтора polyQ и внутриклеточное окружение, о чем свидетельствует тот факт, что цитоплазматические и ядерные включения Httex1 проявляют различные ультраструктурные свойства. Не менее важно, что наша работа подчеркивает важность выяснения роли взаимодействий Htt с липидами и мембранными органеллами в регуляции процесса агрегации Htt, образования включений и токсичности. Распространение представленных здесь исследований и подходов на другие модельные системы HD и другие фрагменты Htt проложит путь к лучшему пониманию механизмов образования включений Htt и того, как они способствуют развитию HD. Недавно мы продемонстрировали, что более длинные N-концевые фрагменты Htt, такие как Htt171, агрегируют in vitro с помощью механизмов, отличных от наблюдаемых для мутантного Httex1, где структурированные домены вне экзона1 могут играть важную роль в регуляции ранних событий, связанных с олигомеризацией Htt, инициацией агрегации Htt.
, и образование включений 150 .В совокупности наша работа дает важные и новые идеи, которые не только улучшают наше понимание механизмов агрегации Htt, но и указывают на новые направления терапевтических вмешательств. Показано, что агрегация Htt и образование включений в цитозоле и ядре происходят по разным механизмам и приводят к образованию включений с различными биохимическими и ультраструктурными свойствами. Эти наблюдения предполагают, что два типа включений могут проявлять свою токсичность через разные механизмы и могут требовать разных стратегий, чтобы препятствовать их образованию, созреванию и токсичности. Кроме того, наша работа предполагает, что идентификация модификаторов роста включений Htt и аберрантных вторичных взаимодействий с другими белками и органеллами представляет собой альтернативу и стратегию для вмешательства в токсичность, вызванную Htt, и замедления прогрессирования заболевания, особенно после начала заболевания. Поэтому мы считаем, что нацеливание на рост и созревание включений представляет собой жизнеспособную терапевтическую стратегию.
Методы
ДНК-конструкции и очистка
Вектор экспрессии pCMV млекопитающих, кодирующий Httex1 16Q, Httex1 16Q-GFP, Httex1 39Q, Httex1 39Q-GFP, Httex1 72Q и Httex1 IR 72Q-GFP, были любезно предоставлены Andrea Caricasole (AndreaBM) . ΔN17-Httex1 39Q, ΔN17-Httex1 39Q-GFP, ΔN17-Httex1 72Q и ΔN17-Httex1 72Q-GFP были приобретены у GeneArt (Германия). Был приобретен вектор экспрессии SIN-PGK, кодирующий Httex1 16Q, Httex1 16Q-GFP, Httex1 72Q и Httex1 72Q-GFP, ΔN17-Httex1 16Q, ΔN17-Httex1 16Q-GFP, ΔN17-Httex1 72Q и ΔN17-Httex1 72Q-GFP. от GeneArt (Германия). Плазмиды трансформировали в химиокомпетентные E. coli стабильные 3 клетки (Stbl3) от Life Technologies (Швейцария) и очистку плазмиды Maxiprep (Life Technologies, Швейцария) проводили в соответствии с инструкциями производителя.
Производство лентивируса
В день 0 клетки HEK 293 ([HEK-293] ATCC ® CRL-1573’“) высевали на семь чашек диаметром 15 см. Через 24 ч после посева клетки трансфицировали интересующей плазмидой (160 мкг векторной плазмиды SIN-PGK), плазмидой, кодирующей белок оболочки G VSV (55,3 мкг pMDG2G), и плазмидой, кодирующей gag, Pol, TAT и Rev.
белков (102,2 мкг pCMVR8.74) с использованием набора для трансфекции фосфата кальция (Takara, Япония). Среду заменяли свежей через 24 часа после трансфекции, а затем собирали и заменяли через 8 часов и 24 часа. Собранные среды центрифугировали при 500 g при 4 °C в течение 5 минут для осаждения клеточного дебриса. Супернатант очищали фильтром 0,22 мкм, а затем центрифугировали при 50 000 g при 16 °C в течение 2 часов. Супернатант отбрасывали, а осадок сушили на воздухе путем переворачивания. Наконец, осадок хорошо ресуспендировали в PBS и хранили при -80 °C. Этот метод был выполнен в соответствии с протоколом Barde et al. 151 .Культура клеток млекопитающих и плазмидная трансфекция
Клетки HEK 293 ([HEK-293/17] ATCC ® CRL-11268’“) содержали в модифицированной Дульбекко среде Игла DMEM (Life Technologies, Швейцария), содержащей 10% FBS ( Life Technologies, Швейцария), 10 мкг/мл стрептомицина и пенициллина (Life Technologies, Швейцария) во влажном инкубаторе и 5% СО -2- при 37 °С.
Клетки высевали с плотностью 100 000 на чашку в µ-чашках со стеклянным дном (IBIDI) или 50 000 клеток/лунку в 24-луночных планшетах с пластиковым покровным стеклом Thermanox (круглым) диаметром 13 мм (Life Technologies, Швейцария) для того, чтобы получить клетки на уровне 70–90% слияние на следующий день для процедуры трансфекции с использованием стандартной процедуры с фосфатом кальция 152 . Вкратце, 2 мкг ДНК разводили в 30 мкл H 2 O и 30 мкл 0,5 М CaCl2 перед добавлением по каплям 60 мкл 2xHBS, pH 7,2 (50 мМ HEPES, pH 7,05; 10 мМ KCL; 12 мМ декстрозы). ; 280 мМ NaCl; 1,5 мМ Na2PO4, pH 7,2, растворенных в h3O) в условиях легкого встряхивания.Первичная культура клеток коры головного мозга мышей и лентивирусная трансдукция
Сначала кору головного мозга выделяли у детенышей P0 мышей дикого типа (C57BL/6JRccHsd, Harlan) в сбалансированном солевом растворе Хенкса (HBSS) 122 153 . Ткани головного мозга расщепляли папаином (20 ЕД/мл, Sigma-Aldrich, Швейцария) в течение 30 мин при 37°С и останавливали расщепление с помощью ингибитора трипсина (Sigma-Aldrich, Швейцария).
Далее ткани диссоциировали механическим растиранием, а выделенные клетки ресуспендировали в адгезивной среде (МЕМ, 10% лошадиная сыворотка, 30% глюкоза, L-глутамин и пенициллин/стрептомицин) (Life Technologies, Швейцария). Наконец, нейроны высевали в 6-луночные планшеты для биохимического анализа при плотности 500 000 клеток/мл или в 24-луночные планшеты, содержащие покровные стекла при плотности 125 000 клеток/мл, предварительно покрытые поли-1-лизином 0,1. % мас./об. в воде (Sigma-Aldrich). Через 3 часа адгезивную среду удаляли и заменяли нейробазальной средой (Life Technologies, Швейцария) с добавлением L-глютамина, пенициллина/стрептомицина (100 ЕД/мл, Life Technologies, Швейцария) и добавки B27 (Life Technologies, Швейцария). .Первичные нейроны коры трансдуцировали частицами лентивируса Httex1 при множественности заражения (MOI) 2 через 5 или 7 дней in vitro (DIV) 154 . Все процедуры были одобрены Федеральным ветеринарным управлением Швейцарии (номер разрешения VD 3392).
Условия содержания мышей: 22°С, световой день 12–14 ч, относительная влажность от 50 до 60%.Иммуноцитохимия (ICC)
В указанный момент времени клетки HEK 293 или кортикальные первичные нейроны дважды промывали PBS pH 7,4 (1X) (Life Technologies, Швейцария) и фиксировали в 3,7% формальдегиде (Sigma-Aldrich, Швейцария). ) в PBS (PFA) в течение 15 мин при комнатной температуре (КТ). После стадии блокирования 3% BSA (Sigma-Aldrich, Швейцария), разбавленного 0,1% Triton X-100 (Applichem, Германия) в PBS (PBST) в течение 30 минут при комнатной температуре, клетки инкубировали с первичными антителами (дополнительная рис. 1) анти-Htt, индуцированные против домена Nt17 (2B7, CHDI [Cure Huntington’s Disease Initiative]; Ab109115, Abcam), или PolyQ (MW1, CHDI), или домен, богатый пролином (PRD) (MAB5492, Millipore; 4C9, CHDI; N18, Santa-Cruz и EGT 414), или против Htt (S830) в разведении 1 /500 в PBST в течение 2 ч при комнатной температуре. Затем клетки пять раз промывали в PBST и инкубировали в течение 1 часа при комнатной температуре со вторичными антителами осла против мыши Alexa647, ослиными антителами против кролика Alexa647 или ослиными антителами против коз 568 (Life Technologies, Швейцария), используемыми в разведении 1/.
800 в PBST и DAPI (Sigma-Aldrich, Швейцария) в концентрации 2 мкг/мл, все разведены в PBST. Кроме того, клетки НЕК контрастировали фаллоидином Atto59.4 (Sigma-Aldrich, Швейцария), который имеет высокое сродство к филаментозному F-актину, а нейроны коры выявляли с помощью антитела к MAP2.Затем клетки промывали пять раз в PBST и последний раз в бидистиллированной воде, после чего помещали в среду для заливки поливинилового спирта (ПВС) с DABCO (Sigma-Aldrich, Швейцария). Клетки исследовали с помощью конфокального лазерного сканирующего микроскопа (LSM 700, Zeiss, Германия) с масляным объективом 40 × 1,3 (Plan-Apochromat) и анализировали с помощью программного обеспечения Zen (Zeiss, Германия) или с помощью конфокального лазерного сканирующего микроскопа (Inverted Leica SP8, Германия) с масляным объективом 63×/1,4 (HC PL APO) и проанализированы с помощью программного обеспечения LASX Leica.
Количественная оценка на основе изображений Экспрессия Httex1 в первичных нейронах
Для каждого независимого эксперимента, как описано выше, визуализировали как минимум пять областей для каждого состояния.
Каждый эксперимент проводили трижды. Функция счетчика клеток использовалась в программном обеспечении LASX Leica для количественной оценки морфологической экспрессии Httex1 в нейронах в разные моменты времени. Классификация проводилась в соответствии с рис. 5c, при этом обнаружение Httex1 классифицировалось как диффузное; маленькие ядерные точки; крупные ядерные включения или цитоплазматические включения. В каждом состоянии для каждого условия количественно определяли примерно 150 нейронов и проводили как минимум три независимых эксперимента.Иммунофлуоресцентное окрашивание мест выхода ЭР
Всего 100 000 клеток HeLa высевали в 6-луночный планшет на покровные стекла. Через 24 ч клетки трансфицировали различными вариантами меченого или не содержащего GFP Httex1 с использованием Fugene 6 в соответствии с инструкциями производителя. Пустой вектор использовали в качестве отрицательного контроля. Через 48 часов после трансфекции клетки фиксировали в 4% PFA в течение 20 минут и окрашивали с использованием анти-Htt-антитела (Millipore, мышиное моноклональное (MAB5492)).
Вкратце, после промывания клеток PBS, содержащим 20 мМ глицина, предметные стекла инкубировали в блокирующем буфере, состоящем из 3% BSA (бычий сывороточный альбумин) в 0,1% Triton X-100 и PBS в течение 30 минут при комнатной температуре. Затем клетки инкубировали с первичными антителами против Htt и Sec13 (R&D Systems) для маркировки сайтов выхода ER с последующей промывкой и инкубацией с вторичными антителами, мечеными Alexa-Fluor. Предметные стекла промывали PBS и заключали в заливочную среду из поливинилового спирта с DABCO (Sigma-Aldrich). Клетки визуализировали с помощью объектива 63× с использованием конфокального микроскопа Zeiss LSM 700.В случае Httex1-GFP клетки промывали в PBS, содержащем 20 мМ глицина, с последующей пермеабилизацией в PBS, содержащем 0,2% Triton X-100. Затем клетки инкубировали с первичными антителами для окрашивания мест выхода ER, разведенных в 3% BSA в PBS. После промывания PBS клетки инкубировали с соответствующими вторичными антителами, помеченными Alexa-Fluor, в 3% BSA-PBS.
Предметные стекла промывали PBS и заключали в заливочную среду из поливинилового спирта с DABCO (Sigma-Aldrich). Клетки визуализировали с помощью объектива 63× с использованием конфокального микроскопа Zeiss LSM 700.Количественная оценка количества и размера выходных участков ЭР была выполнена с использованием инструмента анализа частиц на изображении J после порогового значения размера и интенсивности пикселей.
Корреляционная световая и электронная микроскопия (CLEM)
Клетки HEK 293 выращивали при концентрации 600 000 клеток/мл на 35-мм чашках с буквенно-цифровыми поисковыми сетками, выгравированными на нижнем стекле (MatTek Corporation, Эшленд, Массачусетс, США). Через 48 ч после трансфекции пустым вектором Httex1 72Q или Httex1 72Q-GFP клетки фиксировали в течение 2 ч 1% глутаральдегидом (Electron Microscopy Sciences, США) и 2,0% PFA в 0,1 М фосфатном буфере (ФБ) при рН 7,4. . Точно так же первичные корковые нейроны, выращенные на стеклянных чашках с сеткой (MatTek Corporation, Ашленд, Массачусетс, США) или вручную аннотированные 13-мм пластиковые покровные стекла [Thermanox 174950] (Thermo Fisher Scientific, Уолтем, США) промывали через 7 дней после трансдукции и фиксировали аналогично клеткам НЕК, описанным выше.
После промывания PBS выполняли ICC, как описано выше. Внутриклеточные включения окрашивали Htt-антителами (Millipore MAB5492, а.о. 1-82) и интересующие клетки визуализировали с помощью флуоресцентного конфокального микроскопа (LSM700, Carl Zeiss Microscopy) с объективом 40x. Точное положение выбранных клеток регистрировали с помощью буквенно-цифровой сетки, выгравированной на дне чашки. Затем клетки дополнительно фиксировали 2,5% глутаральдегидом и 2,0% PFA в 0,1 М PB при pH 7,4 в течение еще 2 часов. После пяти 5-минутных промывок 0,1 М какодилатным буфером с рН 7,4 клетки постфиксировали 1% четырехокисью осмия в том же буфере в течение 1 часа, а затем промывали бидистиллированной водой перед контрастированием 1% уранилацетатной водой в течение 1 часа. 1 ч. Затем клетки обезвоживали в возрастающих концентрациях спирта (2 × 50%, 1 × 70%, 1 × 90%, 1 ×95% и 2 ×100%) в течение 3 мин при каждой промывке. Дегидратированные клетки инфильтрировали смолой Durcupan (Electron Microscopy Sciences, Hatfield, PA, USA), разбавленной абсолютным этанолом в соотношении 1:2 в течение 30 мин, в соотношении 1:1 в течение 30 мин, в соотношении 2:1 в течение 30 мин и дважды в чистом этаноле.
Дуркупана по 30 мин каждый. После 2-часовой инкубации в свежей смоле Durcupan чашки переносили в печь при 65°C, чтобы смола могла полимеризоваться в течение ночи. После затвердевания смолы стеклянный CS на дне чашки удаляли повторным погружением в горячую воду (60 °C), а затем в жидкий азот. Затем определяли интересующую ячейку, используя ранее записанные буквенно-цифровые координаты, и лезвием бритвы вырезали эту область из остальной части смолы. Затем этот кусок был приклеен к полимерному блоку акриловым клеем и обрезан стеклянным ножом с использованием ультрамикротома (Leica Ultracut UCT, Leica Microsystems). Затем ультратонкие срезы (50–60 нм) серийно вырезали с лица алмазным ножом (Diatome, Biel, Швейцария) и собирали на 2 мм однощелевые медные сетки, покрытые пластиковой поддерживающей пленкой формвар. Срезы контрастировали уранилацетатом и цитратом свинца и визуализировали с помощью просвечивающего электронного микроскопа (Tecnai Spirit EM, FEI, Нидерланды), работающего при ускоряющем напряжении 80 кВ и оснащенного цифровой камерой (FEI Eagle, FEI).Обработка образцов для электронной микроскопии без пермеабилизации клеток
Через 48 ч после трансфекции клетки НЕК 293 фиксировали в 2% PFA и 2% глутаральдегиде в фосфатном буфере 0,1 М (pH 7,4) в течение 1 ч и 30 мин. Для сохранения внутренних мембран клеток ИКК не проводили. Затем клетки промывали 3 раза по 5 мин в какодилатном буфере (0,1 М, рН 7,4). Затем они были постфиксированы 1% четырехокисью осмия плюс 1,5% ферроцианида калия в какодилатном буфере (0,1 М, рН 7,4) при комнатной температуре в течение 40 мин с последующей постфиксацией 1% четырехокисью осмия в какодилатном буфере (0,1 М, рН 7,4) при комнатной температуре в течение 40 мин. Образцы промывали дважды по 5 мин в дистиллированной воде, затем окрашивали 1% раствором уранилацетата в воде в течение 40 мин и один раз промывали в бидистиллированной воде в течение 5 мин. Образцы обезвоживали в возрастающих концентрациях этанола в течение 3 минут при каждой промывке (2 × 50%, 1 × 70%, 1 × 90%, 1 ×95%, 2 × 100%), а затем заливали эпоксидной смолой (формула Epon: Embed 812: 20 г, DDSA: 6,1 г, NMA: 13,8 г, DMP 30: 0,6 г) (Электронная микроскопия наук, США) за счет непрерывного вращения флаконов.
Процесс заливки начинается со смеси этанола и эпона 1:1 в течение 30 минут, после чего следует 100% EPON в течение 1 часа. Затем EPON заменяли свежим EPON на 2 часа. Наконец, образцы помещали между предметными стеклами с покрытием и помещали в печь при 65 °C на ночь. Серийные срезы толщиной 50 нм вырезали с помощью ультрамикротома (UC7, Leica Microsystems, Германия) и собирали на пленках формваровой подложки на медных сетках с одной щелью (Electron Microscopy Sciences, США) для визуализации в просвечивающей электронной микроскопии (ПЭМ). Изображения ПЭМ были получены при натяжении нити накала 80 кВ с помощью ЭМ-микроскопа Tecnai Spirit с использованием камеры Eagle 4 k × 4 k. Не менее 8 клеток визуализировали в каждом состоянии при 29Увеличение 00× и 4800×.Изображения были выровнены с использованием программного обеспечения Photoshop CC 2019 (Adobe, США), и различные органеллы (ядра, митохондрии, эндоплазматический ретикулум, включения Httex1) были сначала сегментированы вручную с использованием функции ареалист в плагине trackEM2 в программном обеспечении FIJI.
Затем мы перешли к специально разработанному конвейеру на основе машинного обучения, специально адаптированному для данных трехмерной микроскопии (www.ariadne-service.ch) после проверки с использованием ручной сегментации для справки. В конце процесса сегментации различные сегментированные области были экспортированы в виде масок последовательного изображения, затем визуализированы как объекты в плагине 3D-просмотра в FIJI и экспортированы в виде волнового фронта в Blender 9.1441 ® Программное обеспечение для 3D-моделирования (Blender Foundation). С помощью Blender® сначала были скорректированы 3D-оси в соответствии с ориентацией модели, а также была скорректирована шкала Z. Включение Httex1, митохондрии и ядро были сглажены, а включение Httex1 было скорректировано для визуализации внутриагрегатных мембранных структур внутри включения. Дополнительные измерения проводили по электронным микрофотографиям в клетках, содержащих включения Httex1, с использованием FIJI. Бинарное изображение (вставка) рис. 1d было создано из электронной микрофотографии с использованием медианной фильтрации и порога интенсивности Оцу, что позволяет лучше различать ультраструктуру ядра и оболочки включения Httex1 72Q без метки. Длина митохондриального профиля соответствует максимальной длине каждой митохондрии в одной плоскости ЭМ. Расстояние от включения не учитывалось, так как измерения проводились в одной плоскости. Вместо этого была принята во внимание средняя длина всех обнаруженных митохондрий, которая показала значительные различия в состоянии Httex1 72Q по сравнению с контролем EV.Замораживание и заливка под высоким давлением для ПЭМ
Через 48 ч после трансфекции клетки HEK 293 культивировали на сапфировых дисках (диаметром 6 мм) и быстро замораживали под высоким давлением (HPM100, Leica Microsystems). Диски помещали в криопробирки, содержащие ацетон с 1 % четырехокиси осмия, 0,5 % уранилацетата и 5 % воды при -90 °С. Затем их оставляли при этой температуре на 24 часа, а затем нагревали до 0°C в течение следующих 72 часов, после чего их промывали чистым ацетоном, а затем пропитывали увеличивающейся концентрацией смолы Epon.
После погружения в 100% смолу образцы оставляли при комнатной температуре на 24 часа, а затем смола затвердевала в печи при 65°С в течение 48 часов. Серийные срезы собирали на однощелевые медные сетки с формваровой подложкой, а затем окрашивали цитратом свинца и уранилацетатом. Их визуализировали в просвечивающем электронном микроскопе при 80 кВ (Tecnai Spirit, FEI Company) с использованием CCD-камеры (Eagle, FEI Company).Электронная томография (ET)
Шесть серий наклонов были собраны с помощью программного обеспечения Tomo 4.0 (Thermo Fisher Scientific) на Tecnai F20 TEM, работающем на 200 kV (Thermo Fisher Scientific), на камере Falcon III DD (Thermo Fisher Scientific) в линейном режиме при увеличении 29 000×. Серия наклонов регистрировалась от -58° до 58° по схеме непрерывного наклона с шагом 2°. Совмещение серии наклонов и реконструкция томограмм были выполнены с помощью Inspect3D v4.1.2 (Thermo Fisher Scientific) с использованием 22 итераций SIRT.
Полученная томограмма была подвергнута заполнению недостающего клина новым инструментом на основе глубокого обучения в сборке EMAN2 после 20.
03.2020 155 . Томограммы с отсутствующим клином были подвергнуты полуавтоматической аннотации томограмм на основе полуавтоматической сверточной нейронной сети (CNN) без шаблона в EMAN2. Шесть томограмм были импортированы в менеджер проекта EMAN2 и уменьшены в два раза. Затем томограммы проверялись срез за срезом и аннотировались путем ручного выбора нескольких плиток размером 64 × 64 пикселей, содержащих удлиненные элементы, напоминающие фибриллы, для обучения CNN. Кроме того, несколько участков томограммы, не содержащих удлиненных элементов, были вручную аннотированы как отрицательные примеры. Отобранные положительные примеры были вручную сегментированы с точностью до пикселя. Для обучения нейронной сети были предоставлены как положительные, так и отрицательные примеры. Затем обученный CNN был применен к исходной томограмме для полной аннотации. Изображения и видеоролики были сгенерированы с помощью 3dmod из пакета IMOD 9.1441 156 157 .Подготовка образцов белка для биохимического анализа
Образцы были получены в двух экземплярах в ходе эксперимента HRR для анализа митохондриальных маркеров с помощью WB и FT.
Трансфицированные клетки лизировали в 75 мкл буфера RIPA (150 мкл NaCl, 1 мк NP40, 0,5% дезоксихолата, 0,1% додецилсульфата натрия (ДСН), 50 мкл трис, pH 8). Клеточные лизаты инкубировали при 4°С в течение 20 мин, а затем очищали центрифугированием при 4°С в течение 20 мин при 16 000 g. Супернатанты собирали в виде растворимых белковых фракций и хранили при -20°С после добавления LB5x и 5-минутного кипячения. BCA выполняли на растворимой фракции RIPA. Осадки промывали 500 мкл PBS, затем снова центрифугировали в течение 5 мин при 16 000 g. Супернатанты удаляли, а осадок ресуспендировали в 30 мкл PBS с добавлением 2% SDS и обрабатывали ультразвуком тонким зондом [3 раза, 3 с при амплитуде 60% (Sonic Vibra Cell, Blanc Labo, Швейцария)]. Мембрану из ацетата целлюлозы сначала уравновешивали 2% SDS (в PBS) в течение 5 минут, а основную складку фильтра помещали поверх 2 листов бумаги Watman внутри аппарата Bio-Dot (№ 1706545, США). Образцы загружали и фильтровали под вакуумом. Мембрану промывали 3 раза 0,5% SDS в PBS и применяли вакуум. Затем ацетатную мембрану удаляли и один раз промывали 1% PBS-Tween. Мембраны WB и Filter-Trap блокировали в течение ночи блокирующим буфером Odyssey (LiCor), а затем инкубировали при комнатной температуре в течение 2 часов с различными первичными антителами (анти-хантингтин MAB549).2 и анти-VDAC1), разведенных в том же блокирующем буфере (1/5000). Мембраны промывали 1% PBS-Tween, а затем инкубировали со вторичными антителами, разведенными в блокирующем буфере (1/5000), в течение 1 ч при комнатной температуре перед окончательной промывкой в 1% PBS-Tween. Обнаружение белка проводили по флуоресценции с использованием Odyssey CLx от LiCor. Интенсивность сигнала количественно определяли с помощью Image Studio 3.1 от LiCor.Подготовка образцов для масс-спектрометрии
Образцы были созданы в трех экземплярах для количественного масс-спектрометрического анализа. В указанный момент времени HEK-клетки или первичные нейроны лизировали в PBS с добавлением 0,5% NP40, 0,5% Triton x100, 1% ингибитора протеазного коктейля (Sigma PB340) и 1% фенилметансульфонилфторида (PMSF, Applichem).
Клеточные лизаты инкубировали при 4°С в течение 20 мин, а затем очищали центрифугированием при 4°С в течение 20 мин при 20 000 g. Супернатанты собирали в виде неионогенных растворимых белковых фракций. Осадки промывали и ресуспендировали в PBS с добавлением 2% натриевой соли N-лауроилсаркозина (Sarkosyl, Sigma) с ингибиторами протеазы. Осадки кратковременно обрабатывали ультразвуком тонким зондом [3 раза по 3 с при амплитуде 60% (Sonic Vibra Cell, Blanc Labo, Швейцария)], инкубировали 5 мин на льду, затем центрифугировали при 100000g в течение 30 мин при 4 °С. С. Супернатанты собирали в виде растворимых в саркозиле фракций. Гранулы промывали предыдущим буфером и ресуспендировали в PBS с добавлением 2% саркозила и 8 М мочевины и кратковременно обрабатывали ультразвуком, как это делалось ранее. Буфер Лэммли 4x добавляли к образцам перед кипячением при 95 °C в течение пяти минут. Затем образцы разделяли на 16% геле SDS-PAGE перед анализом с помощью окрашивания Кумасси и WB.Для анализа WB нитроцеллюлозные мембраны блокировали в течение ночи блокирующим буфером Odyssey (LiCor, Швейцария), а затем инкубировали при комнатной температуре в течение 2 часов с первичными антителами Htt (MAB5492, Millipore, Швейцария), разведенными в PBS (1/5000).
Мембраны промывали 1% PBS-Tween, а затем инкубировали с флуоресцентно мечеными вторичными антителами, разведенными в PBS (1/5000), в течение 1 часа при комнатной температуре перед окончательной промывкой в 1% PBS-Tween. Обнаружение белка проводили по флуоресценции с использованием Odyssey CLx от LiCor. Интенсивность сигнала количественно определяли с помощью Image Studio 3.1 от LiCor.Для протеомной идентификации образцы, разделенные с помощью SDS-PAGE, окрашивали кумасси синим (25 % изопропанола [Fisher Scientific, США], 10 % уксусной кислоты [Fisher Scientific, США] и 0,05 % кумасси бриллиантового R250 [Applichem, Германия]). Каждая дорожка геля была полностью нарезана, а белки расщеплены в геле 158 . Вкратце, образцы восстанавливали в 10 мМ дитиоэритрите (DTE) (Merck, 1.24511.0005), алкилировали в 55 мМ йодацетамиде (IAA) (Sigma I1149), и кусочки геля сушили. Расщепление проводили в течение ночи при 37 °C с использованием трипсина для масс-спектрометрии (Thermofisher Scientific 9).
0057) в концентрации 12,5 нг/мкл в 50 мМ бикарбоната аммония (Sigma A6141) и 10 мМ CaCl2 (Merck 1.02382.0500). Полученные пептиды экстрагировали 70% этанолом (Merck 1.117272500), 5% муравьиной кислотой (Merck 1.00264.1000) дважды по 20 мин при постоянном встряхивании. Далее образцы высушивали вакуумным центрифугированием и хранили при -20 °С. Пептиды обессоливали на насадках 159 и сушили в вакуумном концентраторе. Для анализа методом жидкостной хроматографии с тандемной масс-спектрометрией (ЖХ-МС/МС) образцы ресуспендировали в 2% ацетонитриле (Biosolve 0001207802BS), 0,1% муравьиной кислоте и разделяли с помощью обращенно-фазовой хроматографии на системе Dionex Ultimate 3000 RSLC nano UPLC, подключенной на одной линии с Orbitrap Lumos (Thermo Fisher Scientific, Уолтем, США). Капиллярная предварительная колонка (Acclaim Pepmap C18, 3 мкм-100 Å, внутренний диаметр 2 см × 75 мкм) использовалась для захвата и очистки проб. Капиллярная колонка (внутренний диаметр 75 мкм; собственная упаковка с использованием ReproSil-Pur C18-AQ 1. 9)микрогранулы кремнезема; доктор Майш; длина 50 см) затем использовали для аналитического разделения при 250 нл/мин в течение 90 мин двухфазного градиента. Сбор данных на Orbitrap Exploris 480 выполнялся в режиме сбора данных с максимальной скоростью, зависящем от данных, с использованием времени цикла 2 с. Первые сканы масс-спектрометрии (МС) были получены с разрешением 60 000 (при 200 m/z), а наиболее интенсивные родительские ионы были отобраны и фрагментированы с помощью высокоэнергетической диссоциации столкновения (HCD) с нормализованной энергией столкновения (NCE) 30% при использовании окна изоляции 1,4 м/z. Сканирование фрагментированных ионов было получено с разрешением 15 000 (при 200 м/z), а затем выбранные ионы были исключены на следующие 20 секунд. Сбор данных на Qexactive HF был выполнен с помощью сбора данных Top N (15). Первые МС-сканы были получены с разрешением 60 000 (при 200 m/z), а наиболее интенсивные исходные ионы были отобраны и фрагментированы с помощью высокоэнергетической диссоциации столкновения (HCD) с нормализованной энергией столкновения (NCE) 27% с использованием изоляционное окно 1,4 м/з. Сканирование фрагментированных ионов было получено с разрешением 15 000 (при 200 м/z), а затем выбранные ионы были исключены на следующие 20 секунд.Идентификацию и количественный анализ белков проводили с помощью поисковой системы MaxQuant 1.6.2.10 160 . Была использована база данных Uniprot человека ( Homo Sapiens , выпуск 2019_06) или мыши ( Mus musculus , выпуск 2020_10) и последовательность Httex1. Карбамидометилирование было установлено как фиксированная модификация, тогда как окисление (М), фосфорилирование (S, T, Y), ацетилирование (N-конец белка) и глутамин до пироглутамата считались переменными модификациями. Допускалось не более двух пропущенных расколов. «Сопоставление между прогонами» было включено. Для идентификации белка было разрешено минимум 2 пептида, а порог ложного обнаружения (FDR) был установлен на уровне 0,01 как для пептидов, так и для белков. Количественная оценка и нормализация без меток выполнялись программой Maxquant с использованием алгоритма MaxLFQ со стандартными настройками 9.
1441 161 . В Perseus 162 отфильтровывались обратные белки, примеси и белки, идентифицированные только по сайтам. Данные по фракции мочевины были проанализированы отдельно в соответствии с тем же рабочим процессом. Биологические повторы были сгруппированы вместе, и группы белков, содержащие как минимум два значения LFQ, по крайней мере, в одной группе были сохранены. Пропущенные значения были заменены случайными числами с использованием распределения Гаусса (ширина = 0,7, сдвиг вниз = 1,9 для доли мочевины). Дифференциально экспрессируемые белки были выделены с помощью t-критерия для двух образцов с последующей коррекцией на основе перестановок (частота ложных открытий). Значительные попадания были определены с помощью стратегии, основанной на графике вулканов, с комбинацией P-значений t-теста с информацией о соотношении 163 . Кривые значимости на графике вулкана соответствовали значению S0, равному 0,5, и отсечке FDR, равной 0,05. Дальнейшие графические дисплеи были сгенерированы с помощью самодельных программ, написанных на R (версия 3. 6.1) 164 . В первичных нейронах был реализован биоинформатический конвейер с использованием анализа дифференциального обогащения протеомных данных (DEP) 105 . Было показано, что DEP обладает большей чувствительностью для обнаружения истинных различий между состояниями по сравнению с парными t-критериями между состояниями, поскольку общая изменчивость всех выборок используется для информирования стабилизирующей дисперсию нормализации и подхода дифференциального статистического теста. Этот конвейер был реализован в R (4.0.3 (2020-10-10)) и код доступен здесь: https://github.com/jannahastings/httex-proteomics-202107 (https://doi.org/10.5281). /zenodo.5469407). Диаграммы Венна были сгенерированы из (http://bioinformatics.psb.ugent.be/webtools/Venn/). Анализ IPA выполняли с помощью программного обеспечения (https://digitalinsights.qiagen.com/products-overview/discovery-insights-portfolio/analysis-and-visualization/qiagen-ipa/). Для анализа терминов GO использовался DAVID 6. 8 (https://david.ncifcrf.gov/). Microsoft Excel использовался для дальнейшего анализа, и в сравнительном анализе известные белки-интеракторы HTT были выбраны с использованием набора данных HDinHD (https://www.hdinhd.org/) и ограничены наборами данных человека и мыши среди клеточных или животных. исключительно учеба.Респирометрия и амплексная красная флуорометрия
Клетки HEK 293 трансфицировали через 24 часа после посева Htt 16Q, Htt 72Q, Htt 16Q-GFP или Htt 72Q-GFP. Через 48 ч после трансфекции клетки осторожно открепляли с помощью 0,05% трипсина, ресуспендировали в DMEM, подсчитывали и сразу использовали для респирометрии с высоким разрешением.
Один миллион клеток перенесли в MiR05 (0,5 мМ EGTA, 3 мМ MgCl 2 , 60 мМ лактобионата калия, 20 мМ таурина, 10 мМ KH 2 PO 4 , 20 мМ HEPES, 110 мМ сахарозы и 0,1% (масса/объем) BSA, pH = 7,1) в калиброванной 2-мл респирометрической оксиграфической камере высокого разрешения (Oroboros Instruments, Австрия) при стабильной температуре 37 °C.
Продукцию митохондриальных АФК (O 2 − и H 2 O 2 ) измеряли с помощью амплексной красной флуорометрии и модулей O2K Fluo-LED2 (Oroboros Instruments, Австрия) 165 . Вкратце, для амплексной красной флуорометрии клетки добавляли в камеры оксиграфа после добавления 10 мкМ амплексного красного, 1 ЕД/мл пероксидазы хрена и 5 ЕД/мл супероксиддисмутазы к MiR05 и калибровки с известным H 2 O 2 концентрации. Флуоресценцию измеряли во время последующего применения протокола респирометрии с высоким разрешением.Обычное дыхание (и продукцию митохондриальных АФК) измеряли в интактных клетках, после чего плазматические мембраны повышали проницаемость путем применения оптимизированной (целостность наружных мембран митохондрий подтверждена тестом на цитохром с) концентрации дигитонина (5 мкг/мл). .
Поток кислорода при различных состояниях дыхания на пермеабилизированных клетках затем определяли с использованием протокола титрования субстрат-разобщитель-ингибитор (SUIT) 166 167 .
Вкратце, дыхание NADH-пути (N) в состоянии LEAK и окислительного фосфорилирования (OXPHOS) анализировали в присутствии малата (2 мМ), пирувата (10 мМ) и глутамата (20 мМ) до и после добавления АДФ. (5 мМ) соответственно (N L , N P ). Добавление сукцината (10 мМ) позволило оценить НАДН- и сукцинат-связанное дыхание в OXPHOS (NS P ) и в несвязанном состоянии (NS E ) после постепенного (∆0,5 мкМ) добавления карбонилцианид м-хлорфенилгидразин (CCCP). Ингибирование комплекса I ротеноном (0,5 мкМ) приводило к сукцинат-связанному дыханию в несвязанном состоянии (S Е ). Тканевые удельные потоки кислорода были скорректированы на остаточное потребление кислорода, Rox, измеренное после дополнительного ингибирования митохондриальной электрон-транспортной системы, ЭТС, комплекса III антимицином А. Для дальнейшей нормализации потоки всех состояний дыхания были разделены на ЭТ-емкость на получить коэффициенты управления потоком, FCR. Терминология применялась согласно http://www. mitoeagle.org/index.php/MitoEAGLE_preprint_2018-02-08.Митохондриальные значения АФК были скорректированы с учетом фоновой флуоресценции и состояния дыхания перед добавлением разобщителя, используемого для анализа.
Анализы токсичности в первичных нейронах
Культуральный супернатант первичных кортикальных нейронов собирали в указанный момент времени и измеряли уровень дегидрогеназы молочной кислоты (ЛДГ) с использованием CytoTox 96 ® Нерадиоактивный анализ цитотоксичности ( Promega, Швейцария) 122 . Вкратце, 50 мкл реагента CytoTox 96 ® смешивали с 50 мкл собранной среды и инкубировали в течение 30 мин при комнатной температуре. После инкубации реакцию останавливали добавлением 50 мкл стоп-раствора и измеряли оптическую плотность при 49°С.0 нм с использованием планшетного ридера Tecan Infinite M200 Pro (Tecan, Maennedorf, Швейцария), что пропорционально указывает количество клеток с пермеабилизированной мембраной, приводящих к высвобождению ЛДГ в культуральной среде.
Кроме того, гибель клеток, связанная с фрагментацией ДНК, измерялась с помощью анализа TUNEL 168 . Вкратце, корковые нейроны промывали и фиксировали в 4% PFA в течение 15 минут при комнатной температуре в указанные моменты времени. Нейроны пермеабилизировали в 0,1% Triton X-100 в 0,1% цитратном буфере, pH 6,0, после чего инкубировали с терминальной дезоксинуклеотидтрансферазой и TMR red dUTP (набор для обнаружения клеточной смерти in situ; Roche, Швейцария) в течение 1 ч при 37°C. . Затем выполняли ICC, как описано выше, с нейронным маркером NeuN для выбора нейронов. Для каждого условия и для каждого независимого эксперимента, проведенного в трех экземплярах, подсчитывали минимум сто нейронов.Статистика и воспроизводимость
Микрофотографии Рис. 1b, e, g, 2a, 4d, 5a, 6a, b, 6c, 7b, e, 8a, e, S2, S3, S5, S6, S7a, S8, S9, S10, S14a, S14b, S14c, S14d, S15, S17, S19, S20a, S20b, S21a, S21b, S21c, S22, S28a и S29a являются репрезентативными изображениями трех независимых экспериментов.
Микрофотографии 1c, d из CLEM (дополнительная рис. 16a, c) представляют собой одно включение и сопоставимы с двумя другими независимыми повторами микрофотографий 1e, 2b, 4a и S3a. Микрофотографии 7c, d из CLEM (дополнительная рис. 16a, d) представляют одно включение и сопоставимы с двумя другими независимыми повторами микрофотографий 7f и S17a. Трехмерные реконструкции 1f, 1h, S7b, S20c и S20d были созданы один раз из репрезентативных электронных микрофотографий S3a, S3b, S8a, S17a и S17b соответственно. На микрофотографиях S4 представлены несколько включений из одного эксперимента. Полные и необработанные сканы WB и Coomassie доступны в исходном файле данных и на рисунках S31 и S32.Статистический анализ
Все эксперименты были независимо повторены не менее 3 раз. Статистический анализ был выполнен с использованием t-критерия Стьюдента, однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки-Крамера или HSD, двухфакторным дисперсионным анализом и повторными измерениями дисперсионного анализа с использованием KaleidaGraph (RRID: SCR_014980) или GraphPad Prism 9.
1. 1. Данные считали статистически значимыми при p < 0,05. Анализы обогащения Gene Ontology (GO) были определены с помощью анализа DAVID (значительно для p значения < 0,05 с поправкой на множественное тестирование на основе Бенджамини и Хохберга) 169 .Сводка отчета
Дополнительная информация о дизайне исследования доступна в Сводке отчета об исследовании природы, связанной с этой статьей.
Ссылки
Cattaneo, E. et al. Потеря нормальной функции гентингтина: новые разработки в исследованиях болезни Гентингтона. Trends Neurosci. 24 , 182–188 (2001).
КАС пабмед Статья Google ученый
Ross, C.A. et al. Болезнь Хантингтона: естественное течение, биомаркеры и перспективы терапии. Нац. Преподобный Нейрол. 10 , 204–216 (2014).
КАС пабмед Статья Google ученый
“>
Stout, J.C. et al. Нейрокогнитивные признаки продромальной болезни Гентингтона. Нейропсихология 25 , 1–14 (2011).
ПабМед ПабМед Центральный Статья Google ученый
Совместная исследовательская группа по болезни Гентингтона. Новый ген, содержащий тринуклеотид, который экспансирован и нестабилен на хромосомах болезни Гентингтона. Cell 72 , 971–983 (1993).
Артикул Google ученый
Кремер, Б. и др. Всемирное исследование мутации болезни Гентингтона. Чувствительность и специфичность измерения CAG повторяются. Н. англ. J. Med 330 , 1401–106 (1994).
КАС пабмед Статья Google ученый
Дж.М., Л. и др. Распространение CAG-повторов при болезни Хантингтона определяет возраст начала полностью доминантным образом.
Неврология 78 , 690–695 (2012).Артикул КАС Google ученый
Consortium GM of HD (GeM-H. Длина CAG-повтора без полиглутамина определяет время начала болезни Гентингтона. Сотовый. 178 , 887–900.e14 (2019).
Арндт, Дж. Р., Чайбва, М. и Леглейтер, Дж. Новая роль первых 17 аминокислот хантингтина при болезни Гентингтона. Биомоль. Концепции 6 , 33–46 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Atwal, R. S. et al. Гентингтин имеет сигнал ассоциации с мембраной, который может модулировать агрегацию хантингтина, проникновение в ядро и токсичность. Гул. Мол. Genet 16 , 2600–2615 (2007 г.).
КАС пабмед Статья Google ученый
Шен, К. и др. Контроль структурного ландшафта и протеотоксичности нейронов мутантного гентингтина с помощью доменов, фланкирующих тракт polyQ. Elife 5 , e18065 (2016).
ПабМед ПабМед Центральный Статья КАС Google ученый
Bäuerlein, F.J.B. et al. Архитектура in situ и клеточные взаимодействия включений полиQ. Cell 171 , 179–187.e10 (2017).
ПабМед Статья КАС Google ученый
Грубер, А. и др. Молекулярная и структурная архитектура агрегатов полиQ в дрожжах.
Проц. Натл акад. науч. 115 , E3446–E3453 (2018 г.).КАС пабмед ПабМед Центральный Статья Google ученый
Пескетт, Т. Р., Рау, Ф., О’Дрисколл, Дж., Патани, Р., Лоу, А. Р. и Сайбил, Х. Р. Переход из жидкой фазы в твердую, лежащий в основе патологической агрегации экзона 1 хантингтина. мол. Cell 70 , 588–601.e6 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Велтер, С. и др. Накопление мутантных фрагментов хантингтина в агресомоподобных тельцах включения в результате недостаточной деградации белка. Мол. биол. Ячейка 12 , 1393–1407 (2001).
КАС пабмед ПабМед Центральный Статья Google ученый
Neueder, A. et al. Белок патогенного экзона 1 HTT продуцируется неполным сплайсингом у пациентов с болезнью Гентингтона.
науч. Респ. 7 , 1307 (2017).ОБЪЯВЛЕНИЕ пабмед ПабМед Центральный Статья КАС Google ученый
Wellington, C.L. et al. Расщепление каспазы мутантного хантингтина предшествует нейродегенерации при болезни Гентингтона. J. Neurosci. 22 , 7862–7872 (2002).
КАС пабмед ПабМед Центральный Статья Google ученый
Schilling, G. et al. Характеристика патологических фрагментов хантингтина при болезни Гентингтона человека, трансгенных мышах и клеточных моделях. Дж. Невропатол. Эксп. Нейрол. 66 , 313–320 (2007).
КАС пабмед Статья Google ученый
Davies, S.W. et al. Образование нейрональных внутриядерных включений лежит в основе неврологической дисфункции у мышей, трансгенных по мутации HD.
Cell 90 , 537–548 (1997).КАС пабмед Статья Google ученый
Mangiarini, L. et al. Экзона 1 гена HD с расширенным повтором CAG достаточно, чтобы вызвать прогрессирующий неврологический фенотип у трансгенных мышей. Ячейка 87 , 493–506 (1996).
КАС пабмед Статья Google ученый
Martindale, D. et al. Длина хантингтина и его полиглутаминового тракта влияет на локализацию и частоту внутриклеточных агрегатов. Нац. Genet 18 , 150–154 (1998).
КАС пабмед Статья Google ученый
Барбаро, Б. А. и др. Сравнительное изучение встречающихся в природе фрагментов Гентингтина у дрозофилы указывает на экзон 1 как на наиболее патогенный вид при болезни Гентингтона. Гул.
Мол. Genet 24 , 913–925 (2015).КАС пабмед Статья Google ученый
Wang, H. et al. Подавление индуцированной полиглутамином токсичности в клеточных и животных моделях болезни Хантингтона с помощью убиквилина. Гул. Мол. Genet 15 , 1025–1041 (2006).
КАС пабмед Статья Google ученый
Луо, Х. и др. Herp способствует деградации мутантного хантингтина: участие протеасом и молекулярных шаперонов. Мол. Нейробиол. 55 , 7652–7668 (2018).
КАС пабмед Статья Google ученый
Scherzinger, E. et al. Самосборка фрагментов хантингтина, содержащих полиглутамин, в амилоидоподобные фибриллы: значение для патологии болезни Гентингтона. Проц. Натл акад. науч. США. 96 , 4604–4609 (1999).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Rockabrand, E. et al. Первые 17 аминокислот Хантингтина модулируют его субклеточную локализацию, агрегацию и влияние на гомеостаз кальция. Гул. Мол. Жене 16 , 61–77 (2007).
КАС пабмед Статья Google ученый
Steffan, J. S. et al. SUMO-модификация патологии Хантингтина и болезни Гентингтона. Наука (80-) 304 , 100–104 (2004).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Тэм, С. и др. Шаперонин TRiC блокирует элемент последовательности хантингтина, который способствует конформационному переключению на агрегацию. Нац. Структура Мол. биол. 16 , 1279–1285 (2009 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Kim, Y.E. et al. Растворимые олигомеры PolyQ-расширенного хантингтина нацелены на множество ключевых клеточных факторов. Мол. Cell 63 , 951–964 (2016).
КАС пабмед Статья Google ученый
Лу, М. и др. Микроскопия сверхвысокого разрешения живых клеток выявляет основную роль диффузии в сборке агресом, управляемой полиглютамином. Дж. Биол. хим. 294 , 257–268 (2019).
КАС пабмед Статья Google ученый
Scherzinger, E.
et al. Экспансия полиглутамина, кодируемая гентингтином, образует амилоидоподобные белковые агрегаты in vitro и in vivo. Cell 90 , 549–558 (1997).КАС пабмед Статья Google ученый
Холленбах, Б. и др. Агрегация укороченных слитых белков экзона 1 GST-HD, содержащих нормальный диапазон и расширенные глутаминовые повторы. Филос. Транс. Р. Соц. ЛБ биол. науч. 354 , 991–994 (1999).
КАС Статья Google ученый
Ансалони, А. и др. Полусинтез экзона 1 белка Хантингтина в одном горшке: новые инструменты для выяснения роли посттрансляционных модификаций в патогенезе болезни Гентингтона. Анжю. хим. Инт Эд. англ. 53 , 1928–1933 (2014).
КАС пабмед Статья Google ученый
Thakur, A.
K. et al. Разрушение полиглутамином N-конца экзона 1 хантингтина запускает сложный механизм агрегации. Нац. Структура Мол. биол. 16 , 380–389 (2009).КАС пабмед ПабМед Центральный Статья Google ученый
Гу, X. и др. Серины 13 и 16 являются критическими детерминантами патогенеза заболевания, вызванного полноразмерным мутантным гентингтином человека, у мышей HD. Нейрон 64 , 828–840 (2009).
КАС пабмед ПабМед Центральный Статья Google ученый
Майури, Т., Волошански, Т., Ся, Дж. и Труант, Р. Домен хантингтина N17 представляет собой многофункциональный CRM1 и Ran-зависимый ядерный и цилиальный экспортный сигнал. Гул. Мол. Genet 22 , 1383–1394 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Актар, Ф. и др. Включение хантингтина представляет собой динамическое отделение с фазовым разделением. Науки о жизни. Альянс 2 , e2019 (2019).
ПабМед ПабМед Центральный Статья Google ученый
Miao, K. & Wei, L. Визуализация живых клеток и количественная оценка агрегатов PolyQ с помощью стимулированного комбинационного рассеяния селективного мечения дейтерием. АКЦ Цент. науч. 6 , 478–486 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Софи, В.
и др. К расшифровке кода Nt17: как последовательность и конформация первых 17 аминокислот в Huntingtin регулируют агрегацию, клеточные свойства и нейротоксичность мутантного Httex1. JMB. 6, https://doi.org/10.1016/j.jmb.2021.167222 (2021).Ивата, А., Райли, Б.Е., Джонстон, Дж.А. и Копито, Р.Р. HDAC6 и микротрубочки необходимы для аутофагической деградации агрегированного гентингтина. Дж. Биол. хим. 280 , 40282–40292 (2005 г.).
КАС пабмед Статья Google ученый
Манта, Н., Дас, Н. Г. и Дас, С. К. Последние тенденции в обнаружении гентингтина и доклинические модели болезни Гентингтона. МСРН Мол. биол. 2014 , 1
(2014).
ПабМед ПабМед Центральный Статья Google ученый
Chongtham, A. et al. Влияние фланкирующих последовательностей и клеточного контекста на субклеточное поведение и патологию мутантного HTT.
Гул. Мол. Жене 29 , 674–688 (2020).КАС пабмед ПабМед Центральный Статья Google ученый
Vieweg, S., Ansaloni, A., Wang, Z.M., Warner, J.B. & Lashuel, H.A. Стратегия на основе интеина для производства белков экзона 1 гентингтина без меток позволяет по-новому взглянуть на зависимость Httex1 от полиглутамина. агрегация и образование фибрилл. Дж. Биол. хим. 291 , 12074–12086 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Азизян Р.А. и др. Установление ограничений на образование амилоида, налагаемых стерическим исключением глобулярных доменов. Дж. Мол. биол. 430 , 3835–3846 (2018).
КАС пабмед Статья Google ученый
Мацумото, Г., Ким, С.
и Моримото, Р. И. Хантингтин и мутантный SOD1 образуют агрегатные структуры с различными молекулярными свойствами в клетках человека. Дж. Биол. хим. 281 , 4477–4485 (2006 г.).КАС пабмед Статья Google ученый
Цинь, З. и др. Тельца Гентингтина секвестрируют белки, ассоциированные с везикулами, посредством полипролин-зависимого взаимодействия. J. Neurosci. 24 , 269–281 (2004).
КАС пабмед ПабМед Центральный Статья Google ученый
Suopanki, J. et al. Взаимодействие фрагментов хантингтина с мембранами головного мозга — ключ к ранней дисфункции при болезни Гентингтона. Дж. Нейрохим 96 , 870–884 (2006).
КАС пабмед Статья Google ученый
Hackam, A. S. et al.
Доказательства как ядра, так и цитоплазмы как субклеточных участков патогенеза при болезни Хантингтона в клеточной культуре и у трансгенных мышей, экспрессирующих мутантный хантингтин. Филос. Транс. Р. Соц. Б биол. науч. 354 , 1047–1055 (1999).КАС Статья Google ученый
Сапп, Э. и др. Локализация гентингтина в головном мозге в норме и у пациентов с болезнью Гентингтона. Энн. Нейрол. 42 , 604–612 (1997).
КАС пабмед Статья Google ученый
ДиФилья, М. и др. Гентингтин представляет собой цитоплазматический белок, связанный с везикулами в нейронах головного мозга человека и крысы. Нейрон 14 , 1075–1081 (1995).
КАС пабмед Статья Google ученый
Sahoo, B., Singer, D.
, Kodali, R., Zuchner, T. & Wetzel, R. Поведение агрегации химически синтезированного полноразмерного экзона гентингтина 1. Биохимия 53 , 3897–3907 (2014).КАС пабмед Статья Google ученый
Aiken, C. T. et al. Фосфорилирование треонина 3: влияние на агрегацию гентингтина и нейротоксичность. Дж. Биол. хим. 284 , 29427–29436 (2009 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Михалек М., Сальников Э. С., Вертен С. и Бехингер Б. Мембранные взаимодействия амфипатического амино-конца хантингтина. Биохимия 52 , 847–858 (2013).
КАС пабмед Статья Google ученый
Burke, K.A., Kauffman, K.J., Umbaugh, C.S., Frey, S.L. & Legleiter, J. Взаимодействие полиглутаминовых пептидов с липидными мембранами регулируется фланкирующими последовательностями, связанными с хантингтином.
Дж. Биол. хим. 288 , 14993–15005 (2013 г.).КАС пабмед ПабМед Центральный Статья Google ученый
Чаибва М. и др. Сфингомиелин и GM1 влияют на связывание гентингтина, разрушение и агрегацию на липидных мембранах. АСУ Омега 3 , 273–285 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Boatz, J.C. et al. Структура протофиламента и супрамолекулярный полиморфизм агрегированного мутантного экзона гентингтина 1. J. Mol. биол. 432 , 4722–4744 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Ди Пардо, А. и др. Мутантный хантингтин взаимодействует с белками, связывающими регуляторные элементы стерола, и нарушает их импорт в ядро.
Гул. Мол. Жене 29 , 418–431 (2020).ПабМед Статья КАС Google ученый
Ди Пардо, А. и Маглионе, В. Ось S1P: новый захватывающий путь лечения болезни Гентингтона. Тенденции Фарм. науч. 39 , 468–480 (2018).
ПабМед Статья КАС Google ученый
Валенса, М. и др. Нарушение пути биосинтеза холестерина при болезни Гентингтона. J. Neurosci. 25 , 9932–9939 (2005 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Дель Торо, Д. и др. Измененный гомеостаз холестерина способствует усилению эксайтотоксичности при болезни Гентингтона. J. Neurochem 115 , 153–167 (2010).
ПабМед Статья КАС Google ученый
Шиппер-Кром, С. и др. Динамическое рекрутирование активных протеасом в тельца включения, инициированные полиглутамином. ФЭБС Письмо. 588 , 151–159 (2014).
КАС пабмед Статья Google ученый
Huang, Z. N. & Her, L. S. Рецептор убиквитина ADRM1 модулирует активность протеасом, индуцированную HAP40. Мол. Нейробиол. 54 , 7382–7400 (2017).
КАС пабмед Статья Google ученый
Juenemann, K. et al. Динамическое рекрутирование убиквитина в мутантные гентингтиновые тельца включения.
науч. Респ. 8 , 1405 (2018).ОБЪЯВЛЕНИЕ пабмед ПабМед Центральный Статья КАС Google ученый
Hipp, M. S. et al. Непрямое ингибирование активности протеасомы 26S в клеточной модели болезни Гентингтона. J. Cell Biol. 196 , 573–587 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Bennett, E.J. et al. Глобальные изменения в системе убиквитина при болезни Гентингтона. Природа 448 , 704–708 (2007).
ОБЪЯВЛЕНИЕ КАС пабмед Статья Google ученый
Шиппер-Кром, С., Юнеманн, К. и Рейтс, Э. А. Дж. Убиквитин-протеасомная система при болезни Гентингтона: нарушены ли протеасомы, инициаторы заболевания или приходят на помощь? Biochem Res Int 2012 , 837015 (2012).
ПабМед ПабМед Центральный Статья КАС Google ученый
Bansal, M., Moharir, S.C. & Swarup, G. Рецептор аутофагии оптинеурин способствует образованию аутофагосом путем потенцирования продукции LC3-II и созревания фагофора. Комм. интегр. биол. 11 , 1–4 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Н’Диайе, Э. Н., Дебнат, Дж. и Браун, Э. Дж. Убиквилины ускоряют созревание аутофагосом и способствуют выживанию клеток при голодании по питательным веществам. Аутофагия 5 , 573–575 (2009).
ПабМед Статья Google ученый
Бейкер Б. и др. Изменение слияния лизосом и слабовыраженное воспаление, опосредованное сверхнизкими дозами эндотоксина. Дж. Биол. хим. 290 , 6670–6678 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Lu, K., Psakhye, I. & Jentsch, S. Аутофагический клиренс белков PolyQ, опосредованный адаптерами убиквитин-Atg8 семейства консервативных белков CUET. Cell 158 , 549–563 (2014).
КАС пабмед Статья Google ученый
Огуро, А., Кубота, Х., Симидзу, М., Ишиура, С. и Атоми, Ю. Защитная роль убиквитин-связывающего белка Tollip против токсичности полиглутаминовых белков расширения. Неврологи. лат. 503 , 234–239 (2011).
КАС пабмед Статья Google ученый
Симидзу М., Огуро-Андо А., Охото-Фуджита Э. и Атоми Ю. Путь белка, взаимодействующего с Toll: деградация убиквитин-связывающего белка. Том 534. 1-е изд. Эльзевир Инк https://doi.
org/10.1016/B978-0-12-397926-1.00018-4 (2014).Shen, W.C., Li, H.Y., Chen, G.C., Chern, Y. & Tu, P.H. Мутации в убиквитин-связывающем домене OPTN/оптиневрина препятствуют аутофагии-опосредованной деградации неправильно свернутых белков по доминантно-негативному механизму . Аутофагия 11 , 685–700 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Korac, J. et al. Убиквитин-независимая функция оптиневрина в аутофагическом клиренсе белковых агрегатов. J. Cell Sci. 126 , 580–592 (2013).
КАС пабмед Статья Google ученый
Schwab, C., Yu, S., McGeer, E.G. & McGeer, P.L. Оптинейрин при внутриядерных включениях при болезни Гентингтона. Неврологи. лат. 506 , 149–154 (2012).
КАС пабмед Статья Google ученый
Денг З. и др. Рецепторы аутофагии и нейродегенеративные заболевания. Trends Cell Biol. 27 , 491–504 (2017).
КАС пабмед Статья Google ученый
Suhr, S. T. et al. Идентичность секвестрированных белков в агрегатах из клеток с индуцированной экспрессией полиглутамина. J. Cell Biol. 153 , 283–294 (2001).
КАС пабмед ПабМед Центральный Статья Google ученый
Казанцев А., Прейзингер Э., Драновски А., Гольдгабер Д. и Хаусман Д. Между патологическими и непатологическими участками полиглутамина в клетках млекопитающих образуются нерастворимые устойчивые к детергентам агрегаты.
Проц. Натл акад. науч. США 96 , 11404–11409 (1999).ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Scior, A. et al. Полное подавление фибрилляции Htt и дезагрегация фибрилл Htt с помощью тримерного шаперонового комплекса. EMBO J. 37 , 282–299 (2018).
КАС пабмед Статья Google ученый
Hjerpe, R. et al. UBQLN2 опосредует независимый от аутофагии клиренс белковых агрегатов протеасомой. Cell 166 , 935–949 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Калвер, Б. П. и др. Белок болезни Гентингтона гентингтин ассоциирует со своей собственной мРНК. Дж. Хантингт. Дис. 5 , 39–51 (2016).
КАС Статья Google ученый
Feyeux, M. et al. Ранние транскрипционные изменения, связанные с естественными мутациями болезни Хантингтона в нейральных производных эмбриональных стволовых клеток человека. Гул. Мол. Genet 21 , 3883–3895 (2012).
КАС пабмед Статья Google ученый
Боровецкий Ф. и др. Полногеномное профилирование экспрессии крови человека выявляет биомаркеры болезни Гентингтона. Проц. Натл акад. науч. США 102 , 11023–11028 (2005 г.).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Мастроколиас, А. и др. Профиль прогрессирования биомаркера болезни Гентингтона, определенный секвенированием транскриптома в периферической крови. евро. Дж. Хам. Genet 23 , 1349–1356 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Кроче К.Р. и Ямамото А. Роль аутофагии при болезни Гентингтона. Нейробиол. Дис. 122 , 16–22 (2019).
КАС пабмед Статья Google ученый
Snapp, E.L. et al. Формирование штабелированных цистерн ER за счет взаимодействий с белками с низким сродством.
J. Cell Biol. 163 , 257–269 (2003).КАС пабмед ПабМед Центральный Статья Google ученый
McCaughey, J. et al. TFG способствует организации переходного ER и эффективной секреции коллагена. Cell Rep. 15 , 1648–1659 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Витте, К. и др. Функция TFG-1 в секреции белка и онкогенезе. Нац. Клеточная биол. 13 , 550–558 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Tellez-Nagel, I., Johnson, A.B. & Terry, R.D. Исследования биопсии головного мозга пациентов с хореей Гентингтона. Дж. Невропатол. Эксп. Нейрол. 33 , 308–332 (1974).
КАС пабмед Статья Google ученый
Майури, Т. и др. Репарация ДНК при болезни Гентингтона и спиноцеребеллярных атаксиях: соматическая нестабильность и альтернативные гипотезы. Дж. Хантингт. Дис. 10 , 165–173 (2021).
КАС Статья Google ученый
Ратовицкий Т. и др. Взаимодействия с белком гентингтина изменены экспансией полиглутамина, как определено количественным протеомным анализом. Клеточный цикл 11 , 2006–2021 (2012).
ПабМед ПабМед Центральный Статья Google ученый
Хосп, Ф. и др. Пространственно-временное протеомное профилирование включений при болезни Гентингтона выявляет обширную потерю функции белка. Cell Rep. 21 , 2291–2303 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Hodges, A. et al. Экспрессия генов в мозге коррелирует с изменениями поведения у мышей R6/1, моделирующих болезнь Гентингтона. Гены, Поведение Мозга. 7 , 288–299 (2008).
КАС Статья Google ученый
Christodoulou, C.C. et al.
Изучение перехода от предсимптомного к симптоматическому статусу болезни Гентингтона на основе данных омики. Int J. Mol. науч. 21 , 7414 (2020).КАС ПабМед Центральный Статья Google ученый
Сафрен, Н., Чанг, Л., Дзики, К. М. и Монтейро, М. Дж. Характерные изменения экспрессии убиквилина в мышиной модели болезни Гентингтона R6/2. Brain Res 1597 , 37–46 (2015).
КАС пабмед Статья Google ученый
Safren, N. et al. Сверхэкспрессия убиквилина-1 увеличивает продолжительность жизни и задерживает накопление агрегатов хантингтина в мышиной модели болезни Гентингтона R6/2. PLoS One 9 , e87513 (2014 г.).
ОБЪЯВЛЕНИЕ пабмед ПабМед Центральный Статья КАС Google ученый
Wear, M. P. et al. Белки с внутренне неупорядоченными доменами предпочтительно рекрутируются в агрегаты полиглутамина. PLoS One 10 , e0136362 (2015 г.).
ПабМед ПабМед Центральный Статья КАС Google ученый
Мизуками, К. и др. Иммуногистохимический анализ убиквилина-1 в гиппокампе человека: связь с патологией нейрофибриллярных клубков. Невропатология 34 , 11–18 (2014).
КАС пабмед Статья Google ученый
Дил Куази, А. и др. Белок NEDD8 участвует в убиквитинированных тельцах включения.
Дж. Патол. 199 , 259–266 (2003).ПабМед Статья КАС Google ученый
Мори, Ф. и др. Накопление NEDD8 в нейрональных и глиальных включениях при нейродегенеративных заболеваниях. Невропатология. Заявл. Нейробиол. 31 , 53–61 (2005).
КАС пабмед Статья Google ученый
Maghames, C.M. et al. NEDDилирование способствует агрегации ядерных белков и защищает протеасомную систему убиквитина при протеотоксическом стрессе. Нац. коммун. 9 , 4376 (2018).
ОБЪЯВЛЕНИЕ пабмед ПабМед Центральный Статья КАС Google ученый
Рэй, П. Д., Хуанг, Б.-В. & Tsuji, Y. Гомеостаз активных форм кислорода (АФК) и регуляция окислительно-восстановительного потенциала в клеточной передаче сигналов.
Cell Signal 24 , 981–990 (2012).КАС пабмед ПабМед Центральный Статья Google ученый
Chang, D.T.W., Rintoul, G.L., Pandipati, S. & Reynolds, I.J. Мутантные агрегаты гентингтина нарушают движение митохондрий и транспорт нейронов коры головного мозга. Нейробиол. Дис. 22 , 388–400 (2006).
КАС пабмед Статья Google ученый
Босси-Ветцель, Э., Петрилли, А. и Нотт, А.Б. Мутантный хантинтин и митохондриальная дисфункция. Trends Neurosci. 31 , 609–616 (2008).
КАС пабмед ПабМед Центральный Статья Google ученый
Кротти, А. и др. Мутантный Huntingtin способствует автономной активации микроглии через факторы, определяющие миелоидную линию. Нац. Неврологи.
17 , 513–521 (2014).КАС пабмед ПабМед Центральный Статья Google ученый
Manczak, M. P. & Hemachandra, R. Ингибитор митохондриального деления 1 защищает от мутантного гентингтина, вызванного аномальной митохондриальной динамикой и повреждением нейронов при болезни Гентингтона. Гул. Мол. Genet 24 , 7308–7325 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Yin, X., Manczak, M. & Reddy, P. H. Молекулы MitoQ и SS31, нацеленные на митохондрии, снижают митохондриальную токсичность, вызванную мутантным хантингтином, и синаптическое повреждение при болезни Гентингтона. Гул. Мол. Жене 25 , 1739–1753 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Хорн А., Раавичарла С., Шах С., Кокс Д. и Джайсвал Дж. К. Фрагментация митохондрий обеспечивает локализованную передачу сигналов, необходимую для восстановления клеток. J. Cell Biol. 219 , e201
4 (2020).КАС пабмед ПабМед Центральный Статья Google ученый
Форно, Л. С. и Норвилл, Р. Л. Ультраструктура телец Леви в звездчатом ганглии. Акта Нейропатол. 34 , 183–197 (1976).
КАС пабмед Статья Google ученый
Gai, W.
P. et al. In situ и in vitro исследование колокализации и сегрегации α-синуклеина, убиквитина и липидов в тельцах Леви. Экспл. Нейрол. 166 , 324–333 (2000).КАС пабмед Статья Google ученый
Шахморадян С.Х. и др. Патология Леви при болезни Паркинсона состоит из переполненных органелл и липидных мембран. Нац. Неврологи. 22 , 1099–1109 (2019).
КАС пабмед Статья Google ученый
Махуль-Мелье, А.-Л. и другие. Процесс образования телец Леви, а не просто фибрилляция α-синуклеина, является одним из основных факторов нейродегенерации. Проц. Натл акад. науч. США 117 , 4971–4982 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
ДиФилья, М. и др.
Агрегация хантингтина в нейрональных внутриядерных включениях и дистрофических нейритах головного мозга. Наука 277 , 1990–1993 (1997).КАС пабмед Статья Google ученый
Choo, Y.S., Johnson, G.V.W., MacDonald, M., Detloff, P.J. & Lesort, M. Мутантный хантингтин непосредственно увеличивает восприимчивость митохондрий к индуцированному кальцием переходу проницаемости и высвобождению цитохрома c. Гул. Мол. Genet 13 , 1407–1420 (2004).
КАС пабмед Статья Google ученый
Song, W. et al. Мутантный хантингтин связывает митохондриальную ГТФазу расщепления динамин-родственного белка-1 и повышает его ферментативную активность. Нац. Мед 17 , 377–382 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Гу, М. и др. Дефект митохондрий в хвостатом ядре при болезни Гентингтона. Энн. Нейрол. 39 , 385–389 (1996).
КАС пабмед Статья Google ученый
Browne, S.E. et al. Окислительное повреждение и метаболическая дисфункция при болезни Гентингтона: избирательная уязвимость базальных ганглиев. Энн. Нейрол. 41 , 646–653 (1997).
КАС пабмед Статья Google ученый
Kegel, K.B. et al. Экспрессия гентингтина стимулирует эндосомально-лизосомальную активность, тубуляцию эндосом и аутофагию.
J. Neurosci. 20 , 7268–7278 (2000).КАС пабмед ПабМед Центральный Статья Google ученый
Stephens, D. J. Формирование de novo, слияние и деление участков выхода эндоплазматического ретикулума, покрытых COPII млекопитающих. EMBO Rep. 4 , 210–217 (2003).
КАС пабмед ПабМед Центральный Статья Google ученый
Фархан, Х., Вайс, М., Тани, К., Кауфман, Р. Дж. и Хаури, Х. П. Адаптация мест выхода эндоплазматического ретикулума к острому и хроническому увеличению грузовой нагрузки. EMBO J. 27 , 2043–2054 (2008).
КАС пабмед ПабМед Центральный Статья Google ученый
Видаль Р., Кабальеро Б., Кув А. и Хетц С. Конвергентные пути возникновения стресса эндоплазматического ретикулума (ЭР) при болезни Гентингтона.
Курс. Мол. Мед 11 , 1–12 (2011).КАС пабмед Статья Google ученый
Jiang, R., Diaz-Castro, B., Looger, L.L. & Khakh, B.S. Дисфункциональная передача сигналов кальция и глутамата в полосатых астроцитах мышей с моделью болезни Гентингтона. J. Neurosci. 36 , 3453–3470 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Lee, K. S. et al. Измененный контакт ER-митохондрий влияет на гомеостаз кальция в митохондриях и способствует нейродегенерации in vivo в моделях заболеваний. Проц. Натл акад. науч. США 115 , E8844–E8853 (2018 г.).
КАС пабмед ПабМед Центральный Google ученый
Эль-Дахер, М.-Т. и другие. Протеолиз гентингтина высвобождает фрагменты, отличные от полиQ, которые вызывают токсичность из-за дисрегуляции динамина-1.
EMBO J. 34 , 2255–2271 (2015).КАС пабмед ПабМед Центральный Статья Google ученый
Leitman, J., Ulrich Hartl, F. & Lederkremer, G. Z. Растворимые формы полиQ-расширенного гентингтина, а не большие агрегаты, вызывают стресс эндоплазматического ретикулума. Нац. коммун. 4 , 2753 (2013).
ОБЪЯВЛЕНИЕ пабмед Статья КАС Google ученый
Higo, T. et al. Механизм повреждения головного мозга, вызванного стрессом ER, с помощью рецептора IP (3). Нейрон 68 , 865–878 (2010).
КАС пабмед Статья Google ученый
Петраш-Парвез, Э. и др. Клеточная и субклеточная локализация агрегатов Хантингтина в головном мозге крысы, трансгенной по болезни Гентингтона. Дж.
Комп. Нейрол. 501 , 716–730 (2007).ПабМед Статья Google ученый
Тагава, К. и др. Различные модели агрегации и гибели клеток среди различных типов первичных нейронов, индуцированные мутантным гентингтиновым белком. J. Neurochem 89 , 974–987 (2004).
КАС пабмед Статья Google ученый
Peters, M. F. et al. Ядерное нацеливание на мутантный хантингтин увеличивает токсичность. Мол. Клеточные нейробиологи. 14 , 121–128 (1999).
КАС пабмед Статья Google ученый
Benn, C.L. et al. Вклад ядерной и внеядерной полиQ в неврологические фенотипы в мышиных моделях болезни Гентингтона. Гул. Мол. Genet 14 , 3065–3078 (2005).
КАС пабмед Статья Google ученый
DeGuire, S.M. et al. Фосфорилирование N-концевого гентингтина (Htt) представляет собой молекулярный переключатель, регулирующий агрегацию Htt, спиральную конформацию, интернализацию и нацеливание на ядро. J. Biol. хим. 293 , 18540–18558 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Ли, Л. и др. Визуализация агрегатов Хантингтина в режиме реального времени отклоняет поиск мишеней и транскрипцию генов. Elife 5 , e17056 (2016).
ПабМед ПабМед Центральный Статья Google ученый
Posey, A.
E. et al. Профилин снижает агрегацию и фазовое разделение N-концевых фрагментов хантингтина за счет предпочтительного связывания с растворимыми мономерами и олигомерами. Дж. Биол. хим. 293 , 3734–3746 (2018).КАС пабмед ПабМед Центральный Статья Google ученый
Пуарье, М. А. и др. Сфероиды и протофибриллы Хантингтина как предшественники полиглутаминовой фибрилляции. Дж. Биол. хим. 277 , 41032–41037 (2002 г.).
КАС пабмед Статья Google ученый
Liu, K.Y. et al. Разрушение ядерной мембраны перинуклеарными включениями мутантного гентингтина вызывает повторный вход в клеточный цикл и гибель стриарных клеток в мышиных и клеточных моделях болезни Гентингтона. Гул. Мол. Жене 24 , 1602–1616 (2015).
КАС пабмед Статья Google ученый
Миллер, Дж. и др. Количественные взаимосвязи между уровнями хантингтина, длиной полиглутамина, образованием телец-включений и гибелью нейронов обеспечивают новое понимание молекулярного патогенеза болезни Хантингтона. Дж. Неврологи. 30 , 10541–10550 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
Колла, Р. и др. Новый химико-ферментативный полусинтетический подход дает представление о роли фосфорилирования за пределами экзона 1 хантингтина и выявляет различные механизмы агрегации, зависящие от длины n-концевого фрагмента. Дж. Ам. хим. соц. 143 , 9798–9812 (2021).
КАС пабмед Статья Google ученый
Барде И., Салмон П. и Троно Д. Производство и титрование лентивирусных векторов. Ч. 4 (Единица 4.21) Curr Protoc Neurosci. 1–23 https://doi.org/10.1002/0471142301.ns0421s53 (2010 г.).
Джордан, М., Шаллхорн, А., Вурм, Ф. М. и Франциско, С. С. Трансфекция клеток млекопитающих: оптимизация критических параметров, влияющих на образование кальций-фосфатного осадка. Nucleic Acids Res 24 , 596–601 (1996).
КАС пабмед ПабМед Центральный Статья Google ученый
Steiner, P. et al. Модуляция цикла рецепторов обогащенным нейронами эндосомальным белком 21 кДа. J. Cell Biol. 157 , 1197–1209 (2002).
КАС пабмед ПабМед Центральный Статья Google ученый
Чен, М. и др. Сверточные нейронные сети для автоматизированной аннотации клеточных криоэлектронных томограмм. Нац. Методы 14 , 983–985 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Кремер, Дж. Р., Мастронард, Д. Н. и Макинтош, Дж. Р. Компьютерная визуализация данных трехмерного изображения с использованием IMOD. Дж. Структура. биол. 116 , 71–76 (1996).
КАС пабмед Статья Google ученый
Дорин-Семблат, Д. и др. Эритроциты, инфицированные малярийным паразитом, секретируют PfCK1, плазмодий-гомолог плейотропной протеинкиназы казеинкиназы 1. PLoS One 10 , 1–22 (2015).
Артикул КАС Google ученый
Rappsilber, J., Mann, M. & Ishihama, Y. Протокол микроочистки, обогащения, предварительного фракционирования и хранения пептидов для протеомики с использованием StageTips. Нац. протокол 2 , 1896–1906 (2007).
КАС пабмед Статья Google ученый
Cox, J. & Mann, M. MaxQuant обеспечивает высокую скорость идентификации пептидов, индивидуальную точность массы в диапазоне частей на миллиард и количественный анализ белков в масштабах всего протеома.
Нац. Биотехнолог. 26 , 1367–1372 (2008).КАС пабмед Статья Google ученый
Кокс, Дж. и др. Точная количественная оценка без меток по всему протеому за счет отсроченной нормализации и извлечения максимального соотношения пептидов, называемая MaxLFQ. Мол. Клеточный протеом. 13 , 2513–2526 (2014).
КАС Статья Google ученый
Тьянова С. и др. Вычислительная платформа Perseus для комплексного анализа (проте)омных данных. Нац. Методы 13 , 731–740 (2016).
КАС пабмед Статья Google ученый
Hubner, N.C. et al. Количественная протеомика в сочетании с BAC TransgeneOmics выявляет взаимодействия белков in vivo. J. Cell Biol. 189 , 739–754 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
Макрекка-Кука М., Крумшнабель Г. и Гнайгер Э. Респирометрия высокого разрешения для одновременного измерения потоков кислорода и перекиси водорода в пермеабилизированных клетках, гомогенате тканей и изолированных митохондриях. Биомолекулы 5 , 1319–1338 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Pesta, D. & Gnaiger, E. Респирометрия высокого разрешения: протоколы OXPHOS для клеток человека и пермеабилизированных волокон из небольших биопсий мышц человека bt – митохондриальная биоэнергетика: методы и протоколы. В Palmeira, CM, Moreno, AJ & Totowa, NJ (ред.) 25–58. https://doi.org/10.1007/978-1-61779-382-0_3 (Humana Press, 2012).
Burtscher, J., Zangrandi, L., Schwarzer, C. & Gnaiger, E. Различия в функции митохондрий в гомогенизированных образцах здоровых и эпилептических специфических тканей мозга, выявленные с помощью респирометрии высокого разрешения.
Митохондрия 25 , 104–112 (2015).КАС пабмед Статья Google ученый
Гавриэли Ю., Шерман Ю. и Бен-Сассон С.А. Идентификация запрограммированной гибели клеток in situ посредством специфического мечения фрагментации ядерной ДНК. J. Cell Biol. 119 , 493–501 (1992).
КАС пабмед Статья Google ученый
Бенджамини Ю. и Хохберг Ю. Контроль уровня ложных открытий: практичный и мощный подход к множественному тестированию. JR Stat. соц. сер. Б. 57 , 289–300 (1995).
MathSciNet МАТЕМАТИКА Google ученый
Thompson, L.M. et al. IKK фосфорилирует гентингтин и нацеливает его на деградацию протеасомой и лизосомой. J. Cell Biol. 187 , 1083–1099 (2009).
КАС пабмед ПабМед Центральный Статья Google ученый
Zheng, Z., Li, A., Holmes, B.B., Marasa, J.C. & Diamond, M.I. N-концевой ядерный экспортный сигнал регулирует перенос и агрегацию экзона 1 белка хантингтина (Htt). J. Biol. хим. 288 , 6063–6071 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Хазеки, Н. и др. Ультраструктура ядерных агрегатов, образованных экспрессией расширенного полиглутамина. Биохим Биофиз. Рез общ. 294 , 429–440 (2002).
КАС пабмед Статья Google ученый
Лаббадиа Дж. и Моримото Р. И. Болезнь Хантингтона: основные молекулярные механизмы и новые концепции. Trends Biochem Sci. 38 , 378–385 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Маруяма Х. и Каваками Х. Оптинейрин и боковой амиотрофический склероз. Гериатр. Геронтол. Int 13 , 528–532 (2013).
ПабМед Статья Google ученый
Bowles, K.R. et al. Факторы транскрипции SMAD изменяются в клеточных моделях HD и регулируют экспрессию HTT. Cell Signal 31 , 1–14 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Runne, H. et al. Анализ потенциальных транскриптомных биомаркеров болезни Гентингтона в периферической крови. Проц. Натл акад. науч. США 104 , 14424–14429 (2007 г.).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Gasset-Rosa, F. et al. Гентингтин, расширенный полиглутамином, усугубляет возрастное нарушение ядерной целостности и ядерно-цитоплазматического транспорта. Нейрон 94 , 48–57.e4 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Сап, К. А. и др. Глобальные изменения протеома и убиквитинома в растворимой и нерастворимой фракциях мозга мышей Генгингтона Q175. Мол. Клеточный протеом. 18 , 1705–1720 (2019).
КАС Статья Google ученый
Стикер, А., Гёмин, Л., Мартенс, Л. и Клемент, Л. Надежное обобщение и вывод в количественном анализе без меток по всему протеому. Мол. Клеточный протеом. 19 , 1209–1219 (2020).
Артикул Google ученый
Ugalde, C.L. et al. Неправильно свернутый α-синуклеин вызывает гиперактивное дыхание без функционального дефицита в живых клетках нейробластомы. Дис. Модель мех. 13 , dmm040899 (2020).
КАС пабмед ПабМед Центральный Google ученый
Brennan, W.A., Bird, E.D. & Aprille, J.R. Региональная митохондриальная дыхательная активность в головном мозге при болезни Гентингтона. J. Neurochem 44 , 1948–1950 (1985).
КАС пабмед Статья Google ученый
Greiner, E. R. & Yang, X. W. Болезнь Гентингтона: щелкнуть выключателем на хантингтине. Нац. хим. биол. 7 , 412–414 (2011).
КАС пабмед Статья Google ученый
Firdaus, W.J.J. et al. Тельца включения Гентингтина являются железозависимыми центрами окислительных событий. FEBS J. 273 , 5428–5441 (2006).
КАС пабмед Статья Google ученый
Mahul-Mellier, A.L. et al. C-Abl фосфорилирует α-синуклеин и регулирует его деградацию: влияние на клиренс α-синуклеина и вклад в патогенез болезни Паркинсона. Гул. Мол. Genet 23 , 2858–2879 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Петтерсен, Э. Ф. и др. UCSF chimera — система визуализации для поисковых исследований и анализа. J. Comput Chem. 25 , 1605–1612 (2004).
КАС пабмед Статья Google ученый
CoreTeam R. R: язык и среда для статистических вычислений. Том 2.; 2019.
Скачать ссылки
Часто задаваемые вопросы: Создание переменных, записывающих свойства других членов группы
Как создать переменные, обобщающие для каждого отдельного свойства другие члены группы?
| Заголовок | Создание переменных, записывающих свойства других членов группы | |
| Автор | Николас Дж. Кокс, Даремский университет, Великобритания |
Начиная с 2016 года, программа сообщества rangestat (SSC) предлагает альтернатива решениям здесь. Вы вполне могли бы установить его и изучить его подробная помощь.
1. Примеры: данные о семьях
Предположим, у вас есть данные о семьях. Для каждого человека в каждой семье это может быть полезно для вычисления переменных, обобщающих свойства других членов одной семьи. Сколько еще детей? Что их средний, максимальный или минимальный возраст? Есть старший ребенок или младший ребенок? Более общая проблема может быть описана как обобщение свойств, для каждого человека, других членов той же группы.
Давайте посмотрим на некоторые выдуманные данные. Для дальнейшего необходимо
иметь идентификатор группы, поэтому в этом примере у нас есть идентификатор для каждого
семья. Не всегда обязательно иметь индивидуальный идентификатор, но
то, что следует далее, зависит от того, что каждый человек встречается только один раз в
набор данных. Однако на практике такие данные обычно включают индивидуальные
идентификаторы.
член семьи пол возраст 1. 1 1 1 36 2. 1 2 1 16 3. 1 3 1 14 4. 2 1 0 45 5. 2 2 1 42 6. 2 3 0 14 7. 2 4 1 12 8. 2 5 0 10 9. 3 1 0 39 10. 3 2 1 36 11. 3 3 0 11 12. 3 4 1 913. 3 5 1 7 14. 3 6 1 3
Предположим, что пол записывается как 1 для женского пола и 0 для мужского пола. Такое кодирование 0–1 в некотором смысле произвольно, но облегчает жизнь. особенно для статистического моделирования, в котором ответ является бинарным переменной и (что более важно здесь) для подсчета значений внутри каждого группа.
2. Конкретная проблема: сколько других детей на каждого ребенка?
Определим детей как тех, чей возраст составляет 17 лет и младше. Для каждого
ребенок, сколько еще детей? Это просто количество
детей в семье, минус 1, если каждый человек является ребенком. (в семье 3,
с 4 детьми, на каждого ребенка приходится 3 других ребенка. )
При любом расчете, подобном этому, всегда стоит посмотреть, Эген предоставляет ответ хотя бы на часть проблемы. написано много функций для egen . В частности, egen, total() by() естественно для подведение итогов, включая подсчеты, отдельно для групп, определенных одним или больше переменных, указанных в качестве аргументов для by() . эген, кол() by() также часто бывает полезен, но он немного менее универсален в применении, поэтому здесь мы сконцентрируемся на total() . Всего () в Стате 9 и более поздние выпуски являются заменой sum() в Stata 8.
. egen nchild = всего (возраст <= 17), по (семье) . заменить nchild = nchild - (возраст <= 17)
age < = 17 будет истинным (оценивается как 1) всякий раз, когда age равно
меньше или равно 17, а в противном случае — false (оценивается как 0). Сложение
1 и 0 в пределах egen, total() то же самое, что и подсчет
наблюдения, для которых возраста <= 17 . Затем мы вычитаем возраста <=
17 из каждого наблюдения. Эффект от опции by (family) :
для подсчета внутри семей, каждая семья представляет собой группу наблюдений с
то же значение семейство . Эффект исправления заменяет ограничиться отдельными наблюдениями.
Синтаксис egen указывает, что total() работает на выражение выражение . Аргумент не обязательно должен быть одной переменной, но может полезно быть чем-то более сложным. Заинтересованность только в другом с детьми женского пола не сложнее:
. egen nsisters = total (возраст <= 17 & пол == 1), by (семья) . заменить nsisters = nsisters - (возраст <= 17 и пол == 1)
Это решение также присваивает значения взрослым, тем, у кого возраст старше.
больше или равно 18. Это может быть полезно или бесполезно, в зависимости от вашего
содержательная проблема. Если вы хотите полностью исключить взрослых из
расчет, вы можете указать , если возраст <= 17 на egen команда, и значения для взрослых будут отсутствовать ( . ).
Если бы мы хотели считать не «других детей», а «других взрослые», мы должны быть немного более осторожными. Выражение возраст >= 18 включает пропущенные значения для age , как в Stata отсутствует считается выше, чем любое другое числовое значение. Часто мы хотим исключить те с условием возраст >= 18 & возраст < . если мы не знаем мы можем относиться к отсутствующим возрастам как к взрослым.
3. Общая задача: суммы и средние
Другие итоги и дополнительные средства могут быть рассчитаны с использованием того же Основной подход. Проще говоря,
- Подсчитайте сумму для каждой группы.
- Вычесть вклад каждого члена из этой суммы (возможно, вклад равен 0).
- При необходимости рассчитайте среднее значение как сумму, деленную на количество ценности.
Каков средний возраст остальных детей в каждой семье? Вот один решение:
. egen totalage = total(возраст), если возраст <= 17, по (семье) .
Это решение исключает взрослых. Они не только не включены в сумма age , но они также получают пропущенные значения для результат. В команде replace мы можем небрежно исключать или включая взрослых; в любом случае пропущенные значения не будут измененный.
Если мы хотим включить взрослых, то есть нам нужна запись для каждого взрослый среднего возраста детей — вот решение:
. egen totalage = total(возраст * (возраст <= 17)), by(семья) . заменить totalage = totalage - age * (возраст <= 17) . генерировать среднее значение = общее количество/ребенок
Здесь множитель age <= 17 говорит, что слагаемое равно 0 всякий раз, когда возраст равен 18 или больше, поэтому сумма является правильной суммой и присваивается всем наблюдениям в каждой семье.
4. Общая проблема: другие статистические данные
То, что мы сделали до сих пор, довольно тонко зависит от двух свойств сумм:
во-первых, сумма для «всех остальных» — это всего лишь сумма для
«все» минус сумма (значение) для этого наблюдения; а также
во-вторых, на значение суммы не влияет добавление или вычитание 0. Когда мы обращаемся к другим сводным статистическим данным, мы больше не можем полагаться на эти данные.
характеристики. Нам нужен более общий подход.
В общих чертах, нам нужно выполнить работу внутри цикла:
для каждого члена семьи {
рассчитать статистику по данным о семье
присвоить результат этому члену этой семьи
}
5. Особая проблема: максимальный возраст других детей
Предположим, что мы хотим знать для каждого ребенка максимальный возраст другие дети в той же семье. Внутри цикла мы найдем сами присваиваем куски значений: для этой задачи мы не можем использовать повторно генерирует . Мы можем многократно использовать вместо , поэтому нам нужно to сгенерировать переменную, прежде чем мы сможем это сделать:
. генерировать maxage = .
Далее нам нужен идентификатор от 1 и выше, чтобы назначить каждому человеку в семье. В нашем небольшом наборе данных уже был такой идентификатор, но, если бы его не было, его можно было бы легко создать, используя с сортировка вариант:
.
Under by varlist : _n интерпретируется внутри каждой группы наблюдений, а не для всего набора данных. Для этой задачи не дело в том, что pid произволен; нам просто нужен систематический способ делать расчеты по очереди для каждого члена семьи. резюмируют показывает нам максимальное значение pid , которое нам скоро понадобится. Мы также можно подобрать значение максимума как r(max) , то есть важно для любой автоматизации всего процесса.
Внутри цикла нам нужен способ исключения каждого значения pid из расчет. Вот один из способов сделать это, используя для значений:
. спокойно forvalues i = 1/`r(max)' {
. сгенерировать include = 1, если pid != `i' & age <= 17
. egen work = max(возраст * включает), by(семья)
. заменить maxage = работать, если pid == `i'
. бросить включить работу
. }
Конструкция forvalues перебирает значения локального макроса. i , который устанавливается по очереди в 1, затем в 2 и так далее, до максимума
из pid , возвращенного , суммировать . Макрос автоматически
увеличивается каждый раз в цикле. На практике большинство программистов Stata
используйте аббревиатуру вместо . Внутри цикла значение i упоминается как ‘i’ . Оператор generate создает
переменная, равная 1, если наблюдение должно быть включено в расчет
и отсутствует в противном случае. Выражение age * включает , что тогда
подается на egen, max() , age * 1 или age когда включает 1, а возраст *. или отсутствует . когда включает отсутствует. Что делает egen, max() , так это исключает отсутствующие
из расчета, и, только если все значения в каждой группе отсутствуют,
максимум будет возвращен как отсутствующий. Хотя у Stata есть общее правило
что отсутствующее числовое значение больше, чем любое другое числовое значение, предполагается, когда
вычисляя максимумы, которые вам действительно нужны, чтобы получить наибольшее непропущенное значение. (Видеть
что происходит, когда вы набираете display max(1,2,_pi,42,.) .) Затем мы используем
результат этого расчета на заменить значением maxage для
текущий член семьи. Наконец, проще всего сбросить переменные включают , а работают с , чтобы Stata могла начать заново
время вокруг петли.
Почему этот цикл не является следующим кодом?
. спокойно forvalues i = 1/`r(max)' {
. egen work = max(age) if age <= 17 & pid != `i', by(family)
. заменить maxage = работать, если pid == `i'
. бросить работу
. }
Причина, по которой это не будет работать должным образом, заключается в Расчет egen будет отсутствовать для наблюдений, исключенных если условие . Фактически результатом цикла является то, что все значения maxage будет отсутствовать.
На каждого ребенка есть старший (строго один или несколько), если maxage больше age ,
.
и мы могли бы использовать аналогичный подход, чтобы получить минимальный возраст другого детей и, таким образом, определить, есть ли младшие дети.
Та же общая схема может быть использована для других функций egen , которые принять выражение exp в качестве аргумента и разрешить by() в качестве вариант; видеть эген.
6. Конкретная проблема: сколько собственных детей в семье?
Рассмотрим семейный опрос, в котором у нас нет прямой информации о количество детей у каждого человека. У нас есть переменные для идентификатора семьи семейный и индивидуальный ID человек а также ID отца отца и матери ID матери (которые отсутствуют, если лицо мать или отец не являются членами одной семьи). Таким образом, в примере
член семьи отецм матьм 1 1 . . 1 2 . . 1 3 1 2 1 4 1 2 1 5 1 2 2 1 . .
семья 1 включает пару и троих детей, все дети те же мать и отец, тогда как семья 2 включает бабушку, ее дочь и внук – сын или дочь этой дочери.
Проблема состоит в том, чтобы создать переменную ownchild , задающую количество собственные дети каждого человека, проживающие в семье. Таким образом, в семье 1, у обоих родителей проживает по трое детей, тогда как в семье 2 оба с бабушкой и ее дочерью проживает по одному ребенку.
Сначала найдем количество детей каждого отца и каждой матери:
. по отцу семьи, сортировать: род fребенок = _N, если отецm < . . по матери m, отсортировать: gen mchild = _N, если motherm < .
Под по varlist : _N интерпретируется внутри каждой группы наблюдений, а не для всего набора данных. Теперь мы инициализируем переменную которая будет произведена, и переменная, которая нам понадобится для ее создания. Оба могут быть байт переменных:
.
Мы собираемся перебрать значения человек в каждой семье. Мы можем видеть в примере, что они варьируются от 1 до 5, но в более общем случае мы можем подобрать максимум из суммировать , как и в предыдущем проблема:
. резюмировать человека, только
Основной цикл такой, который мы сначала посмотрим, а потом распакуем:
. forval i = 1 / `r(max)' {
. заменить ischild = (fatherm == `i') | (мать == `я')
. #разделитель ;
. qui по семейству (ischild), сортировать:
. заменить собственный ребенок =
. cond(motherm[_N] == `i', mchild[_N], fchild[_N])
. если человек == `i' & ischild[_N] ;
. # разделитель кр
. }
По мере прохождения цикла forvalues локальный макрос i варьируется от 1 до максимально наблюдаемых человека , который мы подбираем как р(макс.) . Здесь мы извлекаем выгоду из того, что человека принимает небольшие целые числа от 1 и выше в пределах каждого семейства. Позже мы
посмотрите на метод сопоставления произвольных идентификаторов с этой настройкой. Что может
выглядеть как частный случай – это шаг в сторону от любой схемы идентификатора.
Продолжайте, пока мы начинаем цикл с `i’ , а также человек равно 1. Члены каждой семьи являются детьми этого человек если он или она их отец или их мать. форваль заменители 1 для `i’ :
. заменить ischild = (fatherm == 1) | (мама == 1)
Эта индикаторная переменная будет равна 0 (не является потомком 1) или 1 (является потомком 1). Для получения более подробной информации о том, какие переменные индикатора показывают истинное или ложное значение, см. видеть http://www.stata.com/support/faqs/data-management/true-and-false/.
Внутри каждого семейства мы будем сортировать по этой переменной, чтобы все
дети 9 лет0122 человека 1 в конце каждой семьи. Тогда мы можем выбрать
увеличить количество детей из других переменных в последнем наблюдении,
с учетом условий, которые будут упомянуты в данный момент.
qui по семействам (ischild), сортировать: заменить собственный ребенок = cond(motherm[_N] == `i', mchild[_N], fchild[_N]) если человек == `i' & ischild[_N]
Это много информации в одном утверждении, и лучше всего брать ее по частям:
- qui по семействам (ischild), сортировка:
Мы собираемся сделать вместо отдельно по семействам (напомним, что семейство — идентификатор семейства). В каждой семье мы сортировать сначала по ischild так, чтобы все дети человек 1 идти до конца семьи. Как всегда, сортирует по наименьшему значению. во-первых, поэтому все значения 0 идут перед всеми значениями 1 для индикатора переменные, такие как ischild . Также мы делаем все это тихо , хотя это не принципиально.
- заменить ownchild = … if person == 1 & ischild[_N]
Мы собираемся заменить ownchild , но только для наблюдений с лицо равно 1 и только в том случае, если последним человеком в семье является ребенок этого человека .
Как и прежде, под по varlist : _N интерпретируется внутри каждой определенной группы
к список переменных . Следовательно, ischild[_N] — это значение для последнего
человек в каждой семье. ( ischild[_N] — это ярлык для ischild[_N] == 1 , так как они всегда дают один и тот же результат. За
подробнее см. только что процитированный FAQ.) - Чем мы собираемся заменить собственный ребенок ?
Условие ischild[_N] гарантирует, что мы будем только заменяют значения , когда последнее наблюдение в каждом семействе относится к
ребенок любого лица, для которого лицо равно 1. Если это лицо
мать, мы используем значение для mchild ; если нет, мы используем значение
для fchild :
условие(мать[_N] == `i’, mchild[_N], fchild[_N])
Мы прошли операции для человек равно 1. для значений автоматически повторяет их для других значений человека .
7. Преобразование произвольных идентификаторов в целые числа от 1 и выше
Мы видели, что для некоторых задач есть преимущество в использовании целых чисел. идентификаторы, которые начинаются от 1 и выше в каждой группе. Если такое идентификаторов не существует, их можно создать, как показано в разделе 5.
Что нуждается в большем объяснении, так это то, как сопоставить произвольные существующие идентификаторы с эта установка. Предположим, что идентификаторы были, скажем,
член семьи отецм матьм 1 1001 . . 1 1002 . . 1 1003 1001 1002 1 1004 1001 1002 1 1005 1001 1002 2 2001 . . 2 2002 . 2001 г. 2 2003 . 2002 г.
Во-первых, мы генерируем целых чисел от 1 и выше, как и раньше.
. по семье (человеку), сортировать: gen pid = _n
Нам нужно сопоставить отцов и отцов с согласованными идентификаторами. Мы инициализируем переменные, которые хотим
. gen байт fid = . . средний байт = .
Теперь наш основной цикл состоит в циклическом переборе значений pid , которые конструкция содержит целые числа от 1 и выше. Мы заменяем фид и mid по каждому значению:
. суммировать pid, только в виду
. qui forval i = 1 / `r(max)' {
. #разделитель ;
. по семейству: заменить fid = `i'
. если отцов == человек[`i'] & !missing(отец) ;
. по семейству: заменить mid = `i'
. if motherm == person[`i'] & !missing(motherm) ;
. # разделитель кр
. }
То есть, циклически перебирая все значения pid , мы также перебирая все значения лицо . Хотя пример набор данных содержит числовые идентификаторы для человека , отца и motherm , код достаточно общий, чтобы его можно было применять к строковым идентификаторам как Что ж.
Выполнение этого по семейству: охватывает случай, когда значение человек уникален для человека в семье, но также может быть
идентификатор другого человека в другой семье. То есть один человек может быть человек 1 в одной семье и в другом человеке тоже может быть человек 1, но в другой семье. В качестве альтернативы, если человек имеет уникальное значение
для каждого человека в наборе данных мы ничего не теряем, делая это под на: , за исключением того, что, возможно, это может быть немного медленнее в машинном времени.
Дополнительные условия & !missing(fatherm) и &
!missing(motherm) необходимы. Почему? В примере семья 1 имеет 5
члены и семья 2 имеет 3 члена. Когда цикл forval достигает 4,
мы используем условия , если отец == человек[4] и , если мать
== человек[4] . До по семейству: интерпретируется подписка внутри групп, определенных семейством , но нет 4-го наблюдения для семья 2. Stata оценивает человека[4] как пропавших без вести в этом
обстоятельство, но тогда у нас возникает проблема, заключающаяся в том, что любые значения отцов или отцов , которые отсутствуют, будут сопоставлены с 4.