Пропись цифра 4: Пропись математическая, цифра 4, бесплатно распечатать – Раскраски и прописи для девочек и мальчиков l Загадки l Стенгазеты, де…
Цифра 4 в стихах, загадках и картинках
Изучать цифры гораздо интереснее, если рядом присутствуют интересные картинки, загадки, стихи ребусы для детей. Как правило, мы учим с детьми какое-нибудь число довольно скучно: показываем, как это число выглядит, просим посчитать предметы и соотнести их количество с цифрой и т.д. Особенно актуально привлечь внимание ребенка, когда он идет в 1 класс. В результате ребенок быстро теряет интерес и разочаровывается в обучении. А почему бы ни попробовать его заинтересовать? В этом вам помогут веселые загадки про цифры, презентация, стихи и ребусы, которые можно скачать на сайте. Не забудьте, что вам может понадобиться пропись, ведь число нужно научиться изображать!
На что похожа цифра?
Чтобы привлечь внимание ребенка, можно показать малышу картинки с четверкой и спросить: на что похожа цифра 4. Когда мы учим «сухой» материал, желательно вызвать у ребенка образные, запоминающиеся ассоциации. На что может быть похожа цифра 4? На перевернутый стул, на плуг, на наконечник стрелы и т.д. Такое задание вполне подойдет для детей, которые посещают 1 класс.
На что может быть похожа цифра 4? На перевернутый стул, на плуг, на наконечник стрелы и т.д. Такое задание вполне подойдет для детей, которые посещают 1 класс.
Здесь же оговариваем, что существует не только арабская, но и римская IV. А на что похожа римская цифра? Так постепенно у малыша возникает яркая ассоциация, и он уже не спутает четверку с другой цифрой.
Помочь представить, на что похожа цифра, могут стихи, презентация, раскраска, загадки, ведь в них число представлено с помощью образов. Они и подскажут малышу ответ, стимулируют фантазию.
Здесь можно посмотреть презентацию про число и цифру 4.
Раскраска
Как изучать числа?
Если ваш малыш дошкольник или только пошел в 1 класс, чтобы изучить с ним какое-нибудь число, предложите ребусы, раскраски, пословицы, стихи, загадки и т. д. Чем больше видов деятельности будет привлечено, тем успешнее пойдет обучение.
д. Чем больше видов деятельности будет привлечено, тем успешнее пойдет обучение.
Стихи про цифры отлично помогают запоминанию. Можно использовать увлекательные стихи С.Маршака или других авторов. Стихи не только обучают ребенка, но и способствуют развитию чувства ритма. Если вы любите стихи, обязательно привлеките их к изучению цифр.
Еще один вид творчества – загадки. Именно загадки способствуют развитию сообразительности. Наша задача – подобрать загадки о цифре 4. Скачать такие загадки вы сможете на нашем сайте.
Еще один интересный вид деятельности – ребусы. Чтобы угадать цифру 4, малышу придется хорошенько подумать и составить название цифры из других частей слов. В этом и заключаются ребусы. Использовать ребусы можно, готовя урок в 1 класс, или в старшую группу центра развития.
Если мы учим цифру 4 и хотим удивить ребят, поможет презентация. Скачать ее можно на сайте. Презентация сделает урок ярким и интересным, если вы идете в 1 класс к детям.
Выучить число 4 поможет раскраска для детей. Раскраска не только учит, но и тренирует усидчивость и терпение.
Если учим цифру 4, для детей будут интересны пословицы. Они представляют собой назидательные высказывания. Пословицы часто содержат в себе цифры.
Собираясь к детям в 1 класс, не забудьте пропись. Именно пропись поможет научить детей писать. Скачать пропись можно на сайте. Пропись развивает хороший почерк: чем больше малыш будет тренироваться, используя пропись, тем лучше.
Английская пропись с цифрой 4.Оригами: учимся складывать цифру четыре из бумаги.
Итак, если мы учим с ребятами цифру 4, не забудьте подключить к урокам для детей наглядные пособия, творческие задания, ребусы, пословицы и т.д. Они отлично подойдут, если вы готовите занятие в 1 класс или для развивающего центра.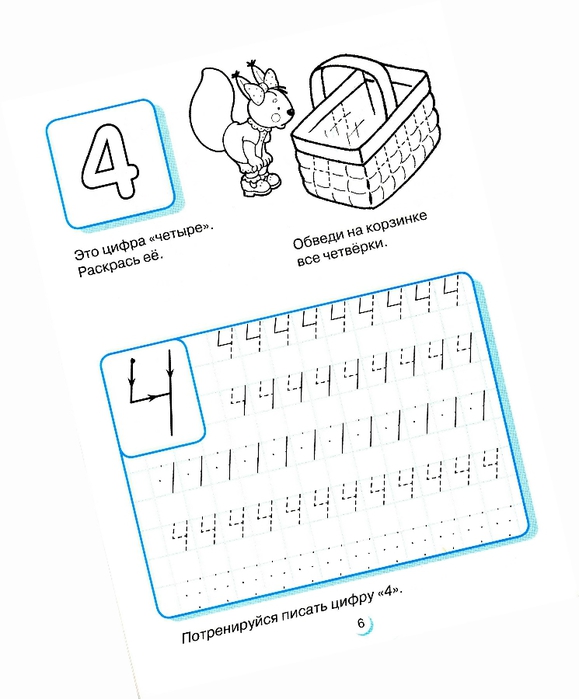
Учим детей с увлечением. Тогда и число 4 запомнится очень быстро!
Развивающие задания
Перед Вами задания о цифре 4.
Итак, что же нужно сделать?
- Посчитать книги (их будет 4).
- Найти среди множества других цифр цифры 4 и обвести в кружок.
- Посчитать бабочек, пчёл и черепах (их будет по 4).
- Провести линию и найти все цифры четыре в сердечках.
- Обвести цифру четыре начиная от стрелки.
- Подрисовать 4 жучка, которые попались в паутину к пауку.
- Найти на картинке с лодкой цифру 4 и раскрасить её.
Скачать задания о цифре 4 можно здесь.
Увлекательные задания про цифру четыре для детейОчень рекомендую добавить к заданиям о цифре 4 этот небольшой видео ролик из Арифметики малышки от Тётушки Совы.
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
| ||
Узнай стоимость своей работыБесплатная оценка заказа! |
 Огромный выбор интересных картинок.
Огромный выбор интересных картинок.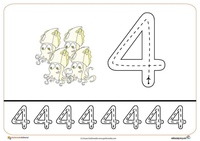

 Коллекция анекдотов регулярно пополняется, следите за обновлениями.
Коллекция анекдотов регулярно пополняется, следите за обновлениями.
Прописи цифры – Распечатать для детей
Здесь вы можете скачать прописи цифры, распечатать их на принтере и использовать в домашнем обучении для подготовки детей к школе. Мы подготовили для вас несколько заданий-прописей в картинках, благодаря которым ваши дети быстро научатся выводить цифры от 1 до 10, а заодно и потренируются считать, выполняя наши нехитрые задания. Владение письмом – один из важнейших навыков образованного человека.
Прописи цифры – Распечатать задания с мультяшными героями
Чтобы приступить к занятиям с ребенком, сначала вам нужно прописи цифры распечатать на принтере, подготовить цветные карандаши и шариковую ручку. Ручкой ребенок будет обводить цифры, а карандашами – раскрашивать мультипликационных героев после выведения всех цифр на листе. Все цифры-прописи находятся на отдельных листах. Всего – 10 листов и, соответственно, 10 цифр.
Очень важно при занятиях с прописями рассчитывать нагрузку в соответствии с возрастом детей. Чем младше ребенок, тем сложнее ему выводить ручкой линии в связи с недостаточно сформировавшимися фалангами пальцев. Поэтому ни в коем случае нельзя допускать переутомления ребенка. Проводить занятия с дошкольниками рекомендуется с небольшими интервалами, давая пальчикам отдохнуть.
Чтобы максимально эффективно использовать в обучении прописи цифры – совмещайте разные задания в одном уроке. Не дайте ребенку заскучать над однотипными упражнениями, подогревая его интерес рисованием, раскрашиванием и выполнением различных заданий, которые мы выложили на этой странице.
Чтобы прописи цифры распечатать, сначала скачайте файлы во вложениях внизу страницы
Прописи – Цифра 1
Напиши по образцу прописью цифру 1. Сколько коровок ты видишь на картинке?
Прописи – Цифра 2
Напиши по образцу прописью цифру 2. Узнал ли ты этих героев? Посчитай, сколько их?
Прописи – Цифра 3
Напиши по образцу прописью цифру 3. Сколько котят ты видишь на картинке?
Прописи – Цифра 4
Руководствуясь образцом напиши прописью цифру 4. Посчитай всех героев мультика на картинке.
Посчитай всех героев мультика на картинке.
Прописи – Цифра 5
Напиши прописью цифру 5, глядя на образец. Посчитай героев мультика про Винни Пуха. Назови их.
Прописи – Цифра 6
Напиши прописью цифру 6 – смотри на образце направление стрелочек. Посчитай героев мультфильма “Простоквашино”. Назови их.
Прописи – Цифра 7
Напиши по образцу прописью цифру 7 (обводи по направлению стрелочек). Посчитай героев сказки про Белоснежку. Назови их имена.
Прописи – Цифра 8
Выведи прописью цифру 8. Посчитай героев мультика “Смешарики”. Вспомни и назови их имена.
Прописи – Цифра 9
Глядя на образец, напиши прописью цифру 9. Обращай внимание на направление стрелок. Посчитай героев сказки про Русалочку. Раскрась картинку.
Прописи – Цифра 10
Напиши прописью цифру 10, как написано в образце. Посчитай героев мультика про пчелу Майю. Вспомни, как их зовут.
Пропись “Цифры для детей” – Задания в картинках
Также мы подготовили для вас еще одну пропись – Цифры для детей – с заданиями в картинках, где каждая цифра представлена на отдельном листе. На каждом листе-задании ребенку предстоит не только обвести несколько раз подряд конкретную цифру, но и решить несколько математических задачек в картинках – посчитать, обвести, раскрасить и т.д. Все задания предназначены для освоения и закрепления навыков счета и вместе с этим приучают ребенка к аккуратному и красивому почерку.
На каждом листе-задании ребенку предстоит не только обвести несколько раз подряд конкретную цифру, но и решить несколько математических задачек в картинках – посчитать, обвести, раскрасить и т.д. Все задания предназначены для освоения и закрепления навыков счета и вместе с этим приучают ребенка к аккуратному и красивому почерку.
С детьми до 5 лет рекомендуется проходить не больше одного задания в день. То есть 1 день – 1 цифра. Так вы сохраните здоровый интерес ребенка к обучению.
Скачать пропись “Цифры для детей” с заданиями в картинках вы можете во вложениях внизу страницы
Изучаем цифру 0 – Ноль.
Нужно раскрасить любым цветом цифру 0 и огрызок яблока. Обведи прописи цифры 0. Написать в пустых клетках цифру, которая соответствует количеству прямоугольников в каждой картинке.
Учим цифру 1 – Один.
Раскрась цифру и животное сверху. Обведи прописи цифры 1. Напиши в пустых клетках количество грибов в каждой картинке.
Учим цифру 2 – Два
Раскрась цифру и два носка на картинках сверху. Обведи прописи цифры 2. Впиши в пустые клетки цифры, соответствующие количеству мячиков на картинках.
Обведи прописи цифры 2. Впиши в пустые клетки цифры, соответствующие количеству мячиков на картинках.
Учим цифру 3 – Три
Раскрась картинки. Обведи все цифры 3. Выполни задание
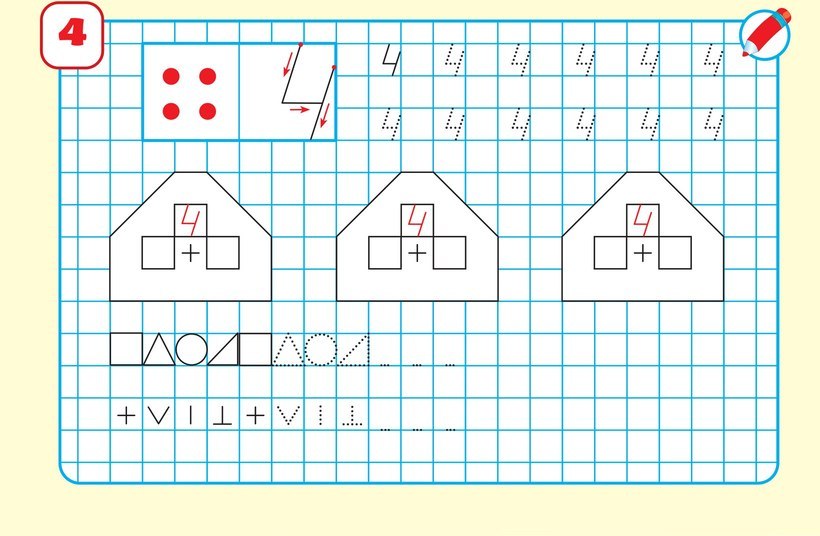
Учим и пишем цифру 4 – Четыре
Раскрась картинки сверху. Обведи прописи цифры 4. Посчитай и напиши в пустых клетках, сколько звездочек в каждой картинке.
Учим и обводим цифру 5 – Пять
Раскрась на картинках сверху цифру и всех сорок. Обведи прописи цифры 5. Посчитай и напиши в пустых клетках, какое количество чашек в каждой картинке.
Учим и пишем цифру 6 – Шесть
Раскрась на картинках сверху цифру и божьих коровок. Обведи прописи цифры 6. Посчитай и напиши в пустых клетках количество желудей в каждой картинке.
Учим и обводим цифру 7 – Семь
Раскрась на картинках сверху цифру и бабочек. Обведи прописи цифры 7. Посчитай и напиши в пустых клетках, сколько кружков в каждой картинке.
Учим и пишем цифру 8 – Восемь
Раскрась на картинках сверху цифру и цветы-тюльпаны. Обведи прописи цифры 8. Посчитай и напиши в пустых клеточках количество вишенок в каждой картинке.
Обведи прописи цифры 8. Посчитай и напиши в пустых клеточках количество вишенок в каждой картинке.
Учимся писать и считать с цифрой 9 – Девять
Раскрась на картинках сверху цифру и пальмы. Обведи прописи цифры 9. Посчитай количество звездочек в квадратиках и впиши соответствующее число в пустые клетки.
Учим и пишем цифру 10 – Десять
Раскрась сверху на картинках цифру и утят. Обведи прописи цифры 9. Посчитай и напиши, какое количество апельсинов в каждой клетке.
Прописи для детей 5, 6 лет – Цифры с заданиями в картинках
Далее мы подготовили для вас раскраски прописи для детей 5, 6 лет – Цифры с заданиями в картинках. Дошкольникам такие занятия не только очень полезны, но и крайне необходимы. Ведь к школе у ребенка должна быть хорошо развита мелкая моторика рук и привита аккуратность в письме. Да и считать до 10 ребенку нужно тренироваться постоянно, особенно последний год перед поступлением в школу.
Посмотри на картинку. Это цифра 1. Обведи ее по пунктирной линии и ты поймешь, как ее писать. Начинай обводить цифру сверху (с начала крючка).
Это цифра 1. Обведи ее по пунктирной линии и ты поймешь, как ее писать. Начинай обводить цифру сверху (с начала крючка).
Посмотри на цифру 2. Обведи ее по пунктиру в направлении стрелочки. Обведи все двойки и раскрась картинки.
А вот какая цифра 3. Если ее положить на бок, то на ней могли бы уместиться несколько птичек. Посчитай, сколько? Обведи по пунктиру цифру 3. Сколько рыбок плавает в аквариуме? Вспомни, в каких сказках упоминается число три (“Три орешка для Золушки”, “Три поросенка”, “Три медведя”, “Три богатыря”, “Три Ивана” и т.д.)
Посчитай, сколько ягод на картинке? Сколько цветов? Обведи все прописи цифры 3. Раскрась картинки.
Выполни задания.
Сейчас мы поговорим о цифре 4.
Посчитай и дорисуй.
А теперь познакомимся с цифрой 5. Обведи по контуру большую цифру, а также остальные прописи цифры 5. Посчитай цыплят и цветочки ландыша. Раскрась картинки.
Посчитай груши. Обведи прописи.
Обведи прописи.
Посчитай рыбок.
Загадка.
Чтоб не мерзнуть, пять ребят
По чуланчикам сидят.
Найди отгадку на картинке.
Посчитай, сколько кустиков клубники и грибочков на картинке. Впиши полученные числа в соответствующие квадратики.
На что похожа цифра 6? Обведи все прописи цифры 6 и раскрась замок.
Ты пригласил в гости на 6 часов пятерых друзей. Назови их имена. Сколько кружек чая ты должен налить, учитывая, что ты тоже будешь пить чай вместе с гостями? Обведи прописи.
Объедини фрукты в группы, как на рисунке. И сосчитай, сколько получилось групп у лимонов, яблок и клубники.
Найди на картинке цифру 6. Посмотри внимательно и найди на картинке предметы, напоминающие по форме цифру 6. Посчитай их количество.
Посмотри на цифру 7. Назови 7 цветов радуги. А теперь 7 дней недели. Вспомни название сказок, в которых присутствует цифра 7 (“Волк и семеро козлят”, “Белоснежка и семь гномов”). Обведи прописи. Какие предметы по форме похожи на цифру 7? Раскрась радугу в 7 цветов.
Обведи прописи. Какие предметы по форме похожи на цифру 7? Раскрась радугу в 7 цветов.
А теперь цифра 8. Какие предметы похожи на эту цифру? Обведи прописи цифры 8. Раскрась картинки.
Посчитай уток. Обведи цифры.
Посмотри на цифру 9. Обведи прописи.
Выполни задания
А вот и цифра 0. Что она тебе напоминает? Обведи прописи.
Обведи все прописи с цифрой 10.
Посчитай всех птиц на картинке. Посчитай цыплят.
Посчитай все звездочки и кошек.
Скачать прописи для детей 5, 6 лет – Цифры – вы можете во вложениях ниже
Веселые прописи – Сосчитай и обведи
И напоследок еще одни прописи для детей 5, 6 лет – Цифры для написания и счета с дополнительными заданиями и раскрасками. В первом задании нужно посчитать карандаши, раскрасить их разными цветами и обвести их по точкам. Во втором задании необходимо посчитать и раскрасить все предметы, а также обвести под каждой картинкой цифру, соответствующую количеству предметов.
Скачать – Веселые прописи со счетом – вы можете во вложениях внизу страницы
Кроме этого вам будут полезны и другие материалы для распечатки:
Прописи для детей – Буквы от А до Я
Если вы уже начали заниматься с прописями, то вам обязательно пригодятся прописи-буквы для дошкольников и учеников 1 класса.
Устный счет в пределах 10 – Картинки с заданиями
Здесь мы подготовили для вас устный счет в пределах 10 в виде математических заданий в картинках. Данные задания формируют у детей навыки счета и способствуют более эффективному обучению простых математических действий.
Раскраски с заданиями на счет в пределах 10
Чтобы дети могли быстро и с интересом освоить счет в пределах 10, мы подготовили для вас веселые раскраски с заданиями. Каждое задание содержит в себе картинки для раскрашивания – это стимулирует ребенка правильно выполнить задание.
Задания с раскрасками – Порядковый счет до 10
В этих интересных заданиях дети узнают, что такое порядковый счет до 10. А те, кто уже знакомы с этим понятием, могут показать свои знания с помощью данного упражнения.
А те, кто уже знакомы с этим понятием, могут показать свои знания с помощью данного упражнения.
Счет до 5 – Картинки с заданиями для малышей
Здесь мы выложили для вас счет до 5 – картинки с математическими заданиями для малышей, благодаря которым ваши дети потренируют не только свои навыки счета, но и умение читать, писать, различать геометрические фигуры, рисовать и раскрашивать.
А также развивающие игры для изучения чисел и тренировки навыков счета:
Развивающая игра – Посчитай картинки – Счет до 10
В этой игре малыш должен посчитать количество предметов на игровом экране и нажать на соответствующее число. После этого он увидит и услышит порядковый счет до данного числа.
Игра – Найди все числа по порядку и вразброс
Здесь ребенку нужно быть внимательным, чтобы найти все спрятанные числа на картинке. В игре также используется порядковый счет.
Игра – Найди самое большое и самое маленькое число
В этой игре ребенку необходимо выбрать среди предложенных чисел самое большое или самое маленькое. После правильного ответа он услышит порядковый счет до этого числа.
После правильного ответа он услышит порядковый счет до этого числа.
Прописи цифры. Учимся писать цифры – задания для детей – Развитие ребенка
Детские прописи цифр – учимся писать цифры
Гармоничное развитие ребенка представляет огромную важность для заботливых родителей, поэтому в раннем возрасте следует обучить его красивому написанию букв и цифр. Как известно, во многих современных школах уделяют недостаточно времени каллиграфии, поэтому дома обязательно следует проводить самостоятельные дополнительные занятия. Если Вы не обладаете навыками профессионального педагога – не беда, ведь прямо на нашем сайте Childdevelop Вы сможете найти и скачать детские прописи цифр, благодаря которым обучение станет не только полезным, но и занимательным!
Бытует мнение о том, что научить малыша писать – несколько сложнее, чем научить читать. Красивый почерк можно сравнить с истинным искусством, поэтому очень важно, чтобы ребенок освоил каллиграфическое мастерство в раннем возрасте.
Посетив наш сайт, вы сможете прописи цифры распечатать бесплатно и легко использовать их в практических занятиях с малышом. Мы предлагаем вашему вниманию самые интересные и разнообразные подборки, в которых представлены прописи цифр для дошкольников. Мы уверены, что они сразу же привлекут внимание вашего ребенка и пробудят в нем неподдельный интерес к обучающему процессу.
Прописи с цифрами для детей
Всем родителям известно, что малышей невозможно заставить что-либо делать против их воли. Особенно, если речь идет об обучении. Поэтому задача родителей – превратить этот процесс в игру, которая сможет увлечь маленьких непосед.
У нас Вы сможете найти прописи для малышей цифры, скачать и распечатать их для практических занятий. Без сомнения, ребенку будет интересно, ведь все цифры прописи сопровождаются увлекательными заданиями, которые способствуют правильному интеллектуальному развитию. Например, требуется написанные прописью цифры раскрасить в разные цвета, или дорисовать на имеющемся изображении элементы в виде той или иной цифры – иголки у ежика в виде единичек, рожки у улиток в виде двоечек, чешуйки у рыбок в виде троечек и т.д. Наши прописи с цифрами, скачанные с сайта и распечатанные на бумаге, помогут ребенку приобрести не только начальные математические знания, но и определенные представления об окружающем мире в целом.
Учимся писать цифры прописи
Предлагаемые нами прописи цифры для детей приносят значительную пользу не только для умственной деятельности, но и для мышц кистей рук. Используя прописи для детей цифры, малыш одновременно получает начальные навыки письма и тренирует руку – в результате таких занятий он очень быстро научится писать уверенным и красивым почерком.
Абсолютно бесплатно вы можете прописи цифры скачать у нас на сайте в разделе практических заданий. Учимся писать цифры прописи – ведь это так увлекательно!
Цифры на английском 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 20, 30, 50, 100, 1000. Английский счет, транскрипция и произношение.
Изучите как записываются прописью английские цифры 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 20, 30, 50, 100, 1000, 1000000 и научитесь счету по-английски.
В аудиокурсе VoxBook множество примеров использования цифр и числительных. Так в романе Роберта Луиса Стивенсона ‘Остров сокровищ’ из знаменитой песни:
Fifteen men on the dead man’s chest [пятнадцать человек на сундук мертвеца]
Yo-ho-ho, and a bottle of rum [йо-хо-хо, и бутылка рома]!
Drink and the devil had done for the rest [пей и дьявол тебя доведет до конца]
Yo-ho-ho, and a bottle of rum!(R.L. Stevenson – “TREASURE ISLAND”)
Ниже приведены приведены цифры на английском языке с транскрипцией и переводом. Далее подробно разобраны способы образования количественных числительных (Cardinal Numerals) и английский счет.
| Цифра | Название по- |
Название по- |
Транскрипция |
|---|---|---|---|
| 1 | один | one | [wʌn] |
| 2 | два | two | [ˈtuː] |
| 3 | три | three | [θriː] |
| 4 | четыре | four | [fɔː] |
| 5 | пять | five | [faɪv] |
| 6 | шесть | six | [sɪks] |
| 7 | семь | seven | [ˈsevn̩] |
| 8 | восемь | eight | [eɪt] |
| 9 | девять | nine | [naɪn] |
| 10 | десять | ten | [ten] |
| 11 | одиннадцать | eleven | [ɪˈlevn̩] |
| 12 | двенадцать | twelve | [twelv] |
| 20 | двадцать | twenty | [ˈtwenti] |
| 30 | тридцать | thirty | [ˈθɜːti] |
| 40 | сорок | forty | [ˈfɔːti] |
| 50 | пятьдесят | fifty | [ˈfɪfti] |
| 60 | шестьдесят | sixty | [ˈsɪksti] |
| 70 | семьдесят | seventy | [ˈsevnti] |
| 80 | восемьдесят | eighty | [ˈeɪti] |
| 90 | девяносто | ninety | [ˈnaɪnti] |
| 100 | сто | a (one) hundred | [ə wʌn ˈhʌndrəd] |
| 1000 | тысяча | a (one) thousand | [ə wʌn ˈθaʊzn̩d] |
| 1000000 | миллион | million / a (one) million | [ə (wʌn) ˈmɪlɪən] |
| 1000000000 | миллиард | milliard / a (one) milliard ( BrE ) billion / a (one) billion ( AmE ) |
[ə (wʌn) ˈmɪlɪɑːd] [ə (wʌn) ˈbɪlɪəŋ] |
Цифры по-английски 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12.
Цифры в английском языке от 1 до 12 являются простыми количественными числительными т.е. обозначают количество предметов и состоят из одного слова (см. Таблицу). Написание английских цифр 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 необходимо запомнить. На основе их первого десятка образуются все остальные английские цифры.
Цифры по-английски от 13 до 19.
В английском языке цифры 13, 14, 15, 16, 17, 18 и 19 являются производными количественными числительными.
Для их образования к английской цифре от 1 до 10 из третьего столбца таблицы добавляется суффикс -teen. В итоге получается цифра, необходимая для счета от 13 до 19:
13 тринадцать – thirteen [ˌθɜːˈtiːn]
14 четырнадцать – fourteen [ˌfɔːˈtiːn]
15 пятнадцать – fifteen [ˌfɪfˈtiːn]
16 шестнадцать – sixteen [sɪkˈstiːn]
17 семнадцать – seventeen [ˌsevnˈtiːn]
18 восемнадцать – eighteen [ˌeɪˈtiːn]
19 девятнадцать – nineteen [ˌnaɪnˈtiːn]
3 three – 13 thirteen
5 five – 15 fifteen
Ударение числительных с суффиксом -teen.
В английском языке числительные, оканчивающиеся на суффикс -teen, имеют два ударения, на первом и на втором слоге (ˌ слабое вторичное(побочное) ударение и ˈ основное ударение). Посмотрите транскрипцию, чтобы не ошибиться в произношении:
13 – thirteen [ˌθɜːˈtiːn]
14 – fourteen [ˌfɔːˈtiːn]
15 – fifteen [ˌfɪfˈtiːn]
Если за числительным с суффиксом -teen не следует существительного, то при произношении основное ударение падает на суффикс -teen:
fifteen [fɪfˈtiːn]
sixteen [sɪksˈtiːn]
Когда числительное с суффиксом -teen являются определением существительного (т.е. за за ним следует существительное), тогда ударение в нем падает не на суффикс, а только на его первый слог:
fifteen pencils [ˈfɪftiːn ˈpensl̩z]
sixteen boxes [ˈsɪkstiːn ˈbɒksɪz]
В аудиокурсе VoxBook в сказке ‘The Fish and the Ring’ из сборника English Fairy Tales , в котором можно услышать подобное ударение (для этого установите аудиокурс на свой компьютер и послушайте сами):
…and she lived there [и она жила там] till she was fifteen years old [пока ей не стало пятнадцать лет], and a fine handsome girl [и (стала) прекрасной красивой девушкой].(English Fairy Tales – “Fish and the Ring”)
Английские цифры 20, 30, 40, 50, 60, 70, 80, 90.
Английские цифры обозначающие целые десятки 20, 30, 40, 50, 60, 70, 80, 90, являются производными количественными числительными. Для их образования к цифре десятков из третьего столбца таблицы добавляется суффикс -ty. В итоге получается необходимое количество десятков:
20 двадцать – twenty [ˈtwenti]
30 тридцать – thirty [ˈθɜːti]
40 сорок – forty [ˈfɔːti]
50 пятьдесят – fifty [ˈfɪfti]
60 шестьдесят – sixty [ˈsɪksti]
70 семьдесят – seventy [ˈsevnti]
80 восемьдесят – eighty [ˈeɪti]
90 девяносто – ninety [ˈnaɪnti]
Обратите внимание, что в английских цифрах 20, 30, 40 и 50, корень от 2 two, 3 three, 4 four и 5 five видоизменяются, а в цифре 80 изчезает повтор буквы t:
2 two – 20 twenty [ˈtwenti]
3 three – 30 thirty [ˈθɜːti]
4 four – 40 forty [ˈfɔːti]
5 five – 50 fifty [ˈfɪfti]
8 eight – 80 eighty [ˈeɪti]
Ударение числительных с суффиксом -ty.
В английских числительных, оканчивающиехся на суффикс -ty ударение приходится только на первый слог. Посмотрите транскрипцию и произношение:
40 – forty [ˈfɔːti]
50 – fifty [ˈfɪfti]
60 – sixty [ˈsɪksti]
Английские цифры 100, 1000, 1000000.
Английские цифры от 100 до 1000 и 1000000 представлены в конце таблицы (см. выше).
В английском языке перед 100 hundred, 1000 thousand, 1000000 million ставится или неопределенный артикль а ( который имеет значение one – один), или ставится слово one:
100 сто – a(one) hundred [ə wʌn ˈhʌndrəd] (т.е. a hundred или one hundred)
1000 тысяча – a(one) thousand [a(one) thousand]
Обратите внимание, что с остальными количественными числительными артикль не употребляется:
Three pencils. Три карандаша.
Two girls. Две девушки.
В 100 hundred, 1000 thousand и 1000000 million не ставится окончания -s, в том числе когда перед ними стоит другое число отличное от единицы, например:
100 сто (одна сотня) – one hundred или a hundred
200 двести – two hundred
300 триста – three hundred
400 четыреста – four hundred
500 пятьсот – five hundred
600 шестьсот – six hundred
700 семьсот – seven hundred
800 восемьсот – eight hundred
900 девятьсот – nine hundred
далее
1000 тысяча (одна тысяча) – one thousand или a thousand
2000 две тысячи – two thousand
3000 три тысячи – three thousand
4000 четыре тысячи – four thousand
5000 пять тысяч – five thousand
и т.д.
1000000 миллион (один миллион) – one million или a million
2000000 два миллиона – two million
3000000 три миллиона – three million
4000000 четыре миллиона – four million
5000000 пять миллионов – five million
6000000 шесть миллионов – six million
и т.д.
Таким образом числительному никогда не ставят окончание -s, однако…
Однако: слово означающее число может относится не только к числительну (которому ставить окончание -s не надо), но и к существительному, у которого уже может быть окончание -s.
Например в 100 hundred, 1000 thousand и 1000000 million ставят окончание -s, когда они выражают неопределенное количество сотен, тысяч или миллионов. В этом случае они являются существительными, а следующее за ними существительное (если оно есть) используется с предлогом of :
Three millions of tons of coal. Три миллиона тонн угля.
Hundreds of ideas for learning. Сотни идей для обучения.
Some birds can fly thousands of kilometers. Некоторые птицы перелетают за тысячи километров.
Four or five millions of specie had been lately thrown into circulation. Четыре, пять миллионов звонкой монеты (за) последнее время были вброшены в оборот.
Все сказаное выше относится и к другим английским цифрам от 2 и выше, которые выступают в качестве существительного:
Magnificent seven. Великолепная семерка. (единственное число)
How many sevens your have? Сколько семерок у тебя? (множественное число)
В аудиокурсе VoxBook в романе Роберта Луиса Стивенсона ‘Остров сокровищ’ , с таким примером:
Here it is about gentlemen of fortune [так всегда с джентльменами удачи = пиратами]. They lives rough and they risk swinging [они живут без удобств и рискуют попасть на виселицу], but they eat and drink like fighting cocks [но едят и пьют как короли: «бойцовые петухи»= шиковать], and when a cruise is done, why, it’s hundreds of pounds instead of hundreds of farthings in their pockets [и когда рейс завершен = после плавания, что же, у них в карманах сотни фунтов вместо сотен фартингов =грошей].(R.L. Stevenson – “TREASURE ISLAND”)
Примечание. Кроме того, в million может ставиться окончание -s, когда перед ним стоит two, three и т.д. при условии, что после него нет другого числа. В этом случае после million следует существительное с предлогом of:
Two millions of books. – Два миллиона книг.
Английский счет. Cоставные количественные числительные в английском языке.
При английском счете в составных числах от 20 до 99 (т.е состоящие из двух чисел – десятнов и единиц), между десятками и следующими за ними единицами ставится дефис (черточка):
20 – twenty [ˈtwenti]
21 – twenty-one [ˈtwenti wʌn]
22 – twenty-two [ˈtwenti ˈtuː]
и т.д.
30 – thirty [ˈθɜːti]
31 – thirty-one [ˈθɜːti wʌn]
32 – thirty-two [ˈθɜːti ˈtuː]
и т.д.
В английском счете с составными числами свыше 100 в пределах каждых трех разрядов перед десятками (а если их нет, то перед единицами) ставится союз and:
235 – two hundred and thirty-five
407 – four hundred and seven
2034 – two thousand and thirty-four
2004 – two thousand and four
5236 – five thousand two hundred and thirty-six
250129 – two hundred and fifty thousand, one hundred and twenty-nine
4221589 – four million two hundred and twenty-one thousand five hundred and eighty-nine
203000000 – two hundred and three million
В американском варианте английского союз and опускается, например: 235 — two hundred hirty-five.
Некоторые правила перевода числительных на английский язык :
1. Русский родительный падеж существительных после количественных числительных не следует переводить на английский язык существительным с предлогом of (распространенная ошибка):
В аудиокурсе VoxBook в сказке ‘JACK THE GIANT-KILLER’ из сборника English Fairy Tales , в котором встречается именно этот случай:
…he is a huge and monstrous giant [он громадный и чудовищный великан] with three heads [с тремя головами]; he’ll fight five hundred men in armour [он победит/поборет пятьсот людей в доспехах], and make them fly before him [и «сделает»= заставит их убежать перед собой].’(English Fairy Tales – “JACK THE GIANT-KILLER “)
2. Русское существительное в единственном числе после составных числительных, оканчивающихся на один (одна, одно), следует переводить на английский язык существительным во множественном числе:
двадцать один день – twenty one days
триста пятьдесят одна книга – three hundred and fifty-one books
3.
Количественные числительные могут стоять в качестве определения после определяемого слова.
Особенно это относится к случаям обозначений номеров страниц, параграфов, глав и частей книг, номеров домов, комнат, размеров одежды и обуви, номеров автобусов и имеет значение порядкового числительного, хотя и замещено количественным числительным:
part two – вторая часть
chapter seven – седьмая глава
Read lesson three. – Читайте третий урок.
Open your book on page nineteen. – Откройте ваши книги на девятнадцатой странице.
He live in apartment 12 (twelve). – Он живет в квартире 12.
4. Сочетания две мои сестры, пять ваших книг, двое его друзей и т.п. переводятся на английский язык two of my sisters, five of your books, two of his friends (а не: two my brothers, five your books, two his friends – частая ошибка).
Например в аудиокурсе VoxBook в романе Роберта Луиса Стивенсона ‘Остров сокровищ’ , именно с таким примером:
There were six of the buccaneers, all told [там находилось шестеро пиратов, «все подсчитано» = в общей сложности]; not another man was left alive [больше никто не остался в живых]. Five of them were on their feet, flushed and swollen [пятеро из них были на ногах, с покрасневшими и опухшими /лицами/],(R.L. Stevenson – “TREASURE ISLAND”)
Конспект урока «Цифра 4» по математике для 1 класса
Ткаченко М.В., учитель классов КРО
ГКС(К)ОУ “Курганская специальная (коррекционная)общеобразовательная
школа-интернат VI вида
Открытый урок математики
Тема: «Цифра 4».
Цель: познакомить с новой цифрой – 4.
Задачи: обр. – научить писать цифру 4;
познакомить с образованием числа 4;
научить соотносить число 4 с цифрой и предметной совокупностью
корр. – развитие мелкой моторики при письме,
развитие мышления при соотнесении цифры с числом;
развитие внимания при решении примеров.
восп. – выработка навыка самостоятельной работы;
выработка навыка аккуратного письма.
ХОД УРОКА
Деятельность учителя | Деятельность ученика | |
1.Орг. момент. | Мы друг к другу повернулись, Мило, дружно улыбнулись, И в хорошем настроении Начинаем наш урок. | Настраивается на занятие |
2. Проверка дом. задания. | Проверка правильности выполнения, исправление ошибок при их наличии | Слушает комментарий учителя, исправляет ошибки. |
3. Актуали-зация знаний. | Повторение знаний об изученных цифрах – 1,2,3, о знаках «- »,«+», «=» – Назови самую маленькую цифру? – Большую? – С какого числа начинается числовой ряд? – Какое число стоит перед 2,3? – Какая цифра ничего не обозначает? – Покажи знак, который используется в примерах на сложение? – При вычитании? Реши пример: 1+1=? 1+2=? 3-1=? | Показывает на раздаточном материале 1 3 1 1 0 + – 2 3 2 |
4. Сообще-ние темы | Знакомство с целью урока | Слушает учителя |
5. Физ. минутка | Зрительная гимнастика (Учитель наглядно показывает фигуры) Рисуй глазам треугольник. Теперь его переверни Вершиной вниз. И вновь глазами Ты по периметру веди. Рисуй восьмерку вертикально. Ты головой не крути, А лишь глазами осторожно Ты вдоль по линиям води. И на бочок ее клади. Теперь следи горизонтально, И в центре ты остановись. Зажмурься крепко, не ленись. Глаза открываем мы, наконец. Зарядка окончилась. Ты молодец! | Выполняет самостоятельно |
6. Изучение нового материала | Объяснение получения новой цифры на раздаточном материале – треугольники. – Был 1 треугольник. Положим еще 1 Сколько станет? – Прибавим еще 1. Сколько станет? – И еще 1? Получится 4. Это новое число. – Определим ее место в числовом ряду? Работа по картинке – с. 99 – Что нарисовано на картинке? – Во что играют мальчики? – Что такое хоккей? Сколько мальчиков? Сколько мальчиков в синей форме? – В красной? – Как записать? Работа по картинке с.100 – Сколько у мальчиков клюшек? – Сколько шлемов? – Сколько вратарей? – Сколько игроков? -Как получили число 4? – 4 это: – 4 это: – 4 это: Работа с палочками с.100 №1, 4 Объяснение практической значимости цифры 4? с.102 №12 Пропись цифры 4 (подробное объяснение написания) | 2 3 Выстраивает числовой ряд, определяет положение нового числа. Отвечает не вопросы Мальчики Хоккей Игра на льду с шайбой 4 2 2 2+2 (выкладывает из цифр) 4 4 1 3 3+1 1+3 2+2 Работает с палочками Слушает рассказ учителя Прописывает вместе с учителем в тетради |
7. Закрепле-ние | С. 101 №8,9 (закрепление количественных представлений и счетных навыков) С. 102 №11 (закрепление цветовых представлений) | Устно выполняет задание |
8. Подведе-ние итога, дом. задан. | Какую цифру изучали? Где она стоит в числовом ряду? Прописать цифру, решить примеры | 4 После 3 Слушает задание |
Здесь представлен конспект к уроку на тему «Цифра 4», который Вы можете бесплатно скачать на нашем сайте. Предмет конспекта: Математика (1 класс). Также здесь Вы можете найти дополнительные учебные материалы и презентации по данной теме, используя которые, Вы сможете еще больше заинтересовать аудиторию и преподнести еще больше полезной информации.
1.Мотивация (организационный) | Начинается урок, Он пойдет, ребята, впрок. Постарайтесь все понять Про числа с цифрами узнать. – Кто из вас готов отправиться за знаниями? – Проверьте все ли у вас для этого готово? Откройте учебник на с.52 и тетрадь на с.50. Отметьте данные страницы закладками. – Сегодня на уроке мы с вами узнаем новое и постараемся помочь друг другу, если в этом будет необходимость. – У Маши и Миши живет интересное животное. А какое, вы догадаетесь, отгадав загадку: У нее четыре лапки. Эти лапки цап-царапки. Пара чутких ушей. Она гроза для мышей. (Кошка) | – Мы все! Дети работают в учебнике и тетради. Переклеивают стикеры с предыдущих страниц. Слушают и отгадывают загадку. | 2. Введение в тему урока. Целеполагание. Открытие новых знаний Работа по учебнику, знакомство с обозначением числа 4 математическим знаком – цифрой 4 Работа в парах. Подготовка к письму цифры 4. Работа в тетради Знакомство с порядковым числительным «четвертый» | Ребята, откройте учебник на странице 52. Посмотрите внимательно на рисунок задания 1. – Сколько лап у кошки? У собаки? Сколько ножек у стула? – Ребята, как вы думаете, о каком числе и цифре пойдет речь сегодня на уроке»? – Определим цель урока. – Для чего мы должны это изучить? – Так как мы должны определить место числу 4 в натуральном ряду чисел, то как вы считаете, где должно стоять число 4? -Почему? – Что мы сейчас сделали? – Посмотрите в задании 2, как выглядит цифра 4. -Теперь мы можем легко узнать её среди других цифр. -Найдите среди данных знаков цифру 4 и поставьте на нее фишку -Сверьте свой ответ с ответом соседа. Если ваши ответы совпали – покажите это. (Руки соединяют вместе) -Сколько раз нам встретилась цифра 4? (три раза) – Вывод: число 4 обозначается цифрой 4. – Рассмотрите рисунки задания 3 – Договоритесь между собой, кто будет работать красными, а кто – синими фишками. Определите, какие предметы или их части похожи на цифру 4 ФИЗМИНУТКА Раз, два, три, четыре, пять! Все умеем мы считать, Отдыхать умеем тоже – Руки за спину положим, Голову поднимем выше И легко – легко подышим… Раз, два – выше голова. Три, четыре – руки шире. Пять, шесть – тихо сесть. Семь, восемь – лень отбросим. – Посмотрите на задание № 4. Сколько очков выпало на кубике? (4) – Найди рамку, поставь пальчик, как пишут цифру 4, обозначающую число 4. Делал Егорка С мамой уборку. Опрокинул стул В квартире, Стал похож он На четыре. – Сейчас мы будем с вами учиться писать цифру 4. Предлагаю вам открыть тетради. – (учитель по ходу объясняет написание цифры – начинаем писать наклонную линию от верхней стороны клетки, немного правее середины и ведем наклонно вниз, доводим до середины клетки и ведем горизонтальную линию вправо. От второй точки ведем наклонную линию вниз, доводим её до нижней стороны клетки, чуть правее её середины). – А сейчас я прочитаю отрывок из сказки «Про Козлёнка, который умел считать до десяти» (слайд) Каким по счёту Козлёнок назвал Быка? Сколько всего животных оказалось у озера? – Запиши в тетради с помощью цифр 0, 4, 2, 3, 1 числа по порядку – от самого маленького до самого большого. Назови число, следующее сразу за числом 3. | «Число и цифра 4» – Знать, что означает число 4; какой цифрой обозначается. -будем учиться писать цифру 4 – Находить место числа в числовом ряду, уметь сравнивать с другими числами После числа 3 -Оно больше 3 на 1 -Определили место числу 4 в натуральном ряду чисел. Проверка по слайду электронного учебника Работая в парах, выясняют, какие предметы или их части похожи на цифру 4 Показ по слайду электронного учебника. Работая самостоятельно, дети пишут сначала элементы цифры, а затем цифру 4. – дети на доске и в тетради выполняют задание. |
границ | Последствия вариации числа копий CYP2D6 для фармакогеномики в психиатрии
Введение
Генетическая вариация, реакция на лекарство и гены CYP
Клиническое применение геномных технологий для улучшения назначения и настройки планов фармацевтического лечения широко известно как фармакогеномика (PGx ). Основные принципы PGx: 1) генетическая изменчивость ключевых генов, участвующих в обработке и транспортировке фармакологических агентов и их метаболитов, может влиять на клинические исходы значимым, индивидуальным для пациента образом и 2) сходные генетические вариации в фармакологических мишенях могут влиять на индивидуальная чувствительность к действию определенных лекарств (1).Хотя большое количество генов было идентифицировано как важных участников PGx, наиболее клинически полезными и лучше всего изученными являются члены суперсемейства цитохромов P450. Эта большая группа из более чем 50 человеческих генов демонстрирует большое сходство в последовательности ДНК, хотя ее члены катализируют множество реакций. Клиническое значение имеет то, что несколько ферментов участвуют в пути детоксикации фазы I, включая хорошо известные локусы PGx, такие как CYP2C9 , CYP2C19 и CYP2D6 (2).
Вариация однонуклеотидного полиморфизма (SNP)
За последние два десятилетия результаты опубликованных исследований в фундаментальных научных исследованиях и клинически ориентированных журналах решительно поддерживают использование PGx в клинической практике (3, 4). Многие подробно описывают влияние конкретных вариантов, обычно однонуклеотидных полиморфизмов (SNP), в генах «CYP» на конечную функцию фермента. Эти так называемые орфографические ошибки в геноме приводят к ряду физиологических последствий, включая отсутствие изменений, измеренное снижение функции белка, полное стирание функции или даже повышение ферментативной активности.Иногда генетическая изменчивость включает небольшие вставки или делеции пар оснований вместо замены одной пары оснований на другую. Эта форма вариации известна как «INDEL», которая представляет собой набор слов «вставка» и «удаление». Их часто группируют вместе с заменяющими мутациями, такими как переходы от A к G или трансверсии от A к C, потому что они имеют тенденцию оказывать сравнимые и часто пагубные эффекты на функцию ферментов. Для большинства участков генома наследуются две копии – одна от матери и одна от отца, – которые вместе могут влиять на физиологию пациента.
Когда у пациента есть два аллеля пониженной функции, его ожидаемое физиологическое состояние в отношении метаболизма лекарственного средства этим ферментом может считаться «нормальным метаболизатором», NM для некоторых генов, включая CYP2D6 , как в случае недавнего генотипа CPIC проект стандартизации трансляции -фенотипов (5). Однако в литературе также есть исторические интерпретации этой комбинации как «промежуточный метаболизатор», IM или «слабый метаболизатор», PM, в зависимости от конкретного гена и конкретной комбинации присутствующих аллелей (6).Клинически снижение или устранение функции ферментов может способствовать изменению ответа на лекарственную терапию. В зависимости от конкретного фармацевтического субстрата, о котором идет речь, IM и PM могут не выводить стандартные дозы лекарств или их биоактивных метаболитов с нормальной скоростью, что приводит к повышенному риску побочных эффектов. Альтернативно, IM и PM могут быть неспособны активировать пролекарства, так что пациент может не в полной мере воспользоваться назначенной терапией.
Когда у пациента есть два аллеля повышенной функции или три или более функциональных копий гена данного интересующего гена, его ожидаемое физиологическое состояние называется «сверхбыстрым метаболизатором», UM.Когда один аллель повышенной функции сочетается с одним аллелем нормальной функции, общая скорость метаболизма называется «сверхбыстрым метаболизатором» или аналогичным «быстрым метаболизатором», RM. Клинически усиленный метаболизм лекарственного средства, доставляемого в его активной форме, может потребовать увеличения дозы или разделения более высокой дозы на несколько доз в день для достижения постоянных терапевтических уровней. Это связано с усилением метаболизма препарата в неактивную, выводимую форму. Для пролекарств может быть рекомендована альтернативная терапия или сниженная доза, поскольку ожидается большее, чем обычно, количество метаболизированного, циркулирующего активного соединения, что может привести к повышенному риску негативных побочных эффектов (7–9).
Варианты числа копий (CNV)
Другой тип генетических изменений с глубокими последствиями для PGx – это варианты числа копий или «CNV». В локусах, которые демонстрируют такие вариации, две или более копий одной и той же последовательности гена могут быть унаследованы от одного родителя или ген может быть полностью удален. Таким образом, общее количество открытых рамок считывания, доступных для продукции полнофункциональных, нарушенных или усиленных белков (в зависимости от структуры других присутствующих вариантов), может существенно отличаться от ожидаемого значения двух копий гена.В некоторых случаях общее количество копий может быть равно нулю или достигать 10 или более (10), что может присутствовать в различных возможных комбинациях (например, пять копий от каждого родителя, шесть копий от одного и четыре от другого и семь копии с одного и три с другого). С клинической точки зрения это дополнительное измерение генетической изменчивости может сильно повлиять на ожидаемую физиологию для данного набора наблюдений, часто внося значительную двусмысленность в процесс интерпретации результатов для конкретного пациента (см. Таблицу 1).Например, когда три или более дублированных аллелей демонстрируют нормальную функцию на основе их паттернов SNP и вариантов INDEL, может возникнуть состояние «сверхбыстрого метаболизма» – ожидается большее количество функционального белка через транскрипцию / трансляцию из более двух открытых рамок считывания. Хотя они могут показаться несколько экзотической формой вариации, CNV на самом деле довольно распространены – примерно 12% генома человека демонстрируют некоторую степень вариации числа копий (11), в том числе ключевые гены PGx, включая CYP2D6 (12 –15).Фактически, одно исследование показало, что 12,6% населения США в целом имели вариацию числа копий в гене CYP2D6 (16).
Таблица 1 Возможные последствия дублирования для интерпретации статуса метаболизатора пациента.
События преобразования гена
Кроме того, из-за высокой степени сходства последовательностей среди генов CYP наблюдались события преобразования генов с участием различных членов, включая CYP2D6 . В случае преобразования гена часть одной последовательности ДНК фактически наклеивается поверх некоторой части другой, создавая гибридный ген, содержащий последовательность из обоих локусов.В случае CYP2D6 было описано большое разнообразие таких гибридных аллелей между ним и близлежащим псевдогеном CYP2D7 (17–21). Клинически, как и многие изменения генома, такие перестройки часто приводят к снижению функции или нефункциональности белков (22).
Стратификация пациентов
Независимо от типа (ов) вариации, многие исследования PGx включают важную информацию о том, как пациенты с определенным статусом метаболизатора различаются по своей реакции на исследуемый терапевтический субстрат.Однако очень немногие из этих исследований содержат прямые и незамедлительные клинические выводы. Например, согласно широко цитируемой базе знаний PGx PharmGKB, только около 7% лекарств имеют связанную геномную информацию, на которую может воздействовать непосредственно врач (1). Интересно, однако, что эти соединения составляют около 18% от всех рецептов, выписанных в США (1). Кроме того, недавние сообщения указывают на то, что большая часть людей несут по крайней мере один вариант с действием PGx (23), причем многие несут два или более.Это говорит о том, что большая часть популяции пациентов демонстрирует, по крайней мере, некоторый потенциал столкнуться с лекарством, для которого доступна информация о PGx. Однако конечная клиническая полезность генетических данных пациента часто зависит от оценки и интерпретации полной комбинации вариантов, которыми они обладают, а не от наличия одного или двух вариантов. Таким образом, хотя очевидно, что генетическая стратификация пациентов может быть ценным подспорьем для практикующих врачей, заказ коммерчески доступных отчетов PGx следует рассматривать как улучшение, а не замену существующих стандартов лечения.
Потенциальные преимущества PGx для психиатрии
С клинической точки зрения, область психиатрии демонстрирует большой потенциал для получения непропорционально большей пользы от принятия PGx, чем другие специальности. Возможно, наиболее важной причиной является относительно высокий уровень неблагоприятных клинических исходов для пациентов, получающих стандартную помощь. Например, от 30% до 50% психиатрических пациентов не реагируют в достаточной степени на неотложное лечение, независимо от того, какое лекарство было первоначально прописано (24–32), и только 35–45% пациентов с большим депрессивным расстройством возвращаются к преморбидному уровню функции после 6-8 недель лечения (24).Таким образом, очевидно, что геномные данные могут использоваться в современной клинической практике. Кроме того, психиатрия является второй наиболее часто наблюдаемой первичной терапевтической областью (20,8%, см. Рисунок 1A) после онкологии (31,9%) в списке всех одобренных Управлением по контролю за продуктами и лекарствами США (FDA) лекарств с доступной информацией о PGx (33 ) (сведено в Таблицу 2). Итак, как и в случае с онкологией, исследовательская литература явно содержит исходный материал для создания ценной поддержки принятия клинических решений психиатрами. Наконец, из них примерно 20.8% одобренных FDA лекарств с информацией о PGx, которые используются в психиатрии, 69,2% полностью обрабатываются каким-либо образом одним геном: CYP2D6 (см. Рисунок 1B). Это представляет собой существенное обогащение субстратов CYP2D6 в психиатрических препаратах, поскольку в целом известно, что CYP2D6 влияет на метаболизм примерно 25% всех лекарств, одобренных FDA. Эти данные предлагают прекрасную возможность сосредоточиться на ценной области генома с большим потенциалом для улучшения результатов лечения пациентов. Таким образом, несмотря на биологические и технические проблемы измерения и интерпретации данных из CYP2D6 , локус может содержать ключ к важным улучшениям в стандарте оказания помощи психиатрическим пациентам.
Рисунок 1 Процент (A) одобренных FDA препаратов с доступной информацией о биомаркерах на этикетках по терапевтическим классам (всего N = 215) и (B) одобренных FDA психиатрических препаратов, на которые влияют определенные PGx представляющие интерес гены (всего N = 45, см. также Таблицу 2).
Таблица 2 45 лекарственных препаратов, одобренных FDA для использования в неврологии и психиатрии, которые содержат фармакогеномную информацию на этикетках лекарств и интересующий ген / биомаркер для каждого из них.В общей сложности 36 этикеток лекарственных средств содержат действенную или информативную фармакогеномную информацию в разделах «Дозировка и введение», «Предупреждения и меры предосторожности», «Побочные реакции», «Взаимодействие с лекарствами», «Клиническая фармакология» и «Использование в определенных группах населения» (33).
CYP2D6: структура, наблюдаемые вариации и номенклатура
Ген CYP2D6 состоит из девяти экзонов и находится на отрицательной цепи между 42 126 499 и 42 130 881 пн (GRCh48.p12) на хромосоме 22q13.2. Как показано на рисунке 2, вариация CYP2D6 происходит в экзонах, интронах, а также в областях, расположенных выше и ниже локуса.
Рис. 2. Структура от 5 ‘до 3’ локуса CYP2D6 и размещение вариантов. Физическое положение известных вариантов фармакогеномики (PGx) в CYP2D6 по rsID (всего N = 198). Цвета указывают на предполагаемые функциональные последствия: красный = отсутствие функции, синий = снижение функции, фиолетовый = неизвестная или неопределенная функция, черный = однонуклеотидный полиморфизм (SNP) с множественными связанными функциональными последствиями [e.g., rs1135840 можно найти в аллелях с нормальной функцией, пониженной функцией и даже нефункциональными (например, * 35 , * 17 и * 4 , соответственно)], зеленый = отсутствие наблюдаемых функциональных последствий на сегодняшний день (нормально). Числа справа от каждого rsID указывают общее количество гаплотипов (* аллелей), на которых, как известно, встречается каждый вариант.
На сегодняшний день каталогизировано 198 отдельных вариантов различных типов для CYP2D6 (18) (PharmVar версия 3.4, рисунок 2А). Чтобы прийти к полезной клинической интерпретации, подмножества, присутствующие у любого данного пациента, должны рассматриваться одновременно. Таким образом, концепция «гаплотипа» – обычно называемая «аллелями *» (читаемыми как «звездные аллели») в генах CYP – и соответствующие стандарты номенклатуры (22) для аллелей CYP2D6 имеют решающее значение для понимания. Здесь «гаплотип» относится к точной комбинации вариантов, обнаруженных на физической цепи ДНК, унаследованной от конкретного родителя. Комбинация двух гаплотипов, унаследованных от обоих родителей, в совокупности известна как «диплотип», и обычно принято перечислять гаплотип с наименьшим номером первым (34).В конечном счете, важно помнить, что аллели * – это гаплотипы CYP2D6 , которые могут включать несколько сайтов и типов вариаций.
Часто один и тот же SNP может быть обнаружен на нескольких генетических фонах, которые, исходя из совокупности имеющихся вариантов, демонстрируют разные активности (rsID выделены черным цветом на рисунке 2). Кроме того, многие названные / охарактеризованные коллекции вариантов определяются большим количеством генетических изменений (Рисунок 3 и дополнительные рисунки 1–4, см. * 35B ( CYP2D6 * 35.002 ), который содержит 38 вариантов, и * 2A ( CYP2D6 * 2.001 ), который несет 16), некоторые из которых являются общими для нескольких названных аллелей. Например, вариант C> T в экзоне 1, известный как rs1065852 (22: 42130692 в GRCh47, c.100C> T, p.Pro34Ser, P34S), присутствует по крайней мере в 36 различных гаплотипах (рис. 2). Таким образом, одна важная техническая задача для преобразования необработанных генетических данных в точное определение диплотипа состоит в том, чтобы понять, какие варианты были унаследованы вместе от каждого родителя.
Рис. 3 Физические положения (красные вертикальные линии) и общее количество вариантов, обнаруженных на 19 гаплотипах (* аллели), по прогнозам, продуцируют нормально функционирующие ферменты при транскрипции / трансляции. См. Также дополнительные рисунки 1–4. Серые прямоугольники обозначают экзоны (сборка генома 37).
После характеристики каждому уникальному наблюдаемому гаплотипу присваивается собственное обозначение «*», и он регистрируется в различных базах данных и общедоступных ресурсах (35, 36). В самых простых случаях один вариант полностью определяет гаплотип.Например, наличие буквы «C» в позиции rs5030867 в настоящее время – это все, что необходимо для идентификации гаплотипа * 7 в CYP2D6 . В более сложных случаях одновременное присутствие многих типов вариаций (например, SNP, CNV и событий генной конверсии) определяет гаплотипы, переносимые данным пациентом. Чтобы правильно идентифицировать такие сложные аллели *, необходимы измерения всех или почти всех участков вариации. Если ни на одном из тестируемых участков не наблюдается изменений, гаплотипу по умолчанию присваивается обозначение * 1 .
Когда наблюдается вариация числа копий для одного или нескольких гаплотипов, обозначение дублированного аллеля включается как « xN », где N – это количество копий определенного аллеля, когда это возможно определять. Например, диплотип CYP2D6 * 1×2 / * 2 указывает, что один аллель несет дупликацию гена * 1 , а другой аллель несет одну копию гена * 2 .
Гаплотипирование CYP2D6, анализы SNP и клиническая интерпретация: соображения и проблемы
Существует важная техническая проблема в разрешении определенных комбинаций гаплотипов, включающих гетерозиготные варианты в нескольких местах, которая не возникает, когда наблюдаемая вариация является гомозиготной.В частности, если два варианта наблюдаются в одном и том же месте (например, результат теста для определенного варианта является гомозиготным при отсутствии события дупликации или делеции целого гена), один должен происходить от одного родителя, а другой – от другого родителя. . Таким образом, схема наследования ясна. Однако, если два варианта наблюдаются в разных местах, неясно, произошли ли оба варианта от одного родителя или один вариант был унаследован от каждого. Последствия многовариантных генотипов особенно сложны, когда они включают изменения, полностью исключающие функцию ферментов.Пациенты, демонстрирующие эти комбинации, унаследовали либо две нефункциональные копии гена, либо одну нормальную копию, которая сочетается с одной нефункциональной копией, несущей оба варианта. Эти две возможности могут иметь очень разные физиологические интерпретации в зависимости от применяемого препарата и, следовательно, различных клинических последствий. Реальные ситуации могут быть значительно более сложными для решения, чем этот простой пример с двумя локусами, и у некоторых пациентов также могут наблюдаться новые аллели, что может значительно усложнить клиническую отчетность.Однозначное определение специфического распределения вариантов на каждой хромосоме дает «поэтапные» гаплотипы, то, что способны произвести очень немногие из современных технологий. Скорее, фаза обычно оценивается с использованием существующих знаний о гаплотипах, которые ожидаются в генофонде пациента и / или с помощью математических алгоритмов .
В настоящее время гаплотипы CYP2D6 , которые обеспечивают повышенную общую функцию, делают это посредством повышенной трансляции мРНК в белок из-за присутствия двух или более последовательностей генов, обеспечивающих нормальную функцию (гаплотипы с переменной CNV, см. Ниже и рисунок 4 ).Таким образом, они должны быть идентифицированы с помощью анализа, разработанного специально для целей их обнаружения. Аналогичная проблема возникает с полиморфизмами конверсии генов, которые производят гибридные аллели. Поскольку анализы различных типов вариаций, показывающих измеримые последствия для общей активности CYP2D6 у пациентов, обычно выполняются независимо, объединение необработанных данных из всех источников в целостную картину клинической применимости может быть довольно сложной задачей. Кроме того, поскольку эффекты конкретных комбинаций проявляют ряд физиологических эффектов от нефункционального белка CYP2D6 до повышенной скорости метаболизма фермента CYP2D6, клиническая интерпретация даже точных генетических профилей может быть трудной, а иногда и неоднозначной.
Рисунок 4 Последствия дупликации гена CYP2D6 для (A) уровень активности 13 ожидаемых CNV-переменных (дублированных) аллелей CYP2D6 с известной функцией фермента. Не показан: CYP2D6 * 43 , который, как известно, демонстрирует дупликации с неопределенным фенотипом. (B) Индивидуальная активность гаплотипа (* аллель) и общий статус метаболизатора для конкретного результата пациента « * 1 / * 4 , CNV = 4». Обратите внимание, что число копий, равное четырем, вводит неоднозначность в сообщаемом статусе метаболизатора из-за технической неопределенности относительно того, какой конкретный аллель дублируется.
Следует также отметить, что большое количество гаплотипов показывает либо неизвестные – комбинации, которые слишком редки или для которых имеется слишком мало опубликованных данных для эффективной интерпретации, – либо неопределенная функция, то есть результаты тестов / исследования, которые являются противоречивые или неубедительные. Диплотипы, включающие гаплотипы с неизвестными или неопределенными функциями, особенно трудно интерпретировать клинически полезными способами, хотя тематические исследования с их участием будут полезны для разрешения неоднозначности.Например, изучение случая не отвечающего пациента, который несет известный гаплотип с противоречивыми доказательствами в других исследованиях, но который явно получает пользу от изменения терапии, будет полезным наблюдением и предлагает потенциально плодотворные направления будущих исследований. Эти направления могут включать in vivo, фенотипирование или фармакокинетические исследования подобных лиц, которые проливают дополнительный свет на функцию рассматриваемого аллеля.
Когда диплотипы пациента включают два гаплотипа с четко определенными функциями, они могут быть сгруппированы в потенциально субстрат-специфические группы статуса метаболизатора, включая нормальный (NM, ранее называвшийся экстенсивным, EM), промежуточный (IM), плохой (PM) и сверхбыстрый ( UM).В зависимости от полного понимания клинических и фундаментальных научных исследований, ожидаемые физиологические последствия членства в каждой группе могут быть затем преобразованы в поддержку принятия клинических решений.
CYP2D6: гаплотипы с переменными CNV
Аллель CYP2D6 * 5 представляет собой полную делецию последовательности гена, которая может быть унаследована от одного или обоих родителей. Функционально, поскольку из аллеля * 5 нельзя продуцировать белок, он полностью устраняет ферментативную функцию CYP2D6.Следовательно, в физиологической интерпретации статуса метаболизатора его обычно рассматривают так же, как и другие аллели, лишенные функциональности, и он представляет собой крайний конец функционального континуума для всех субстратов. Однако существуют технические проблемы, которые возникают, когда присутствуют аллели * 5 . Например, когда * 5 сочетается с аллелем * 1 (т. Е. Не наблюдаются варианты SNP или INDEL), технические ограничения могут привести к тому, что пациент будет иметь два нормально функционирующих аллеля ( * 1 / * 1 ), пока не будет измерен статус номера копии (37).Таким образом, в отсутствие данных CNV точная клиническая интерпретация результатов может оказаться невозможной даже для примеров, которые кажутся относительно простыми с генетической точки зрения. Подобные осложнения возникают, когда * 5 сочетается с аллелями, несущими различные комбинации вариантов – технические результаты будут выглядеть так, как будто пациент гомозиготен по всем наблюдаемым вариантам, а не гетерозиготен вместе с аллелем * 5 . Это, в свою очередь, может внести значительную двусмысленность в клиническую интерпретацию.
Так же, как могут быть оптовые делеции аллеля CYP2D6 , хромосомная область, в которой обнаружен CYP2D6 , может нести две или более копии гена CYP2D6 . Такие дупликации могут включать генные единицы, которые являются функциональными (например, * 1xN и * 2xN ) или нефункциональными (например, * 4xN ) или имеют пониженную функцию (например, * 41xN ), ведущие к множеству потенциальных клинических последствий. На сегодняшний день было обнаружено, что 14 гаплотипов, несущих различные комбинации SNP и INDEL, дублируются у одного или нескольких человек (14, 38) (Рисунок 4A).Чаще всего встречаются дублированные аллели * 1 , * 2 и * 4 (15). В то время как другие дупликации появляются реже, они действительно встречаются с заметной частотой в клинических популяциях и, следовательно, должны учитываться при определении комбинаций диплотипов и сообщении об их клинических последствиях. Следует отметить, что, поскольку текущие каталоги геномных вариаций человека являются неполными, особенно для популяций неевропейского происхождения, другие дупликации, включающие известные и пока еще недокументированные аллели, вероятно, существуют где-то в человеческой популяции.По мере того, как базы данных и другие геномные ресурсы улучшаются в их выборке глобально разнообразных популяций с течением времени, ожидается, что повысится как точность диплотипирования, так и точность соответствующей поддержки принятия клинических решений.
Анализы CYP2D6 CNV: соображения и проблемы
Хотя техническая чувствительность лабораторных анализов на CNVs на CYP2D6 может варьироваться, некоторые из них способны точно определить общее количество присутствующих до 5 и даже 10 копий. Однако многие анализы могут обеспечить разрешение CNV только> 2, и в большинстве случаев разрешение становится менее определенным при уровнях> 4.Важно отметить, что, как отмечалось выше, анализы CNV и SNP / INDEL часто выполняются независимо, а доступные базы данных / каталоги вариаций являются неполными. Таким образом, может быть трудно определить, какой аллель * должен быть назначен какому копийному номеру. Например, для пациента, для которого был проведен анализ как CYP2D6 * 1 / * 2 с числом копий 4, в полностью описательном клиническом отчете результаты должны быть представлены как « * 1×3 / * 2 или * 1×2 / * 2×2. или * 1 / * 2×3 », потому что гаплотип дублированного гена не был определен с уверенностью.К счастью, в этом случае физиологическая интерпретация статуса метаболизатора и, следовательно, клинические последствия идентичны для всех трех возможностей. В частности, поскольку и * 1 , и * 2 показывают нормальную функцию, каждый потенциальный результат дает одно и то же обозначение статуса сверхбыстрого метаболизатора (рис. 4A, нижние поля: черный, зеленый и синий). Аналогичным образом, пациент CYP2D6 * 4 / * 6 с любым значением числа копий может быть интерпретирован как PM, поскольку независимо от того, сколько копий любого аллеля присутствует, ожидается, что весь продуцируемый белок будет нефункциональным для конкретный представляющий интерес субстрат (рис. 4A, верхние красные прямоугольники).
Ситуация становится все более сложной, когда присутствуют дублированные аллели с различными функциональными характеристиками . Например, у пациента CYP2D6 * 1 / * 4 с CNV = 4 (рис. 4B) снова есть три различных возможности для их диплотипа: * 1×3 / * 4 , * 1×2 / * 4×2 и * 1 / * 4×3 . Однако в этом случае каждый из них связан с отдельным статусом метаболизатора (UM, NM и IM, соответственно), и поэтому каждый может иметь различную клиническую интерпретацию (например,g., увеличенная доза, стандартная доза или уменьшенная доза лекарственного средства, доставленного в его активной форме). Такие неоднозначные результаты следует интерпретировать с осторожностью и с учетом истории болезни конкретного пациента и реакции на лекарства (если таковая имеется). Предыдущие побочные реакции на лекарственные препараты и эффективность лекарств в прошлом могут пролить или не пролить свет на фактический имеющийся статус диплотипа и метаболизатора. Такое сочетание генетического тестирования и традиционных клинических подходов к лечению, вероятно, представляет собой лучший сценарий для определенных генетически сложных результатов.
Сверхбыстрые метаболизаторы CYP2D6 в клинической практике
Измерение общего числа копий в локусе CYP2D6 особенно важно для PGx в клинической психиатрии. В настоящее время единственный известный способ увеличения метаболизма CYP2D6 – это за счет дупликации одного или нескольких аллелей переменной CNV * с нормальной или пониженной функцией. Кроме того, наиболее полезная поддержка принятия клинических решений на основе PGx для многих препаратов, используемых в психиатрии, может быть предоставлена для сверхбыстрых метаболизаторов, которые, по определению, должны нести один или несколько дублированных аллелей.
Несмотря на отсутствие всеобъемлющих руководств по использованию статуса метаболизатора в клинической практике, убедительные научно-обоснованные исследования и данные о результатах подтверждают их полезность во многих контекстах. Например, Центр оценки и исследований лекарственных средств FDA США разрешает и утверждает добавление информации о метаболическом статусе и дозировке, а также предупреждений непосредственно на этикетках лекарств. Точно так же независимые фармакогеномные консорциумы включили в руководящие принципы действенную информацию о PGx. Например, у Голландской рабочей группы по фармакогенетике (DPWG) есть 47 руководств (39), а у Консорциума по внедрению клинической фармакогенетики (CPIC) есть 19 рецензируемых и опубликованных руководств по 40 парам ген-лекарство (40).В этих рекомендациях обычно подчеркивается, что сверхбыстрый метаболизм CYP2D6 важен для лечения пациентов.
Действительно, давно известно, что дополнительные функциональные копии CYP2D6 влияют на фармакокинетику различных субстратов, включая нортриптилин и дебризохин (38, 41). Было показано, что скорость выведения нортриптилина была в пять раз выше, чем у PM, у субъектов, несущих только одну дополнительную копию CYP2D6 , то есть три полные копии (38).Когда присутствовало 13 функциональных копий, частота была в 17 раз выше, чем у субъектов без активного фермента CYP2D6. Кроме того, впоследствии было показано, что ингибирование хинидином метаболизма дебризохина у лиц, несущих 3, 4 или 13 нормальных копий CYP2D6 , может быть использовано для потенциально изменения клинических исходов. Эти данные вместе предполагают, что предварительное знание статуса метаболизатора пациента на CYP2D6 с помощью генетического тестирования может быть неоценимым для избежания таких проблем, как резистентность к лечению и / или токсичность (42).
Подобная информация может существенно повлиять на выбор терапии и дозировки в различных контекстах, поскольку CYP2D6 обрабатывает несколько клинически ценных анксиолитиков, антидепрессантов и нейролептиков (см. Таблицу 2). Например, метаанализ сообщает, что доза ~ 50% обычно используемых нейролептиков зависит от генотипа CYP2D6 (24). Кроме того, чрезвычайно высокие скорости клиренса антидепрессанта тримипрамина наблюдались у трех носителей дупликаций CYP2D6 , взятых у группы здоровых субъектов.В конечном итоге авторы предполагают, что таким людям может потребоваться доза до 200% от средней для достижения концентраций, аналогичных нормальным метаболизаторам (43). Кроме того, поскольку CYP2D6 отвечает за гидроксилирование тримипрамина, а не за его метилирование (44), необходимо также учитывать влияние статуса CYP2D6 UM на активный метаболит десметилтримипрамин, поскольку он также метаболизируется CYP2D6. Таким образом, потенциальная разница в клинических эффектах может быть даже больше, чем при рассмотрении только тримипрамина (43).
В том же исследовании изучалось влияние дупликации CYP2D6 на трициклический антидепрессант доксепин у здоровых субъектов. Авторы заметили, что сверхбыстрые метаболизаторы показали уровни активного метаболита N -десметилдоксепина на уровне 40% от уровней, наблюдаемых у нормальных метаболизаторов, и, учитывая как это активное дочернее соединение, так и уровни самого доксепина, НМ показали уровни в два раза выше, чем UM . Аналогичным образом интерпретируемые тенденции наблюдаются для трициклических антидепрессантов имипрамин и дезипрамин (10, 45).
В ретроспективном исследовании невосприимчивости к антидепрессантам, метаболизируемым CYP2D6, полное отсутствие UM наблюдалось в подгруппе пациентов ( N = 28), у которых наблюдались нежелательные лекарственные явления (ADE), в то время как восемь были идентифицированы как PM. Авторы приходят к выводу, что это обогащение в четыре раза выше, чем ожидалось случайно. И наоборот, в подгруппе из 16 неответчиков без ADE не наблюдались UM и только один PM (10). Хотя размер выборки, по общему признанию, невелик, эти тенденции убедительно подтверждают роль дупликаций CYP2D6 в важных показателях клинических исходов.Наконец, в исследовании, которое выявило 81 человека, не ответившего на антидепрессанты, метаболизируемые CYP2D6, 10% (восемь субъектов) имели дупликации. Авторы отмечают, что это существенное обогащение по сравнению с 0,8–1,0% заболеваемости, ожидаемой для северных европеоидов, и что оценки наихудшей недели по шкале оценки депрессии Гамильтона были выше у тех, у кого были дублированные аллели, чем у тех, кто этого не сделал (10, 46 ).
Клиренс миртазапина в форме S (+) демонстрирует аналогичную картину для разных групп метаболизаторов и, как было установлено, равен 1.В 6 раз больше в UM CYP2D6, чем в NMs (47). Поскольку форма R (-) не метаболизируется CYP2D6 (48, 49), но, по-видимому, отвечает за сердечно-сосудистые НЯ, UM могут иметь более высокий риск как терапевтической неудачи, так и побочных эффектов при назначении высоких доз миртазапина (10 , 47). Кроме того, терапевтическая неудача из-за эффектов повышенного метаболизма CYP2D6 может усилить суицидальное поведение у пациентов с депрессией (50–52). Более того, было обнаружено, что UM имеют повышенный риск получения высоких баллов по одной из шкал оценки депрессии Гамильтона, которая измеряет суицидальность среди стационарных пациентов с униполярной или биполярной депрессией (53).UMs также могут показывать более низкие концентрации флуоксетина или амитриптилина в плазме при монотерапии, чем PM, IM и NM, если начальные концентрации находятся на нижнем конце диапазона при лечении большого депрессивного расстройства (54).
В одном из крупнейших долгосрочных слепых рандомизированных контролируемых исследований [Геномика, используемая для улучшения принятия решений о депрессии (GUIDED)], в котором участвовали 1167 амбулаторных пациентов с диагнозом большого депрессивного расстройства (БДР) и неадекватным ответом пациента или клинициста на по крайней мере, одним антидепрессантом, было обнаружено, что лечение под контролем фармакогенетических тестов по сравнению с лечением как обычно на 8 неделе показало статистически значимое улучшение ответа (26.0% против 19,9%) и ремиссии (15,3% против 10,1%). Эти результаты дополнительно подтверждают потенциальную роль фармакогеномного тестирования в управляемом лечении трудно поддающихся лечению психиатрических пациентов и в улучшении ответа и показателей ремиссии (55).
Было высказано предположение, что сверхбыстрый метаболизм CYP2D6 взаимодействует с другими генетическими факторами, влияя на реакцию на лечение у определенных групп пациентов. Например, недавняя статья предполагает, что ремиссия венлафаксина-XR чаще встречается у пациентов с большим депрессивным расстройством, которые: 1) не ответили на циталопрам / эсциталопрам, 2) имели сверхбыстрый метаболизм CYP2D6 и 3) имели определенные комбинации аллелей SLC6A4 . и SLC6A2 (56).Этот потенциально более высокий уровень генетического разрешения для поддержки принятия клинических решений предполагает, что в будущем могут быть возможны более точные и точные уровни специфичности для конкретных групп пациентов. В некоторых случаях это может включать не только взаимодействия, обусловленные метаболизмом соединений множеством генов на определенной стадии процессинга, но также действие одного и того же гена на разных стадиях детоксикации.
Обсуждение
Биология, технология, интерпретация и поддержка принятия клинических решений
Локус CYP2D6 показывает очень сложный паттерн генетических вариантов, которые наследуются во множестве комбинаций.Эффект любой данной комбинации вариантов на транслируемый белок также может значительно варьироваться. В свою очередь, клинические результаты, измеренные у пациентов, несущих аналогичные функциональные аллели, также демонстрируют некоторую степень вариабельности, но также и достаточную статистическую согласованность, чтобы показать большие перспективы для добавления нового понимания в уход за пациентами и улучшения стандартной практики. Однако для того, чтобы воспользоваться этими преимуществами, необходимо преодолеть технические проблемы, связанные с точным сбором и интерпретацией необработанных лабораторных данных, тем, кто пытается оказывать поддержку в принятии клинических решений на их основе.К ним относятся: 1) точное получение данных для каждого SNP / INDEL, конверсии гена и варианта числа копий, 2) их систематизация в вероятные гаплотипы, 3) определение статуса метаболизатора, который, вероятно, будет передаваться каждой комбинацией, и 4) точное соединение каждого из них. статус к самой последней информации о клинической безопасности и эффективности в постоянно меняющемся ландшафте первичной литературы. Последним шагом, бременем медицинского работника, является, конечно, интеграция этой информации в планы лечения таким образом, чтобы это принесло пользу пациентам в клинических сценариях.
Точный анализ CNV в локусе CYP2D6 особенно важен для обеспечения максимальной клинической пользы от тестирования. Без этой ключевой информации во многих случаях результатам можно приписать очень мало уверенности. К сожалению, в то время как одобренные FDA методы анализа вариабельности CYP2D6 (например, AmpliChip CYP450 от Roche и xTAG CYP2D6 от Luminex) принимают во внимание CNV, некоторые лабораторные тесты (LDT) не учитывают (57). Действительно, в несколько тревожной недавней публикации, посвященной отчетам о PGx, было отмечено, что почти треть обследованных лабораторий, по-видимому, не смогли включить CNV в тестирование CYP2D6 (58).Более того, поскольку методы измерения SNP и INDEL обычно выполняются независимо от методов измерения CNV, почти все тесты ограничены в их способности объединять эти многочисленные источники данных в окончательные гаплотипы (т.е. ген) (15, 59). Как мы также видели, наличие аллельной делеции (аллель CYP2D6 * 5 ) вызывает осложнения как с технической точки зрения, так и с точки зрения отчетности.Однако, хотя современные лабораторные методы, вероятно, несовершенны, они, тем не менее, дают клинически полезные сведения в целом с большим потенциалом для поддержки создания планов лечения для конкретных пациентов. Фактически, было подсчитано, что затраты на лечение «экстремальных» метаболизаторов при CYP2D6 (либо PM, либо UM), по сравнению с NM, могут достигать от 4000 до 6000 долларов больше в год (60, 61), и, таким образом, новые знания статуса метаболизатора пациента может помочь как снизить затраты, так и повысить качество лечения.
Передовой опыт
В конечном счете, существует несколько рекомендаций по передовому опыту использования PGx в психиатрии. Во-первых, как и в случае любого клинического анализа, важно выбрать клинические фармакогеномные тесты, которые проводятся в лаборатории, аккредитованной в соответствии с поправками к клинической лаборатории (CLIA). Это лаборатории, которые проводят клиническое тестирование на людях, а не генетическое тестирование в исследовательских целях. Они обязаны соблюдать установленный набор правил и положений и регулярно проверяются.Во-вторых, выберите анализ, который измеряет разумное подмножество CYP2D6, SNP, INDEL и CNV. Анализы, выполняемые для обнаружения вариации SNP / INDEL, должны быть способны улавливать ключевые * аллели, подходящие для соответствующей популяции пациентов. Что касается CNV, анализы должны быть способны определять количественные различия в общем количестве аллелей как минимум между 0 и 5 копиями. В-третьих, для целей отчетности и интерпретации влияние всех измеренных вариаций на функцию ферментов следует рассматривать одновременно как гаплотипы, а не как один вариант за раз.Это означает, что результаты CYP450 следует сообщать как диплотипы (пары * аллелей), а не только наличие и / или отсутствие конкретных индивидуальных вариантов. Кроме того, следует сообщать обо всех возможных результатах неоднозначных комбинаций, когда они возникают (например, * 1×3 / * 2 , или * 1×2 / * 2×2 , или * 1 / * 2×3 ).
Наконец, при рассмотрении результатов результаты должны быть помещены в их надлежащий клинический контекст и оценены за клиническую полезность, которую они могут или не могут представлять.Например, такие результаты, как « CYP2D6 * 1 / * 1 , CNV = 2» (т. Е. Отсутствие наблюдаемых отклонений в рассматриваемом анализе), не просто «не результаты», а скорее указывают на то, что стандартные меры предосторожности и процедуры уместно с признанием того, что могут присутствовать неизмеряемые варианты, что является клинически полезным выводом. Кроме того, отчеты, показывающие неоднозначные результаты – с точки зрения так называемых диплотипов, статуса метаболизатора или потенциального клинического воздействия наблюдаемых вариантов у данного пациента – могут не обеспечивать четких клинических действий, но могут быть полезны в контексте истории болезни пациента и другие клинические факторы при определении тактики медикаментозной терапии.Например, генетика может указывать на то, что пациент может быть либо UM, либо NM при CYP2D6, но побочные реакции в его медицинской карте могут убедительно свидетельствовать о том, что один статус более вероятен, чем другой. Такие случаи представляют собой важные тематические исследования, которые следует рассмотреть для публикации. Существуют и другие уникальные клинические сценарии, которые усложняют интерпретацию генетических результатов. Например, ожидается, что реципиенты трансплантата печени будут иметь сложные профили обработки лекарств, которые напоминают статус метаболизатора донора, а не статус, определяемый собственной ДНК пациента.Ясно, что такие факторы выходят за рамки лабораторного тестирования, основанного на крови, слюне или замене щеки, и, таким образом, поставщик обязан интегрировать всю соответствующую информацию в общую картину ухода за пациентом.
Клиническая значимость популяционных эффектов и
CYP2D6 * 1 / * 1Частоты определенных вариантов, включая некоторые ключевые дублированные аллели, показывают нетривиальные уровни вариабельности среди глобальных популяций (62). Это наблюдение имеет несколько клинически значимых эффектов.Первое и наиболее важное заключается в том, что обозначение * 1 в настоящее время наиболее подходит для популяций европейского происхождения и может вводить в заблуждение лиц, недавних предков из других континентальных популяций. Текущие геномные базы данных всех типов, включая те, которые широко используются для разработки тестов PGx, сильно не отражают глобальное геномное разнообразие. Таким образом, многие SNP, INDEL и CNV с потенциально важным влиянием на функцию ферментов и клинические исходы просто не каталогизируются и поэтому не появляются в лабораторных анализах CYP2D6 .Таким образом, пока базы знаний и другие ресурсы не станут более полными, следует проявлять определенную осторожность при интерпретации результатов * 1 / * 1 .
Кроме того, различия в частоте аллелей среди континентальных популяций, вероятно, влияют на широкий спектр локусов в масштабах всего генома. Поскольку детоксикация фазы I – сложный процесс, в котором задействовано множество ферментов, который интегрирован со многими другими биологическими системами, ожидается некоторое количество вариаций в эффектах локусов PGx.Например, если процессинг CYP2D6 обычно является этапом, ограничивающим скорость метаболизма конкретного лекарственного средства, но другой локус отвечает за процессинг биоактивного метаболита, изменения частот аллелей во втором локусе вполне могут изменить клинически наблюдаемые эффекты измеренных вариантов CYP2D6 . Таким образом, любой конкретный пациент среднего европейского происхождения может демонстрировать больший или меньший эффект, связанный с CYP2D6 для данной комбинации вариантов, чем ожидается, исходя из потенциально измеримых вариаций в других локусах, влияющих на общую физиологию пациента.
Будущее тестирования PGx и клинического руководства
Несмотря на общепризнанные ограничения, тестирование PGx сейчас явно клинически полезно. Это особенно верно в отношении психиатрической помощи. В качестве усовершенствования современной практики и важного источника понимания физиологии пациента и ожидаемого ответа на лекарства PGx может помочь обеспечить максимально эффективные и минимально рискованные планы лечения, улучшить результаты лечения пациентов и внести свой вклад в столь необходимую эффективность расходов на здравоохранение. Интересно, что эти преимущества, как ожидается, только увеличатся, учитывая: 1) большой объем качественных исследований, публикуемых ежегодно, 2) поддержание и курирование критически важных баз знаний, ориентированных на агрегирование ключевых результатов, 3) разработку нескольких коммерческих продуктов, обеспечивающих как академические, так и научные исследования. участие отрасли в этой области, 4) растущее применение PGx в клинической практике и 5) окончательное появление четких практических руководств.Более широкое применение PGx и инструментов поддержки принятия решений по рецепту (63) в повседневной практике особенно важно, поскольку оно предоставит неоценимые возможности для определения и уточнения плодотворных гипотез и целевых исследовательских инициатив, которые напрямую связывают исследовательские усилия с конечными точками и результатами, имеющими очевидную клиническую важность. В частности, развивающейся полезности и расширению области применения PGx в ближайшем будущем будут способствовать разработки в трех ключевых областях: 1) преодоление текущих технологических ограничений, 2) необходимость в четких данных о результатах для каждой комбинации лекарства / диплотипа, и 3) использование исследований PGx и связанных с ними баз данных для изучения генетического влияния на метаболизм эндогенных соединений и ксенобиотиков в целом.
Как объяснялось выше, в настоящее время не существует единой технологии, которая могла бы точно, эффективно и одновременно анализировать все критические типы генетических вариаций и однозначно связывать их с конкретной молекулой ДНК, унаследованной от конкретного родителя. Это включает в себя большинство стратегий для полногеномного секвенирования, которые также не могут надежно произвести такого рода «полностью поэтапные» геномные данные. Тем не менее, в разработке находятся многообещающие методы, которые выходят за рамки этого обзора для подробного изучения, такие как методы долгого чтения, одномолекулярного секвенирования (64), которые потенциально предлагают такой уровень разрешения генома.Кроме того, способность однозначно определять местоположение каждого варианта в каждой копии гена, присутствующего у конкретного пациента, окажет глубокое влияние на наше понимание клинического воздействия гаплотипов с переменным CNV. Как только проблемы, связанные с этими подходами, будут решены, появится возможность изучить полную картину геномных вариаций в ключевых локусах, таких как CYP2D6 , от пациента к пациенту и, таким образом, более точно разместить их в клинически значимых группах в соответствии ожидаемая функция фермента.В сочетании с улучшенными подходами к физиологическому тестированию для понимания процесса детоксикации, этот повышенный уровень детализации, несомненно, позволит выявить определенные подгруппы пациентов, которым непропорционально выгодно или которые подвергаются непропорциональному риску при применении определенных терапевтических стратегий.
Однако эта соответствующая фенотипическая группировка пациентов (например, по статусу метаболизатора) также зависит от дальнейшего уточнения нашего понимания клинического воздействия конкретных, потенциально редких комбинаций вариантов.Таким образом, для будущего развития отчетности PGx также будет критически важно, чтобы данные о ключевых результатах продолжали собираться в нескольких представляющих интерес популяциях на основе диплотипа за диплотипом. Улучшение нашего более широкого понимания полезности геномных вариаций, обеспечиваемое такими усилиями, несомненно, позволит перейти от интерпретаций PGx с «информативных» на текущих этикетках FDA к «действенным». Это также повысит уверенность в отчетности для групп населения, которые в настоящее время недостаточно изучены, и, вероятно, позволит получить более подробную информацию о дозах (например, доступную для арипипразола) для большего числа фармацевтических препаратов.В идеале эта работа должна быть выполнена на больших выборках пациентов-людей, хотя это может быть не полностью выполнимо из-за практических ограничений, включая очень большое количество потенциальных смешивающих переменных. В качестве альтернативы, при изучении и установлении «истинных» градаций пониженной функции между нефункциональной и нормальной, может быть полезно дополнить исследования на людях работой на культуре клеток или модельных организмах, чтобы получить достаточные размеры выборки и статистическую поддержку для физиологически значимых Выводы.
Наконец, расширение области применения PGx из генетики процессинга / транспорта и клинических эффектов фармацевтически продаваемых соединений в другие области биохимии и фармакокинетики также явно не за горами. Например, CYP2D6 был идентифицирован как потенциальный метаболизатор эндогенных нейроактивных субстратов (65), что предполагает будущее применение точных данных из CYP2D6 в психиатрической помощи, связанной с внутренними гомеостатическими процессами / физиологией в «нормальном» диапазоне и естественным прогрессированием заболевания.Кроме того, генетическая архитектура, отвечающая за переработку коммерчески производимых соединений, та же самая, что отвечает за метаболизм ксенобиотиков в более общем плане. Таким образом, повторное использование полногеномных данных PGx для понимания последствий непреднамеренного воздействия на окружающую среду химических веществ, таких как перфторалкил и полифторалкильные вещества (PFAS) и многие другие вероятные нейротоксины, вероятно, станет важным фактором будущего. здоровье и благополучие населения в целом.
Выводы
Использование PGx в качестве расширения к текущему стандарту ухода за психиатрическими пациентами показывает большой потенциал для направления терапии и улучшения результатов в самых разных клинических контекстах. Однако это не следует рассматривать как панацею. Следует иметь в виду важные ограничения, как технические, так и биологические, и информацию о PGx следует тщательно интегрировать с другими данными, специфичными для пациента, при разработке индивидуальных планов лечения. Некоторые из этих ограничений будут продолжать давать неоднозначные результаты для некоторых пациентов в обозримом будущем в таких локусах, как CYP2D6 , особенно при наличии CNV.Однако многие неоднозначные результаты по-прежнему предоставляют полезную и полезную информацию, если они полностью объяснены и поняты. Аналогичным образом, * 1 / * 1 может также предоставить полезные клинические рекомендации в поддержку стандартного курса лечения. Однако их также следует рассматривать с осторожностью в зависимости от объема вариантов, протестированных в данном отчете (т. Е. Чем больше, тем лучше), и от исходной популяции некоторых пациентов (например, мы должны помнить, что базы знаний в настоящее время неполны для неевропейского происхождения).Таким образом, последующее тестирование с расширенными анализами, повторное тестирование в какой-то момент в будущем или инвестирование в постоянно обновляемые клинические интерпретирующие отчеты могут быть полезны в определенных обстоятельствах.
Также важно следить за развитием PGx в ближайшие годы. Ожидается, что технологические достижения повысят полезность геномных данных в клинике, а постоянно расширяющиеся базы данных клинических результатов, вероятно, улучшат и расширит поддержку клинических решений, которую можно предоставить.В конечном счете, PGx является ценным инструментом в арсенале любого клинициста, и его разумное использование для уточнения планов лечения для конкретного пациента может значительно улучшить здоровье и благополучие многих психиатрических пациентов.
Вклад авторов
Все авторы внесли свой вклад в исследование, написание рукописей и исправления, а также прочитали и одобрили представленную версию.
Финансирование
Эта работа была поддержана Coriell Life Sciences.
Заявление о конфликте интересов
Авторы JJ, AP и JS работали в компании Coriell Life Sciences.JJ и JS имеют долю в Coriell Life Sciences.
Благодарности
Мы благодарим Пола Р. Шамана, DDS, MS, а также рецензентов, чьи предложения помогли улучшить и прояснить эту рукопись.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fpsyt.2019.00432/full#supplementary-material
Ссылки
2. Iyanagi T • Молекулярный механизм ферментов, метаболизирующих лекарства фазы I и фазы II: значение для детоксикации. Int Rev Cytol (2007) 260: 35–112. doi: 10.1016 / S0074-7696 (06) 60002-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
5. Консорциум по внедрению клинической фармакогенетики (CPIC). Проект стандартизации от генотипа к фенотипу CYP2D6. (2019). По состоянию на 9 мая 2019 г.
Google Scholar
7. Ahmed S, Zhou Z, Zhou J, Chen SQ. Фармакогеномика ферментов и переносчиков метаболизирующих лекарств: актуальность для точной медицины. Геномика Протеомика Биоинформатика (2016) 14: 298–313.doi: 10.1016 / j.gpb.2016.03.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
11. Ян И, Боттон М.Р., Скотт Э.Р., Скотт С.А. Секвенирование гена CYP2D6: от открытия вариантного аллеля до клинического фармакогенетического тестирования. Фармакогеномика (2017) 18: 673–85. doi: 10.2217 / pgs-2017-0033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
13. Дель Тредичи А.Л., Малхотра А., Дедек М., Эспин Ф., Роуч Д., Чжу Г.Д. и др. Частота аллелей CYP2D6, включая структурные варианты, в США. Front Pharmacol (2018) 9: 305. doi: 10.3389 / fphar.2018.00305
PubMed Аннотация | CrossRef Полный текст | Google Scholar
14. Gaedigk A, Ndjountche L, Divakaran K, Dianne Bradford L., Zineh I., Oberlander TF, et al. Гетерогенность локуса гена цитохрома P4502D6 (CYP2D6): характеристика случаев дупликации генов. Clin Pharmacol Ther (2007) 81: 242–51. doi: 10.1038 / sj.clpt.6100033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
17.Gaedigk A, Bradford LD, Alander SW, Leeder JS. Расположение генов CYP2D6 * 36 в локусе cyp2d6: ассоциация CYP2D6 * 36 с плохим метаболическим статусом. Drug Metab Dispos (2006) 34: 563–9. doi: 10.1124 / dmd.105.008292
PubMed Аннотация | CrossRef Полный текст | Google Scholar
18. Консорциум по изменению фармакогенов (PharmVar). Версия 3.4.
Google Scholar
19. Gaedigk A, Jaime LK, Bertino JS Jr., Berard A, Pratt VM, Bradfordand LD, et al.Идентификация новых гибридов CYP2D7–2D6: нефункциональные и функциональные варианты. Front Pharmacol (2010) 1: 121. doi: 10.3389 / fphar.2010.00121
PubMed Аннотация | CrossRef Полный текст | Google Scholar
21. Gaedigk A, Fuhr U, Johnson C, Berard LA, Bradford D, Leeder JS. Гибридные тандемы CYP2D7–2D6: идентификация новых механизмов дупликации CYP2D6 и их значение для предсказания фенотипа. Фармакогеномика (2010) 11: 43–53. DOI: 10.2217 / стр.09.133
PubMed Аннотация | CrossRef Полный текст | Google Scholar
22. Gaedigk A, Ingelman-Sundberg M, Miller NA, Leeder JS, Whirl-Carrillo M, Klein TE, et al. Консорциум по изменению фармакогена (PharmVar): включение базы данных номенклатуры аллелей цитохрома P450 (CYP) человека. Clin Pharmacol Ther (2018) 103: 399–401. doi: 10.1002 / cpt.910
PubMed Аннотация | CrossRef Полный текст | Google Scholar
23. Мостафа С., Киркпатрик CMJ, Байрон К., Шеффилд Л.Анализ частот аллелей, генотипов и фенотипов, действенных фармакогеномных (PGx) вариантов и феноконверсии у 5408 австралийских пациентов с генотипами CYP2D6, CYP2C19, CYP2C9 и VKORC1. J Neural Transm (Вена) (2019) 126: 5–18. doi: 10.1007 / s00702-018-1922-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
24. Kirchheiner J, Nickchen K, Bauer M, Wong ML, Licinio J, Roots I, et al. Фармакогенетика антидепрессантов и нейролептиков: вклад аллельных вариаций в фенотип лекарственного ответа. Mol Psychiatry (2004) 9: 442–73. doi: 10.1038 / sj.mp.4001494
PubMed Аннотация | CrossRef Полный текст | Google Scholar
25. Тема E, Стефанович М., Иванишевич AM, Блазинич Ф., Кулав Дж., Скочилич З. Генотипирование CYP2D6 у пациентов, получающих психоактивную лекарственную терапию. Clin Chem Lab Med (2000) 38: 921–7. doi: 10.1515 / CCLM.2000.135
PubMed Аннотация | CrossRef Полный текст | Google Scholar
26. Мерфи Г. М. мл., Кремер К., Родригес Х. Э., Шацберг А. Ф.Фармакогенетика непереносимости антидепрессантов. Am J Psychiatry (2003) 160: 1830–5. doi: 10.1176 / appi.ajp.160.10.1830
PubMed Аннотация | CrossRef Полный текст | Google Scholar
29. Фоли К.Ф., Куигли Д.И. Фармакогеномный потенциал психиатрических препаратов и CYP2D6. MLO Med Lab Obs (2010) 42: 32–4.
Google Scholar
30. Уолден Л. М., Брандл Э. Дж., Тивари А. К., Чима С., Фриман Н., Браганса Н. и др. Генетическое тестирование на CYP2D6 и CYP2C19 предполагает улучшение результатов лечения антидепрессантами и антипсихотическими препаратами. Psychiatry Res (2018). doi: 10.1016 / j.psychres.2018.02.055
CrossRef Полный текст | Google Scholar
31. Розенблат Дж. Д., Ли Й., Макинтайр Р. С.. Влияние фармакогеномного тестирования на ответ и частоту ремиссии при остром лечении большого депрессивного расстройства: метаанализ. J Affect Disord (2018) 241: 484–91. doi: 10.1016 / j.jad.2018.08.056
PubMed Аннотация | CrossRef Полный текст | Google Scholar
32. Bousman CA, Arandjelovic K, Mancuso SG, Eyre HA, Dunlop BW.Фармакогенетические тесты и ремиссия депрессивных симптомов: метаанализ рандомизированных контролируемых исследований. Фармакогеномика (2019) 20: 37–47. doi: 10.2217 / pgs-2018-0142
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. Таблица фармакогеномных биомаркеров в маркировке лекарств. (2018). 3 августа.
Google Scholar
34. Дейли А.К., Брокмоллер Дж., Броли Ф., Эйхельбаум М., Эванс В.Е., Гонсалес Ф.Дж. и др. Номенклатура аллелей CYP2D6 человека. Фармакогенетика (1996) 6: 193–201. DOI: 10.1097 / 00008571-199606000-00001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35. Робарж Д.Д., Ли Л., Деста З., Нгуен А., Флокхарт Д.А. Номенклатура звездных аллелей: переоснащение для трансляционной геномики. Clin Pharmacol Ther (2007) 82: 244–8. doi: 10.1038 / sj.clpt.6100284
PubMed Аннотация | CrossRef Полный текст | Google Scholar
36. Kalman LV, Agundez J, Appell ML, Black JL, Bell GC, Boukouvala S, et al.Номенклатура фармакогенетических аллелей: рекомендации международной рабочей группы по составлению отчетов о результатах тестов. Clin Pharmacol Ther (2016) 99: 172–85. doi: 10.1002 / cpt.280
PubMed Аннотация | CrossRef Полный текст | Google Scholar
37. Kim J, Lee SY, Lee KA. Вариации числа копий и перестройки генов при генотипировании CYP2D6 с использованием мультиплексной амплификации зонда, зависимой от лигирования, у корейцев. Фармакогеномика (2012) 13: 963–73. doi: 10.2217 / pgs.12.58
PubMed Аннотация | CrossRef Полный текст | Google Scholar
38.Дален П., Даль М.Л., Бернал Руис М.Л., Нордин Дж., Бертилссон Л. 10-гидроксилирование нортриптилина у белых людей с 0, 1, 2, 3 и 13 функциональными генами CYP2D6. Clin Pharmacol Ther (1998) 63: 444–52. doi: 10.1016 / S0009-9236 (98)
-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
39. Bank PCD, Caudle KE, Swen JJ, Gammal RS, Whirl-Carrillo M, Klein TE и др. Сравнение руководств Консорциума по внедрению клинической фармакогенетики и Голландской рабочей группы по фармакогенетике. Clin Pharmacol Ther (2018) 103: 599–618. doi: 10.1002 / cpt.762
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40. Реллинг М.В., Кляйн Т.Э. CPIC: Консорциум по внедрению клинической фармакогенетики Сети исследований фармакогеномики. Clin Pharmacol Ther (2011) 89: 464–7. DOI: 10.1038 / clpt.2010.279
PubMed Аннотация | CrossRef Полный текст | Google Scholar
41. Дален П., Даль М.Л., Эйхельбаум М., Бертилссон Л., Уилкинсон Г.Р.Распределение дебризохина у европеоидов с разными генотипами CYP2D6, в том числе с множественными генами. Фармакогенетика (1999) 9: 697–706. doi: 10.1097 / 01213011-199
PubMed Аннотация | CrossRef Полный текст | Google Scholar
42. Дален П., Даль М., Андерссон К., Бертилссон Л. Ингибирование гидроксилирования дебризохина хинидином у субъектов с тремя или более функциональными генами CYP2D6. Br J Clin Pharmacol (2000) 49: 180–4. DOI: 10,1046 / j.1365-2125.2000.00120.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
43. Кирххайнер Дж., Бертилссон Л., Брюус Х., Вольф А., Рутс И., Бауэр М. Индивидуализированная медицина – внедрение фармакогенетической диагностики в антидепрессивную лекарственную терапию основных депрессивных расстройств. Фармакопсихиатрия (2003) 36 Дополнение 3: S235–43. doi: 10.1055 / s-2003-45136
CrossRef Полный текст | Google Scholar
44. Bolaji OO, Coutts RT, Baker GB. Метаболизм тримипрамина in vitro изоферментом CYP2D6 человека. Res Commun Chem Pathol Pharmacol (1993) 82: 111–20.
PubMed Аннотация | Google Scholar
45. Шенк П.В., ван Фессем М.А., Верплоег-Ван Ридж С., Матот Р.А., ван Гелдер Т., Вулто АГ и др. Связь градуированных аллель-специфических изменений функции CYP2D6 с требуемой дозой имипрамина у большой группы пациентов с депрессией. Mol Psychiatry (2008) 13: 597–605. doi: 10.1038 / sj.mp.4002057
PubMed Аннотация | CrossRef Полный текст | Google Scholar
46.Каваниши С., Лундгрен С., Агрен Х., Бертилссон Л. Повышенная частота дупликации гена CYP2D6 у пациентов с постоянными расстройствами настроения: сверхбыстрый метаболизм антидепрессантов как причина отсутствия ответа. Eur J Clin Pharmacol (2004) 59: 803–7. doi: 10.1007 / s00228-003-0701-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
47. Брокмоллер Дж., Мейнеке И., Кирххайнер Дж. Фармакокинетика миртазапина: энантиоселективные эффекты генотипа сверхбыстрого метаболизатора CYP2D6 и корреляция с побочными эффектами. Clin Pharmacol Ther (2007) 81: 699–707. doi: 10.1038 / sj.clpt.6100116
PubMed Аннотация | CrossRef Полный текст | Google Scholar
49. Dodd S, Boulton DW, Burrows GD, De Vane CL, Norman TR. Метаболизм энантиомеров миртазапина in vitro ферментами цитохрома Р450 человека. Hum Psychopharmacol (2001) 16: 541–44. doi: 10.1002 / hup.328
PubMed Аннотация | CrossRef Полный текст | Google Scholar
50. Закриссон А.Л., Линдблом Б., Алнер Дж. Высокая частота дупликации / размножения гена CYP2D6, указывающая на сверхбыстрый метаболизм среди случаев суицида. Clin Pharmacol Ther (2010) 88: 354–9. DOI: 10.1038 / clpt.2009.216
PubMed Аннотация | CrossRef Полный текст | Google Scholar
51. Penas-Lledo EM, Dorado P, Aguera Z, Gratacos M, Estivill X, Fernandez-Aranda F, et al. Высокий риск суицидальных попыток в течение всей жизни среди сверхбыстрых метаболизаторов CYP2D6 с расстройствами пищевого поведения. Mol Psychiatry (2011) 16: 691–2. doi: 10.1038 / mp.2011.5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
52.Penas-Lledo EM, Blasco-Fontecilla H, Dorado P, Vaquero-Lorenzo C, Baca-Garcia E, Llerena A. CYP2D6 и серьезность суицидальных попыток. Фармакогеномика (2012) 13: 179–84. doi: 10.2217 / pgs.11.146
PubMed Аннотация | CrossRef Полный текст | Google Scholar
54. Penas-Lledo EM, Trejo HD, Dorado P, Ortega A, Jung H, Alonso E, et al. Ультрабыстрый метаболизм CYP2D6 и ранний отказ от монотерапии флуоксетином или амитриптилином у пациентов с тяжелой депрессией. Mol Psychiatry (2013) 18: 8–9.doi: 10.1038 / mp.2012.91
PubMed Аннотация | CrossRef Полный текст | Google Scholar
55. Греден Дж. Ф., Парих С. В., Ротшильд А. Дж., Тэз М. Э., Данлоп Б. В., Дебаттиста С. и др. Влияние фармакогеномики на клинические исходы при большом депрессивном расстройстве в управляемом исследовании: крупном, слепом рандомизированном контролируемом исследовании с участием пациентов и экспертов. J Psychiatr Res (2019) 111: 59–67. doi: 10.1016 / j.jpsychires.2019.01.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
56.Ахмед А.Т., Бирнака Дж. М., Дженкинс Дж., Раш А. Дж., Шинозаки Дж., Велдик М. и др. Фармакокинетико-фармакодинамическое взаимодействие, связанное с ремиссией венлафаксина-XR у пациентов с большим депрессивным расстройством с историей неэффективности лечения циталопрамом / эсциталопрамом. J Affect Disord (2018) 246: 62–68. doi: 10.1016 / j.jad.2018.12.021
PubMed Аннотация | CrossRef Полный текст | Google Scholar
58. Bousman CA, Jaksa P, Pantelis C. Систематическая оценка коммерческого фармакогенетического тестирования в психиатрии: акцент на охват аллелей CYP2D6 и CYP2C19 и отчетность о результатах. Pharmacogenet Genomics (2017) 27: 387–93. doi: 10.1097 / FPC.0000000000000303
PubMed Аннотация | CrossRef Полный текст | Google Scholar
59. Langaee T., Hamadeh I., Chapman AB, Gums JG, Johnson JA. Новый простой метод определения числа копий гена CYP2D6 и идентификации аллеля (ов) с дупликацией / размножением. PLoS One (2015) 10: e0113808. doi: 10.1371 / journal.pone.0113808
PubMed Аннотация | CrossRef Полный текст | Google Scholar
61.Чоу У.Х., Ян FX, де Леон Дж., Барнхилл Дж., Роджерс Т., Кронин М. и др. Расширение пилотного исследования: влияние полиморфизма цитохрома P450 2D6 на исход и затраты, связанные с тяжелым психическим заболеванием. J Clin Psychopharmacol (2000) 20: 246–51. DOI: 10.1097 / 00004714-200004000-00019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
62. Gaedigk A, Sangkuhl K, Whirl-Carrillo M, Klein T, Leeder JS. Прогнозирование фенотипа CYP2D6 из генотипа в разных популяциях мира. Genet Med (2017) 19: 69–76. doi: 10.1038 / gim.2016.80
PubMed Аннотация | CrossRef Полный текст | Google Scholar
63. Chang DD, Eyreeuro HA, Abbott R, Coudreaut M, Baune BT, Shaman JA, et al. Фармакогенетические рекомендации и инструменты поддержки принятия решений для лечения депрессии: применение к пожилым людям. Фармакогеномика (2018) 19: 1269–84. doi: 10.2217 / pgs-2018-0099
PubMed Аннотация | CrossRef Полный текст | Google Scholar
64. Цяо В., Ян И., Себра Р., Мендиратта Г., Гаедигк А., Десник Р. Дж. И др.Полное секвенирование генов цитохрома P450–2D6 в режиме реального времени с использованием длинных считываемых одиночных молекул. Hum Mutat (2016) 37: 315–23. doi: 10.1002 / humu.22936
PubMed Аннотация | CrossRef Полный текст | Google Scholar
65. Пенас-Лледо Е.М., Ллерена А. Вариации, поведение и психопатология CYP2D6: значение для клинических испытаний под руководством фармакогеномики. Br J Clin Pharmacol (2014) 77: 673–83. doi: 10.1111 / bcp.12227
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Связь между вариацией числа копий комплемента 4 и системной красной волчанкой: метаанализ
Javinani A, Ashraf-Ganjouei A, Aslani S, et al. Изучение этиопатогенеза системной красной волчанки: генетическая перспектива. Иммуногенетика. 2019; 71: 283–97.
Артикул Google Scholar
Dammacco R. Системная красная волчанка и поражение глаз: обзор. Clin Exp Med. 2018; 18 (2): 135–49.
CAS Статья Google Scholar
Чен Л., Моррис Д.Л., Вайс Т.Дж. Генетические достижения в области системной красной волчанки: обновление. Curr Opin Rheumatol. 2017; 29 (5): 423–33.
Артикул Google Scholar
Теруэль М., Аларкон-Рикельме, Мэн. Генетическая основа системной красной волчанки: каковы факторы риска и что мы узнали. J Autoimmun. 2016; 74: 161–75.
CAS Статья Google Scholar
Machado-Contreras JR, Muñoz-Valle JF, Cruz A, et al. Распределение полиморфизмов PTPN22 при СКВ из западной Мексики: корреляция с экспрессией мРНК и активностью заболевания. Clin Exp Med. 2016; 16 (3): 399–406.
CAS Статья Google Scholar
Palafox-Sánchez CA, Oregon-Romero E, Salazar-Camarena DC, et al. Связь гаплотипов промотора интерлейкина-10 с восприимчивостью к заболеванию и уровнями IL-10 у мексиканских пациентов с системной красной волчанкой.Clin Exp Med. 2015; 15 (4): 439–46.
Артикул Google Scholar
Бомба Л., Вальтер К., Соранцо Н. Влияние редких и низкочастотных генетических вариантов на общее заболевание. Genome Biol. 2017; 18 (1): 77.
Артикул Google Scholar
Маседо А.С., Исаак Л. Системная красная волчанка и недостаточность ранних компонентов классического пути комплемента.Фронт Иммунол. 2016; 7: 55.
Артикул Google Scholar
Чен М., Даха М.Р., Калленберг К.Г. Система комплемента при системном аутоиммунном заболевании. J Autoimmun. 2010. 34 (3): J276–286.
CAS Статья Google Scholar
Сон М., Даймонд Б., Сантьяго-Шварц Ф. Фундаментальная роль C1q в аутоиммунитете и воспалении. Immunol Res. 2015; 63 (1–3): 101–6.
CAS Статья Google Scholar
Sandhu V, Quan M. СКВ и сывороточный комплекс: причинная, сопутствующая или случайная причина? The Open Rheumatol J. 2017; 11: 113–22.
CAS Статья Google Scholar
Мортенсен С., Кидмос Р.Т., Петерсен С.В. и др. Структурная основа функции компонента С4 комплемента в классическом и лектиновом путях комплемента.J Immunol. 2015; 194 (11): 5488–96.
CAS Статья Google Scholar
Юптнер М., Флаксбарт Ф., Калибе А. и др. Низкое количество копий комплемента C4 и гомозиготный дефицит C4A могут предрасполагать к тяжелому заболеванию и более раннему началу заболевания у пациентов с системной красной волчанкой. Волчанка. 2018; 27 (4): 600–9.
CAS Статья Google Scholar
Tsang A, Sjoe MWP, Bultink IEM, Korswagen LA и др. Комплексный подход к изучению комплемента C4 при системной красной волчанке: полиморфизмы генов, уровни белка и функциональная активность. Мол Иммунол. 2017; 92: 125–31.
Артикул Google Scholar
Chen JY, Wu YL, Mok MY, et al. Влияние вариаций числа копий гена комплемента C4, размерной дихотомии и дефицита C4A на генетический риск и клинические проявления системной красной волчанки в популяциях Восточной Азии.Arthritis Rheumatol (Хобокен, Нью-Джерси). 2016; 68 (6): 1442–533.
CAS Статья Google Scholar
Kim JH, Jung SH, Bae JS, et al. Варианты делеции RABGAP1L, 10q21.3 и C4 связаны с риском системной красной волчанки у корейских женщин. Ревматоидный артрит. 2013; 65 (4): 1055–63.
CAS Статья Google Scholar
Ботева Л., Моррис Д.Л., Кортес-Эрнандес Дж. И др.Генетически детерминированные состояния частичной недостаточности комплемента C4 не являются независимыми факторами риска СКВ у населения Великобритании и Испании. Am J Hum Genet. 2012. 90 (3): 445–56.
CAS Статья Google Scholar
Lv Y, He S, Zhang Z, et al. Подтверждение вариабельности числа копий гена C4 и связи с системной красной волчанкой в китайской ханьской популяции. Rheumatol Int. 2012. 32 (10): 3047–53.
CAS Статья Google Scholar
Каматани Ю., Мацуда К., Охиси Т. и др. Выявление значительной ассоциации однонуклеотидного полиморфизма TNXB с системной красной волчанкой в популяции Японии. J Hum Genet. 2008. 53 (1): 64–73.
CAS Статья Google Scholar
Yang Y, Chung EK, Wu YL, et al. Вариации числа копий гена и связанные с ними полиморфизмы компонента комплемента С4 при системной красной волчанке (СКВ) человека: низкое число копий является фактором риска, а высокое число копий является фактором защиты от восприимчивости к СКВ у американцев европейского происхождения.Am J Hum Genet. 2007. 80 (6): 1037–54.
CAS Статья Google Scholar
Lo CK, Mertz D, Loeb M. Newcastle-Ottawa Scale: сравнение оценок рецензентов и авторов. BMC Med Res Methodol. 2014; 1 (14): 45.
Артикул Google Scholar
Yu CY, Belt KT, Giles CM, et al. Структурные основы полиморфизма компонентов комплемента человека C4A и C4B: размер гена, реактивность и антигенность.EMBO J. 1986; 5 (11): 2873–81.
CAS Статья Google Scholar
Dodds AW, Ren XD, Willis AC, et al. Механизм реакции внутреннего тиоэфира в человеческом компоненте комплемента С4. Природа. 1996. 379 (6561): 177–9.
CAS Статья Google Scholar
Кэрролл Х.А., Тумпакари З., Джонсон Л. и др. Предполагаемая осуществимость методов снижения предвзятости публикации.PLoS ONE. 2017; 12 (10): e0186472.
Артикул Google Scholar
Варианты числа копий (CNV): мощный инструмент для моделирования ASD на основе iPSC | Молекулярный аутизм
Зандт Ф., Прайор М., Кириос М. Повторяющееся поведение у детей с высокофункциональным аутизмом и обсессивно-компульсивным расстройством. J Autism Dev Disord. 2007. 37 (2): 251–9.
PubMed Статья Google Scholar
Фахури М. Расстройства аутистического спектра: обзор клинических особенностей, теории и диагностики. Int J Dev Neurosci. 2015; 43: 70–7.
PubMed Статья Google Scholar
Пиао И, Хунг С.С., Лим С.И., Вонг Р.С., Ко МС. Эффективное создание свободных от интеграции индуцированных человеком плюрипотентных стволовых клеток из кератиноцитов путем простой трансфекции эписомальных векторов. Стволовые клетки Transl Med. 2014; 3 (7): 787–91.
CAS PubMed PubMed Central Статья Google Scholar
Brown ME, Rondon E, Rajesh D, Mack A, Lewis R, Feng X и др. Получение индуцированных плюрипотентных стволовых клеток из Т-лимфоцитов периферической крови человека. PLoS One. 2010; 5 (6): e11373.
PubMed PubMed Central Статья CAS Google Scholar
Секи Т., Юаса С., Фукуда К. Получение индуцированных плюрипотентных стволовых клеток из периферических циркулирующих Т-клеток человека. Curr Protoc Stem Cell Biol. 2011; Глава 4: Unit4A 3.
PubMed Google Scholar
Чжоу Т., Бенда С., Данзингер С., Хуанг Й., Хо Дж. К., Ян Дж. И др. Генерация индуцированных человеком плюрипотентных стволовых клеток из образцов мочи. Nat Protoc. 2012; 7 (12): 2080–209.
CAS PubMed Статья Google Scholar
Чжоу Т., Бенда С., Дузингер С., Хуанг И, Ли Х, Ли И и др. Генерация индуцированных плюрипотентных стволовых клеток из мочи.J Am Soc Nephrol. 2011; 22 (7): 1221–8.
PubMed PubMed Central Статья CAS Google Scholar
Velasco S, Kedaigle AJ, Simmons SK, Nash A, Rocha M, Quadrato G, et al. Отдельные органоиды головного мозга воспроизводимо образуют клеточное разнообразие коры головного мозга человека. Природа. 2019; 570 (7762): 523–7.
CAS PubMed PubMed Central Статья Google Scholar
Юн С.Дж., Элахи Л.С., Паска А.М., Мартон Р.М., Гордон А., Рева О. и др. Надежность генерации корковых органоидов человека. Нат методы. 2019; 16 (1): 75–8.
CAS PubMed Статья Google Scholar
Ефремова В., Маникакис Г., Креффт О., Джабали А., Вейнанс К., Вилкенс Р. и др. Модель коркового развития на основе органоидов выявляет не автономные клетки дефекты в передаче сигналов Wnt, вносящие вклад в синдром Миллера-Дикера.Cell Rep., 2017; 19 (1): 50–9.
CAS PubMed Статья Google Scholar
Меллиос Н., Фельдман Д.А., Шеридан С.Д., ИП JPK, Квок С., Амоа С.К. и др. Регулируемые MeCP2 miRNA контролируют ранний нейрогенез человека посредством различных эффектов на передачу сигналов ERK и AKT. Мол Психиатрия. 2018; 23 (4): 1051–65.
CAS PubMed Статья Google Scholar
Ван П., Мохтари Р., Педроса Е., Киршенбаум М., Байрак С., Чжэн Д. и др.CRISPR / Cas9-опосредованный гетерозиготный нокаут гена аутизма CHD8 и характеристика его транскрипционных сетей в церебральных органоидах, полученных из iPS-клеток. Молочный аутизм. 2017; 8: 11.
PubMed PubMed Central Статья CAS Google Scholar
Lopez-Tobon A, Villa CE, Cheroni C, Trattaro S, Caporale N, Conforti P, et al. Органоиды коры головного мозга человека демонстрируют дифференциальную функцию GSK3 в корковом нейрогенезе.Отчеты о стволовых клетках. 2019; 13 (5): 847–61.
CAS PubMed PubMed Central Статья Google Scholar
Munnich A, Demily C, Frugere L, Duwime C, Malan V, Barcia G, et al. Влияние клинических генетических консультаций на частоту диагностики у детей и молодых людей с расстройствами аутистического спектра. Молочный аутизм. 2019; 10:33.
PubMed PubMed Central Статья CAS Google Scholar
Себат Дж., Лакшми Б., Малхотра Д., Троге Дж., Лезе-Мартин С., Уолш Т. и др. Сильная связь мутаций числа копий de novo с аутизмом. Наука. 2007. 316 (5823): 445–9.
CAS PubMed PubMed Central Статья Google Scholar
Киров Г., Рис Э., Уолтерс Дж. Что психиатру нужно знать о вариантах числа копий. Б. Дж. Психологические достижения. 2015; 21 (3): 157–63.
Google Scholar
Рис Э., Уолтерс Дж. Т., Георгиева Л., Айлз А. Р., Чембер К. Д., Ричардс А. Л. и др. Анализ вариаций числа копий в 15 локусах, ассоциированных с шизофренией. Br J Psychiatry. 2014. 204 (2): 108–14.
PubMed PubMed Central Статья Google Scholar
Кендалл К.М., Рис Э., Эскотт-Прайс В., Эйнон М., Томас Р., Хьюитт Дж. И др. Когнитивные способности среди носителей патогенных вариантов числа копий: анализ 152 000 субъектов биобанка Великобритании.Биол Психиатрия. 2017; 82 (2): 103–10.
PubMed Статья Google Scholar
Пинто Д., Делаби Е., Мерико Д., Барбоса М., Мерикангас А., Клей Л. и др. Конвергенция генов и нарушение регуляции клеточных путей при расстройствах аутистического спектра. Am J Hum Genet. 2014; 94 (5): 677–94.
CAS PubMed PubMed Central Статья Google Scholar
Малхотра Д., Себат Дж.CNVs: предвестники редкой вариантной революции в психиатрической генетике. Клетка. 2012. 148 (6): 1223–41.
CAS PubMed PubMed Central Статья Google Scholar
van der Zwaag B, Staal WG, Hochstenbach R, Poot M, Spierenburg HA, de Jonge MV, et al. Совместная микродупликация хромосомы 15q11.2 выявляет два гена риска для расстройства аутистического спектра. Am J Med Genet B Neuropsychiatr Genet. 2010. 153B (4): 960–6.
PubMed PubMed Central Google Scholar
Депьенн С., Морено-де-Лука Д., Херон Д., Бутеиллер Д., Женнетье А., Делорм Р. и др. Скрининг геномных перестроек и аномалий метилирования в области 15q11-q13 при расстройствах аутистического спектра. Биол Психиатрия. 2009. 66 (4): 349–59.
CAS PubMed Статья Google Scholar
Серур Ю., Софрин Фрумер Д., Даон К., Соболь-Хавиа Д., Вайнбергер Р., Шульман С. и др.Психиатрические расстройства и аутизм у детей раннего возраста с синдромом делеции 22q11.2 по сравнению с детьми с идиопатическим аутизмом. Eur Psychiatry. 2019; 55: 116–21.
PubMed Статья Google Scholar
Морроу Б.Э., Макдональд-МакГинн Д.М., Эмануэль Б.С., Вермиш-младший, Скамблер П.Дж. Молекулярная генетика синдрома делеции 22q11.2. Am J Med Genet A. 2018; 176 (10): 2070–81.
CAS PubMed PubMed Central Статья Google Scholar
Мерфи KC, Джонс Л.А., Оуэн MJ. Высокие показатели шизофрении у взрослых с велокардиофациальным синдромом. Arch Gen Psychiatry. 1999. 56 (10): 940–5.
CAS PubMed Статья Google Scholar
Schneider M, Debbane M, Bassett AS, Chow EW, Fung WL, van den Bree M, et al. Психиатрические расстройства от детства до взрослого возраста при синдроме делеции 22q11.2: результаты международного консорциума по мозгу и поведению при 22q11.2 делеционный синдром. Am J Psychiatry. 2014. 171 (6): 627–39.
PubMed PubMed Central Статья Google Scholar
Демарел В., Мостовой Ю., Йилмаз Ф., Вервурт Л., Пастор С., Хестанд М.С. и др. Низкокопийные повторы 22q11 характеризуются беспрецедентным размером и структурной изменчивостью. Genome Res. 2019; 29 (9): 1389–401.
CAS PubMed PubMed Central Статья Google Scholar
Zhang X, Zhang Y, Zhu X, Purmann C, Haney MS, Ward T, et al. Локальные и глобальные взаимодействия хроматина изменяются большими геномными делециями, связанными с развитием человеческого мозга. Nat Commun. 2018; 9 (1): 5356.
PubMed PubMed Central Статья CAS Google Scholar
Гирираджан С., Розенфельд Дж. А., Коу Б. П., Парих С., Фридман Н., Гольдштейн А. и др. Фенотипическая гетерогенность геномных нарушений и редких вариантов числа копий.N Engl J Med. 2012. 367 (14): 1321–31.
CAS PubMed PubMed Central Статья Google Scholar
Girirajan S, Rosenfeld JA, Cooper GM, Antonacci F, Siswara P, Itsara A, et al. Повторяющаяся микроделеция 16p12.1 поддерживает модель с двумя ударами для серьезной задержки развития. Нат Жене. 2010. 42 (3): 203–9.
CAS PubMed PubMed Central Статья Google Scholar
Bergen SE, Ploner A, Howrigan D, Group CNVA. Рабочая группа по шизофрении Психиатрической геномики C, O’Donovan MC, et al. совместный вклад редких вариантов числа копий и общих SNP в риск шизофрении. Am J Psychiatry. 2019; 176 (1): 29–35.
PubMed Статья Google Scholar
Мартин Дж., О’Донован М.С., Тапар А., Лэнгли К., Уильямс Н. Относительный вклад общих и редких генетических вариантов в СДВГ.Перевод Психиатрия. 2015; 5: e506.
CAS PubMed PubMed Central Статья Google Scholar
Велинов М. Вариации числа копий генома в клинике аутизма в работе. Front Cell Neurosci. 2019; 13:57.
CAS PubMed PubMed Central Статья Google Scholar
Bassett AR. Редактирование генома hiPSC с помощью CRISPR / Cas9: модели болезни.Мамм Геном. 2017; 28 (7-8): 348–64.
CAS PubMed PubMed Central Статья Google Scholar
Canver MC, Bauer DE, Dass A, Yien YY, Chung J, Masuda T. и др. Характеристика эффективности геномной делеции, опосредованной кластерами коротких палиндромных повторов (CRISPR) / нуклеаза Cas9 с регулярными интервалами в клетках млекопитающих. J Biol Chem. 2014. 289 (31): 21312–24.
PubMed PubMed Central Статья CAS Google Scholar
Tai DJ, Ragavendran A, Manavalan P, Stortchevoi A, Seabra CM, Erdin S и др. Инженерия микроделеций и микродупликаций путем нацеливания на сегментарные дупликации с помощью CRISPR. Nat Neurosci. 2016; 19 (3): 517–22.
CAS PubMed PubMed Central Статья Google Scholar
Li X, Burnight ER, Cooney AL, Malani N, Brady T., Sander JD, et al. Инструменты транспозазы piggyBac для геномной инженерии. Proc Natl Acad Sci U S A.2013; 110 (25): E2279–87.
CAS PubMed PubMed Central Статья Google Scholar
Адхья Д., Сваруп В., Надь Р., Шум С., Новосиад П., Йозвик К. М. и др. Атипичный нейрогенез и генерация возбудительно-ингибирующих предшественников в индуцированных плюрипотентных стволовых клетках (ИПСК) от аутичных людей. bioRxiv. 2019: 349415.
Дено Э., Фахим М., Уайт С.Х., Родригес, округ Колумбия, Сан С., Вэй В. и др. CNTN5 (-) (/ +) или EHMT2 (-) (/ +) нейроны, полученные из ИПСК человека от людей с аутизмом, развивают гиперактивные нейронные сети.eLife. 2019; 8.
Лам М., Мусульманин М., Бриойс Дж., Пронк Р.Дж., Улин Э., Элстром И.Д. и др. Одноклеточный анализ пациента с аутизмом с биаллельной делецией NRXN1-альфа показывает искаженный выбор судьбы в нейрональных предшественниках и нарушение нейрональной функции. Exp Cell Res. 2019; 383 (1): 111469.
PubMed Статья CAS Google Scholar
Гридина М.М., Матвеева Н.М., Фишман В.С., Мензоров А.Г., Кизилова Н.А., Береговой Н.А. и др.Аллель-специфическая предвзятая экспрессия гена CNTN6 в нейронах, полученных из iPS-клеток пациента с умственной отсталостью и микродупликацией 3p26.3 с участием гена CNTN6. Mol Neurobiol. 2018; 55 (8): 6533–46.
CAS PubMed Статья Google Scholar
Piovani G, Lanzi G, Ferraro RM, Masneri S, Barisani C, Savio G, et al. Генерация индуцированных плюрипотентных стволовых клеток (ИПСК) от пациента с синдромом Кри дю Шат.Stem Cell Res. 2019; 35: 101393.
CAS PubMed Статья Google Scholar
Адамо А., Аташпаз С., Жермен П.Л., Занелла М., Д’Агостино Г., Альбертин В. и др. 7q11.23 дозозависимая дисрегуляция в плюрипотентных стволовых клетках человека влияет на программы транскрипции в связанных с заболеванием клонах. Нат Жене. 2015; 47 (2): 132–41.
CAS PubMed Статья Google Scholar
Frega M, Linda K, Keller JM, Gumus-Akay G, Mossink B, van Rhijn JR, et al. Дисфункция нейрональной сети в модели синдрома Клифстры, опосредованная усиленной передачей сигналов NMDAR. Nat Commun. 2019; 10 (1): 4928.
PubMed PubMed Central Статья CAS Google Scholar
Willemsen MH, Beunders G, Callaghan M, de Leeuw N, Nillesen WM, Yntema HG, et al. Семейный синдром Клифстры, обусловленный соматическим мозаицизмом матери для интерстициального 9q34.3 микроделеции. Clin Genet. 2011; 80 (1): 31–8.
CAS PubMed Статья Google Scholar
Мариани Дж., Коппола Дж., Чжан П., Абызов А., Провини Л., Томасини Л. и др. FOXG1-зависимая дисрегуляция дифференцировки ГАМК / глутаматных нейронов при расстройствах аутистического спектра. Клетка. 2015; 162 (2): 375–90.
CAS PubMed PubMed Central Статья Google Scholar
Абызов А., Мариани Дж., Палеев Д., Чжан Ю., Хейни М.С., Томасини Л. и др. Мозаицизм соматического числа копий в коже человека, выявленный индуцированными плюрипотентными стволовыми клетками. Природа. 2012. 492 (7429): 438–42.
CAS PubMed PubMed Central Статья Google Scholar
Das DK, Tapias V, D’Aiuto L, Chowdari KV, Francis L, Zhi Y, et al. Генетические и морфологические особенности нейронов, полученных из ИПСК человека с хромосомой 15q11.2 (BP1-BP2) делеции. Мол нейропсихиатрии. 2015; 1 (2): 116–23.
CAS PubMed PubMed Central Статья Google Scholar
Ариока Ю., Кусима И., Мори Д., Одзаки Н. Три линии индуцированных плюрипотентных стволовых клеток, полученные от пациента с синдромом дупликации 15q11.2-q13.1. Stem Cell Res. 2018; 31: 240–3.
CAS PubMed Статья Google Scholar
Germain ND, Chen PF, Plocik AM, Glatt-Deeley H, Brown J, Fink JJ и др. Анализ экспрессии генов индуцированных человеком нейронов, полученных из плюрипотентных стволовых клеток, несущих варианты хромосомы 15q11-q13.1 с числом копий. Молочный аутизм. 2014; 5: 44.
PubMed PubMed Central Статья CAS Google Scholar
Gillentine MA, Yin J, Bajic A, Zhang P, Cummock S, Kim JJ, et al. Функциональные последствия изменений числа копий CHRNA7 в индуцированных плюрипотентных стволовых клетках и клетках-предшественниках нейронов.Am J Hum Genet. 2017; 101 (6): 874–87.
CAS PubMed PubMed Central Статья Google Scholar
Deshpande A, Yadav S, Dao DQ, Wu ZY, Hokanson KC, Cahill MK, et al. Клеточные фенотипы нейронов, происходящих от ИПСК человека, из генетической модели расстройства аутистического спектра. Cell Rep., 2017; 21 (10): 2678–87.
CAS PubMed PubMed Central Статья Google Scholar
Джонстон М., Васиштха Н.А., Барбу М.К., Дандо О., Берр К., Кристофер Э. и др. Устранение дефицита пролиферации, вызванного микродупликацией хромосомы 16p13.11, посредством нацеливания на передачу сигналов NFkappaB: интегрированное исследование полученных от пациента клеток-предшественников нейронов, церебральных органоидов и визуализации мозга in vivo. Мол Психиатрия. 2019; 24 (2): 294–311.
CAS PubMed Статья Google Scholar
Берштейн М., Новаковски Т.Дж., Пыльца А.А., Ди Лулло Э., Нене А., Виншоу-Борис А. и др.Церебральные органоиды, полученные из ИПСК человека, моделируют клеточные особенности лиссэнцефалии и выявляют длительный митоз наружной радиальной глии. Стволовая клетка. 2017; 20 (4): 435–49 e4.
CAS PubMed PubMed Central Статья Google Scholar
Щегловитов А., Щегловитова О., Ядзава М., Портманн Т., Шу Р., Себастьяно В. и др. SHANK3 и IGF1 восстанавливают синаптический дефицит в нейронах пациентов с синдромом делеции 22q13. Природа.2013. 503 (7475): 267–71.
CAS PubMed PubMed Central Статья Google Scholar
Катурия А., Новосиад П., Джагасиа Р., Эйгнер С., Тейлор Р.Д., Андреа Л.С. и др. Нейроны, полученные из стволовых клеток аутичных людей с мутацией SHANK3, обнаруживают морфогенетические аномалии на раннем этапе развития. Мол Психиатрия. 2018; 23 (3): 735–46.
CAS PubMed Статья Google Scholar
Cocks G, Curran S, Gami P, Uwanogho D, Jeffries AR, Kathuria A и др. Польза специфических для пациента индуцированных плюрипотентных стволовых клеток для моделирования расстройств аутистического спектра. Психофармакология. 2014. 231 (6): 1079–88.
CAS PubMed Статья Google Scholar
Росс П.Дж., Чжан В.Б., Мок RSF, Заславский К., Дено Э., Д’Абате Л. и др. Синаптическая дисфункция в нейронах человека с ассоциированными с аутизмом делециями в PTCHD1-AS.Биол Психиатрия. 2019.
Machado CO, Griesi-Oliveira K, Rosenberg C, Kok F, Martins S, Passos-Bueno MR, et al. Коллибистин связывает и подавляет передачу сигналов mTORC1: потенциально новый механизм, способствующий умственной отсталости и аутизму. Eur J Hum Genet. 2016; 24 (1): 59–65.
PubMed Статья CAS Google Scholar
Nageshappa S, Carromeu C, Trujillo CA, Mesci P, Espuny-Camacho I, Pasciuto E, et al.Измененная нейронная сеть и спасение в модели дупликации MECP2 человека. Мол Психиатрия. 2016; 21 (2): 178–88.
CAS PubMed Статья Google Scholar
Бакгольц Дж. В., Мейер-Линденберг А. Психопатология и человеческий коннектом: к трансдиагностической модели риска психических заболеваний. Нейрон. 2012; 74 (6): 990–1004.
CAS PubMed Статья Google Scholar
Фальк А., Хайне В.М., Харвуд А.Дж., Салливан П.Ф., Пейтц М., Брюстл О. и др. Моделирование психических расстройств: от геномных данных до клеточных фенотипов. Мол Психиатрия. 2016; 21 (9): 1167–79.
CAS PubMed PubMed Central Статья Google Scholar
Gouder L, Vitrac A, Goubran-Botros H, Danckaert A, Tinevez JY, Andre-Leroux G, et al. Измененный спиногенез в нейронах коры головного мозга, происходящих от ИПСК, у пациентов с аутизмом, несущих de novo мутации SHANK3.Научный доклад 2019; 9 (1): 94.
PubMed PubMed Central Статья CAS Google Scholar
ДеРоса Б.А., Эль-Хокайем Дж., Артимович Е., Гарсия-Сержи С., Филлипс А.В., Ван Бувен Д. и др. Конвергентные пути при идиопатическом аутизме выявлены с помощью транскриптомного анализа динамики нейронов, полученных от пациента. Научный доклад 2018; 8 (1): 8423.
PubMed PubMed Central Статья CAS Google Scholar
Schafer ST, Paquola ACM, Stern S, Gosselin D, Ku M, Pena M и др. Патологический прайминг вызывает гетерохронность генной сети развития в нейронах, происходящих от аутистов. Nat Neurosci. 2019; 22 (2): 243–55.
CAS PubMed PubMed Central Статья Google Scholar
Mozzetta C, Pontis J, Fritsch L, Robin P, Portoso M, Proux C. et al. Гистон h4 лизин 9 метилтрансферазы G9a и GLP регулируют опосредованное поликомб репрессивным комплексом 2 подавление гена.Mol Cell. 2014. 53 (2): 277–89.
CAS PubMed Статья Google Scholar
Khattak S, Brimble E, Zhang W, Zaslavsky K, Strong E, Ross PJ, et al. Человеческие индуцированные нейроны, полученные из плюрипотентных стволовых клеток, как модель синдрома Вильямса-Бёрена. Мол мозг. 2015; 8 (1): 77.
PubMed PubMed Central Статья CAS Google Scholar
Хуанг Дж., Чен С., Чен Х, Чжэн Дж., Сюй З., Достпараст Торшизи А. и др.Выявление функциональной связи между делециями SHANK3 и недостаточностью развития нервной системы с помощью нейронов человека, происходящих из ИПСК. Фронт нейроанат. 2019; 13:23.
CAS PubMed PubMed Central Статья Google Scholar
Siniscalco D, Schultz S, Brigida AL, Antonucci N. Воспаление и нейроиммунные нарушения при расстройствах аутистического спектра. Фармацевтика. 2018; 11 (2).
Жан Й., Паоличелли Р.С., Сфораццини Ф., Вайнхард Л., Боласко Дж., Пагани Ф. и др.Недостаточная передача сигналов между нейронами и микроглией приводит к нарушению функциональной связи мозга и нарушению социального поведения. Nat Neurosci. 2014. 17 (3): 400–6.
CAS PubMed Статья Google Scholar
Хаан Н., Картер Дж., Вестакотт Л.Дж., Оуэн М.Дж., Грей В.П., Холл Дж. И др. Гаплонедостаточность гена риска шизофрении Cyfip1 вызывает аномальный постнатальный нейрогенез гиппокампа через новый микроглиозависимый механизм. bioRxiv. 2018: 417832.
Руссо Ф.Б., Фрейтас BC, Пигнатари ГК, Фернандес И.Р., Себат Дж., Муотри А.Р. и др. Моделирование взаимодействия нейронов и астроцитов при аутизме с использованием индуцированных человеком плюрипотентных стволовых клеток. Биол Психиатрия. 2018; 83 (7): 569–78.
PubMed Статья Google Scholar
Eftekharian MM, Ghafouri-Fard S, Noroozi R, Omrani MD, Arsang-Jang S, Ganji M, et al. Цитокиновый профиль у аутичных пациентов. Цитокин.2018; 108: 120–6.
CAS PubMed Статья Google Scholar
Nadadhur AG, Alsaqati M, Gasparotto L, Cornelissen-Steijger P, van Hugte E, Dooves S, et al. Взаимодействия нейронов и глии увеличивают нейрональные фенотипы в моделях пациентов с туберозным склерозом, полученных из ИПСК. Отчеты о стволовых клетках. 2019; 12 (1): 42–56.
PubMed Статья Google Scholar
Courchesne E, Carper R, Akshoomoff N. Свидетельства чрезмерного роста мозга в первый год жизни при аутизме. Джама. 2003. 290 (3): 337–44.
PubMed Статья Google Scholar
Courchesne E, Karns CM, Davis HR, Ziccardi R, Carper RA, Tigue ZD, et al. Необычные модели роста мозга в раннем возрасте у пациентов с аутизмом: исследование МРТ. Неврология. 2001. 57 (2): 245–54.
CAS PubMed Статья Google Scholar
Lainhart JE, Piven J, Wzorek M, Landa R, Santangelo SL, Coon H и др. Макроцефалия у детей и взрослых с аутизмом. J Am Acad Детская подростковая психиатрия. 1997. 36 (2): 282–90.
CAS PubMed Статья Google Scholar
Piven J, Arndt S, Bailey J, Havercamp S, Andreasen NC, Palmer P. Исследование размера мозга при аутизме с помощью МРТ. Am J Psychiatry. 1995. 152 (8): 1145–9.
CAS PubMed Статья Google Scholar
Лин А., Чинг ЦРК, Вайди А., Сан Д., Йонас Р.К., Ялбжиковски М. и др. Картирование влияния дозировки гена 22q11.2 на морфометрию мозга. J Neurosci. 2017; 37 (26): 6183–99.
CAS PubMed PubMed Central Статья Google Scholar
Sun D, CRK C, Lin A, Forsyth JK, Kushan L, Vajdi A, et al. Крупномасштабное картирование корковых изменений при синдроме делеции 22q11.2: конвергенция с идиопатическим психозом и влияние размера делеции.Мол Психиатрия. 2018.
Fan CC, Brown TT, Bartsch H, Kuperman JM, Hagler DJ Jr, Schork A, et al. Специфический для синдрома Вильямса нейроанатомический профиль и его связи с особенностями поведения. NeuroImage Clin. 2017; 15: 343–7.
PubMed PubMed Central Статья Google Scholar
Меда С.А., Приуэллер Дж. Р., Торнтон-Уэллс ТА. Региональные различия мозга в толщине коры, площади поверхности и подкорковом объеме у людей с синдромом Вильямса.PLoS One. 2012; 7 (2): e31913.
CAS PubMed PubMed Central Статья Google Scholar
Сильва А.И., Ульфарссон МО, Стефанссон Х., Густафссон О., Уолтерс Г.Б., Линден ДЭЙ и др. Взаимные изменения белого вещества, связанные с изменением числа копий в 15q11.2 BP1-BP2: исследование с визуализацией тензора диффузии. Биол Психиатрия. 2019; 85 (7): 563–72.
CAS PubMed PubMed Central Статья Google Scholar
Ulfarsson MO, Walters GB, Gustafsson O, Steinberg S, Silva A, Doyle OM и др. 15q11.2 CNV влияет на когнитивные, структурные и функциональные корреляты дислексии и дискалькулии. Перевод Психиатрия. 2017; 7 (4): e1109.
CAS PubMed PubMed Central Статья Google Scholar
Комитет по написанию E-CNVWG, van der Meer D, Sonderby IE, Kaufmann T, Walters GB, Abdellaoui A, et al. Ассоциация вариации числа копий файла 15q11.2 Область BP1-BP2 с корковой и подкорковой морфологией и познанием. JAMA Psychiatry. 2019; 77: 1–11.
Google Scholar
Maillard AM, Ruef A, Pizzagalli F, Migliavacca E, Hippolyte L, Adaszewski S, et al. Локус 16p11.2 модулирует структуры мозга, общие для аутизма, шизофрении и ожирения. Мол Психиатрия. 2015; 20 (1): 140–7.
CAS PubMed Статья Google Scholar
Martin-Brevet S, Rodriguez-Herreros B, Nielsen JA, Moreau C, Modenato C, Maillard AM и др. Количественная оценка влияния вариантов числа копий 16p11.2 на структуру мозга: многоузловое генетическое исследование. Биол Психиатрия. 2018; 84 (4): 253–64.
CAS PubMed Статья Google Scholar
Куреши А.Ю., Мюллер С., Снайдер А.З., Мукерджи П., Берман Дж. И., Робертс Т.П. и др. Противодействие различиям в мозге носителей делеции и дупликации 16p11.2.J Neurosci. 2014. 34 (34): 11199–211.
CAS PubMed PubMed Central Статья Google Scholar
Sonderby IE, Gustafsson O, Doan NT, Hibar DP, Martin-Brevet S, Abdellaoui A, et al. Дозовая реакция варианта дистального числа копий 16p11.2 на интракраниальный объем и базальные ганглии. Мол Психиатрия. 2020; 25 (3): 584–602.
PubMed Статья CAS Google Scholar
Chen CP, Keown CL, Jahedi A, Nair A, Pflieger ME, Bailey BA, et al. Диагностическая классификация внутренней функциональной связности выделяет соматосенсорные области, режим по умолчанию и визуальные области при аутизме. Neuroimage Clin. 2015; 8: 238–45.
PubMed PubMed Central Статья Google Scholar
Хаар С., Берман С., Берманн М., Динштейн И. Анатомические аномалии при аутизме? Cereb Cortex. 2016; 26 (4): 1440–52.
PubMed Статья Google Scholar
Маркетто М.С., Белинсон Х., Тиан Й., Фрейтас BC, Фу С., Вадодария К. и др. Измененная пролиферация и сети в нервных клетках, происходящих от идиопатических аутичных людей. Мол Психиатрия. 2017; 22 (6): 820–35.
CAS PubMed Статья PubMed Central Google Scholar
Coghlan S, Horder J, Inkster B, Mendez MA, Murphy DG, Nutt DJ. Дисфункция системы ГАМК при аутизме и связанных с ним расстройствах: от синапсов до симптомов.Neurosci Biobehav Rev.2012; 36 (9): 2044–55.
CAS PubMed PubMed Central Статья Google Scholar
Ecker C, Suckling J, Deoni SC, Lombardo MV, Bullmore ET, Baron-Cohen S, et al. Анатомия мозга и ее связь с поведением у взрослых с расстройством аутистического спектра: многоцентровое исследование магнитно-резонансной томографии. Arch Gen Psychiatry. 2012. 69 (2): 195–209.
PubMed Статья Google Scholar
Lenroot RK, Yeung PK. Неоднородность расстройств аутистического спектра: что мы узнали из исследований нейровизуализации? Front Hum Neurosci. 2013; 7: 733.
PubMed PubMed Central Статья Google Scholar
Зикопулос Б., Барбас Х. Изменения в префронтальных аксонах могут нарушить работу сети при аутизме. J Neurosci. 2010. 30 (44): 14595–609.
CAS PubMed PubMed Central Статья Google Scholar
Nuninga JO, Bohlken MM, Koops S, Fiksinski AM, Mandl RCW, Breetvelt EJ, et al. Аномалии белого вещества у пациентов с синдромом делеции 22q11.2, демонстрирующие снижение когнитивных функций. Psychol Med. 2018; 48 (10): 1655–63.
PubMed Статья Google Scholar
Drakesmith M, Parker GD, Smith J, Linden SC, Rees E, Williams N, et al. Генетический риск шизофрении и задержки развития связан с формой и микроструктурой срединных структур белого вещества.Перевод Психиатрия. 2019; 9 (1): 102.
PubMed PubMed Central Статья Google Scholar
Сильва А.И., Хаддон Дж. Э., Ахмед Сайед Ю., Трент С., Лин Т. Е., Патель Ю. и др. У крыс с гаплонедостаточностью Cyfip1 наблюдаются изменения белого вещества, истончение миелина, аномальные олигодендроциты и поведенческая негибкость. Nat Commun. 2019; 10 (1): 3455.
PubMed PubMed Central Статья CAS Google Scholar
Хусейн С.М., Батада Н.Н., Вуористо С., Чинг Р.В., Аутио Р., Нарва Е. и др. Изменение числа копий и выбор во время перепрограммирования до плюрипотентности. Природа. 2011. 471 (7336): 58–62.
CAS PubMed Статья Google Scholar
Draper JS, Smith K, Gokhale P, Moore HD, Maltby E, Johnson J, et al. Повторяющееся увеличение хромосом 17q и 12 в культивируемых эмбриональных стволовых клетках человека. Nat Biotechnol. 2004. 22 (1): 53–4.
CAS PubMed Статья Google Scholar
International Stem Cell I, Amps K, Andrews PW, Anyfantis G, Armstrong L, Avery S, et al. Скрининг этнически разнообразных эмбриональных стволовых клеток человека определяет минимальный ампликон 20 хромосомы, обеспечивающий преимущество в росте. Nat Biotechnol. 2011; 29 (12): 1132–44.
Артикул CAS Google Scholar
Laurent LC, Ulitsky I., Slavin I, Tran H, Schork A, Morey R, et al. Динамические изменения числа копий генов плюрипотентности и пролиферации клеток в человеческих ЭСК и ИПСК во время репрограммирования и времени в культуре.Стволовая клетка. 2011; 8 (1): 106–18.
CAS PubMed PubMed Central Статья Google Scholar
Maitra A, Arking DE, Shivapurkar N, Ikeda M, Stastny V, Kassauei K, et al. Геномные изменения в культивируемых эмбриональных стволовых клетках человека. Нат Жене. 2005. 37 (10): 1099–103.
CAS PubMed Статья Google Scholar
Mertens J, Paquola ACM, Ku M, Hatch E, Bohnke L, Ladjevardi S, et al.Непосредственно перепрограммированные нейроны человека сохраняют связанные со старением транскриптомные сигнатуры и выявляют связанные с возрастом ядерно-цитоплазматические дефекты. Стволовая клетка. 2015; 17 (6): 705–18.
CAS PubMed PubMed Central Статья Google Scholar
Sheng C, Jungverdorben J, Wiethoff H, Lin Q, Flitsch LJ, Eckert D, et al. Стабильно самообновляющиеся нервные стволовые клетки, полученные из крови взрослых, демонстрирующие способность к паттерну и эпигенетическое омоложение.Nat Commun. 2018; 9 (1): 4047.
PubMed PubMed Central Статья CAS Google Scholar
Приджионе А., Хоссини А.М., Лихтнер Б., Серин А., Фаулер Б., Меггес М. и др. Связанные с митохондриями механизмы гибели клеток сбрасываются до эмбрионального состояния в старых донорских iPS-клетках, несущих хромосомные аберрации. PLoS One. 2011; 6 (11): e27352.
CAS PubMed PubMed Central Статья Google Scholar
Perrera V, Martello G. Как перепрограммирование плюрипотентности влияет на геномный импринтинг? Front Cell Dev Biol. 2019; 7: 76.
PubMed PubMed Central Статья Google Scholar
Xie Y, Schutte RJ, Ng NN, Ess KC, Schwartz PH, O’Dowd DK. Воспроизводимое и эффективное создание функционально активных нейронов из hiPSC человека для доклинического моделирования заболеваний. Stem Cell Res. 2018; 26: 84–94.
CAS PubMed Статья Google Scholar
van den Hurk M, Erwin JA, Yeo GW, Gage FH, Bardy C. Протокол Patch-Seq для анализа электрофизиологии, морфологии и транскриптома целых отдельных нейронов, полученных из плюрипотентных стволовых клеток человека. Front Mol Neurosci. 2018; 11: 261.
PubMed PubMed Central Статья Google Scholar
Trujillo CA, Gao R, Negraes PD, Gu J, Buchanan J, Preissl S, et al. Сложные колебательные волны, исходящие от органоидов коры, моделируют раннее развитие сети мозга человека.Стволовая клетка. 2019; 25 (4): 558–69 e7.
CAS PubMed Статья Google Scholar
Schrode N, Ho SM, Yamamuro K, Dobbyn A, Huckins L, Matos MR, et al. Синергетические эффекты распространенных вариантов риска шизофрении. Нат Жене. 2019; 51 (10): 1475–85.
CAS PubMed PubMed Central Статья Google Scholar
Ян Н., Чанда С., Марро С., Нг Ю. Х., Джанас Дж. А., Хааг Д. и др.Генерация чистых ГАМКергических нейронов путем программирования факторов транскрипции. Нат методы. 2017; 14 (6): 621–8.
CAS PubMed PubMed Central Статья Google Scholar
Борасо М., Вивиани Б. Сокультуры сэндвичей с глией-нейроном: подход in vitro для оценки межклеточной коммуникации при нейровоспалении и нейротоксичности. Методы Мол биол. 2011; 758: 135–52.
CAS PubMed Статья Google Scholar
Salaris F, Colosi C, Brighi C, Soloperto A, Turris V, Benedetti MC и др. 3D-биопечать кортикальные нейронные конструкции человека, полученные из индуцированных плюрипотентных стволовых клеток. J Clin Med. 2019; 8 (10): 1595.
CAS PubMed Central Статья Google Scholar
Tripathy SJ, Savitskaya J, Burton SD, Urban NN, Gerkin RC. НейроЭлектро: окно в данные электрофизиологии нейронов мира. Фронт нейроинформатика. 2014; 8:40.
Артикул Google Scholar
Паулл Д., Севилья А., Чжоу Х., Хан А. К., Ким Х., Наполитано С. и др. Автоматизированное высокопроизводительное получение, характеристика и дифференциация индуцированных плюрипотентных стволовых клеток. Нат методы. 2015; 12 (9): 885–92.
CAS PubMed Статья Google Scholar
Европейское действие по затратам CA16210 Максимальное воздействие исследований расстройств нервного развития (MINDDS).Доступно по адресу: https://mindds.eu/.
Томпсон Р., Джонстон Л., Тарусио Д., Монако Л., Бероуд С., Гут И.Г. и др. RD-connect: интегрированная платформа, объединяющая базы данных, реестры, биобанки и клиническую биоинформатику для исследования редких заболеваний. J Gen Intern Med. 2014; 29 (Приложение 3): S780–7.
PubMed Статья Google Scholar
Вариации количества копий – обзор
Вариации количества копий в геноме
M.fascicularisВариации числа копий ДНК (CNV) – это генетические различия в нормальной популяции, проявляющиеся как микроскопически невидимые делеции или амплификации участков геномной ДНК в диапазоне от 1 т.п.н. и до мегабазной шкалы [14]. CNV геномных сегментов обычно обнаруживаются в геномах людей [15] и модельных организмов, таких как мышь, крыса или муха Drosophila melanogaster [16]. У людей к настоящему времени идентифицировано более 2,3 миллиона различных CNV, отображающих примерно 200 000 геномных областей [17].Они вносят значительный вклад в генетическую изменчивость, охватывая большее количество нуклеотидов на геном, чем SNP (например, примерно 0,8% длины генома человека различается между двумя людьми) [18]. Поскольку CNV могут находиться в областях генома, несущих гены, очевидно, что CNV в форме гомозиготных делеций действуют как мутанты с нокаутом генов, что приводит к полной потере экспрессии и функции генов. Однако помимо этого было показано, что CNV не только влияют на экспрессию генов, различающихся по количеству копий (амплификации или гетерозиготные делеции по сравнению с эталоном), но также влияют на тканевые транскриптомы в глобальном масштабе, изменяя уровень и время экспрессии. генов, которые локализуются внутри CNV [16,19] и на его флангах [20-24], эффект, который может распространяться на всю длину пораженной хромосомы [25].Эти эффекты CNV трудно понять и не обязательно предсказуемы. Тем не менее, понимание полного влияния CNV на регуляцию экспрессии генов помогает обеспечить механистическое понимание заболеваний, связанных с CNV либо посредством дозировки одного гена, таких как чувствительность к СПИДу [26], гломерулонефрит у крыс и людей [27] и псориаз. [28], или через изменения экспрессии непрерывного набора генов, такие как синдром Вильямса-Бёрена и детские спазмы [29,30], синдром ДиДжорджи [31], синдром Смита-Магениса [32], синдром Потоцкого-Лупски [33] ] и расстройства аутистического спектра [34–36].Более того, влияние CNV на экспрессию генов дает понимание и ассоциацию индивидуальной вариабельности фармакологических ответов, с CNV CYP2D6 в качестве наиболее яркого примера. Было показано, что такие CNV вызывают у людей невосприимчивость к терапии антидепрессантами, очевидно более высокую частоту суицидов и токсичность после лечения анальгетическим кодеином [37].
Учитывая важность CNV в биомедицинских исследованиях, мы разработали первый полногеномный тайлинг-массив яванских макак, содержащий 4.2 миллиона 60-мерных зондов, которые опрашивают весь геном в среднем с интервалом в 600 пар оснований. Из сканирования генома исключены повторяющиеся области центромер и Y-хромосома, которые еще не секвенированы. В качестве эталона CNV мы использовали объединенную ДНК, выделенную от 25 самцов яванского макака маврикийского происхождения. Использование мужской ДНК в качестве базовой имеет то преимущество, что женские хромосомы могут использоваться в качестве эталона для дупликаций генов и производительности массива. На рисунке 4.1 мы показываем пример плотности тайлинг-зонда в области 60 т.п.н. на хромосоме 1 с усилением CNV и области в 90 т.п.н. на хромосоме 19 с сигналом потери CNV, оба у самца макаки из Маврикия. .Амплификация влияет на интрон гена кальпонина 3 ( CNN3 ), необходимый для слияния трофобластических клеток [38], тогда как делеция содержит весь локус, кодирующий кристаллический галектин Шарко-Лейдена ( CLC ). По аналогии с CNV для интронных регуляторных элементов трансмембранного регулятора проводимости при муковисцидозе, амплифицированный сегмент может влиять на сплайсинг транскриптов и, следовательно, на экспрессию генов [39]. Потеря CNV в этом примере, скорее всего, повлияет на уровни экспрессии гена CLC , который связан с предрасположенностью к аллергическому риниту [40].
Рисунок 4.1. Обнаружение вариаций числа копий (CNV) у одной яванской макаки с Маврикия. Область размером 60 т.п.н. на хромосоме 1 и область размером 90 т.п.н. на хромосоме 19 были выбраны в качестве примеров для обнаружения CNV у одной маврикийской яванской макаки (вверху). Схематические изображения местоположений генов и экзонов в этих геномных областях показаны в середине вместе с геномными положениями тайлинговых зондов. Зазоры, не покрытые зондами, либо повторяются, либо не уникальны, а высота стержня на зонд представляет собой увеличение потенциальной неоднозначности зонда.Сигналы CNV по сравнению с эталонным стандартом отображаются в виде отношения log2 (черные точки), показывающего усиление CNV (синий) и потери (красный), обнаруженные алгоритмом SegMNT (красные линии).
В качестве следующего шага мы исследовали CNV среди группы отдельных макак cynomolgus. Мы выбрали трех самок из Китайского центра разведения (Юго-Восточная Азия) и Маврикия. Основываясь на представлении всех точек данных на хромосоме с низким разрешением, мы указали, в качестве примера, две области CNV на хромосомах 4 и 13, которые показывают значительную потерю CNV, и одну область CNV на хромосоме 7, показывающую прирост CNV у всех людей (рис. 4.2). В том же масштабе мы обнаружили выигрыш или потерю CNV, которые происходят только у одного из лиц, которые отмечены красными треугольниками на рис. 4.2. Мы расширили этот анализ CNV на весь геном на 24 обезьяны cynomolgus и определили, что среднее количество CNV на геном составляет около 800, при средней длине около 12,5 т.п.н. на локус CNV. Эти числа вполне сопоставимы с наблюдениями, сделанными для CNV в нормальных человеческих популяциях, где большинство CNV имеют небольшие размеры (<10-30 т.п.н.) [17,41] и где любой человек несет в среднем около 670 сегментов CNV [41].Кроме того, мы всесторонне проанализировали частоту областей CNV у этих 24 обезьян и обнаружили «частные» и общие CNV (рис. 4.3). С этой целью мы обнаружили, что CNV яванского макака неслучайно распределены по геному, что указывает на концентрацию в хромосомных «горячих точках», где CNVs, вероятно, будут возникать либо de novo, либо на протяжении всей эволюции. Это открытие предполагает строгие селективные механизмы, лежащие в основе генеза или поддержания CNV.
Рисунок 4.2. Полногеномное отображение профилей CNV у cynomolgus обезьян.Синие стрелки обозначают сохраненные приросты или потери CNV, присутствующие у всех трех человек. Красные треугольники обозначают «частные» CNV. Для каждой обезьяны указано географическое происхождение. Обратите внимание на сдвиг Х-хромосомы в правом конце дисплея в результате сравнения трех самок обезьян с эталонным эталоном самцов.
Рисунок 4.3. Идеограммы участков CNV у яванских макак. На идеограммах показаны области CNV и частоты CNV для 24 яванских макак при разрешении хромосом. Обычные CNV отражаются длинными столбцами, а «частные» CNV – короткими столбцами.Прибыли CNV отмечены синим цветом, а убытки – красным.
Затем мы задали вопрос, группируются ли профили CNV в соответствии с географическим происхождением приматов. Основываясь на относительно небольшом наборе SNP, Street et al. обнаружили, что по крайней мере некоторые из этих вариаций действительно ограничиваются географическим происхождением обезьян [42]. Более актуальным для биомедицинских исследований является тот факт, что половина этих SNP являются общими с макаками-резусами [42]. Это означает, что «частные» SNP яванского макака полезны для обнаружения геномного проникновения последовательностей резуса в результате межвидовой гибридизации.Эти наблюдения позволяют предположить, что CNV как индикаторы генетической рекомбинации также будут сгруппированы в соответствии с географическим происхождением, что и произошло (рис. 4.4). Основные аутосомные профили CNV 22 из 24 обезьян cynomolgus группируются в зависимости от происхождения (Филиппины, Юго-Восточная Азия или Маврикий). Это снова означает, что хромосомные целевые локусы для CNV неслучайно распределены по всему геному и что многие локусы CNV «фиксированы» в пределах географической субпопуляции и внутри семейств макак cynomolgus, что согласуется с данными SNP.Если теперь принять во внимание, что скорость генерации SNP de novo составляет около 69 новых SNP на геном потомства у людей [43] и что скорость генерации de novo человеческих CNV составляет около 1,2 новых CNV на 100 геномов потомства [44], тогда мы можем предположить, что скорость образования CNV de novo у яванских макак довольно низка. Это, в свою очередь, привело нас к выводу, что генетическая адаптация может управлять поддержанием количества копий в этих конкретных локусах. Первоначальный случай адаптивных аллелей CNV при положительном отборе был недавно обнаружен у людей при изменении дозировки гена альфа-амилазы 1A ( AMY1A ).Этот ген кодирует белок, который катализирует первый этап переваривания диетического крахмала и гликогена. Число копий AMY1A у человека в три раза выше, чем у шимпанзе, что позволяет предположить, что людям была предпочтительна дозировка гена из-за сопутствующего увеличения потребления крахмала [45]. Таким образом, вероятно, что генетическая адаптация отдельных CNV яванского макака может влиять на уровни экспрессии генов, обеспечивая селективное преимущество для условий окружающей среды, таких как питание.Мы проверили эту гипотезу, проанализировав локус размером 2 Mb на хромосоме 19, где мы обнаружили делецию размером примерно 13 kb у двух яванских макак из Юго-Восточной Азии, которая затрагивает межгенный регион на этой хромосоме. У нас были данные микроматрицы экспрессии генов, доступные для сердца, почек, печени, легких и селезенки шести животных дикого типа для этого локуса в дополнение к двум животным с потерей числа копий. Неожиданно потеря числа копий межгенного сегмента хромосомы статистически значимо модулировала уровни экспрессии соседних генов в почках и сердце, но не в других тканях.Например, уровни мРНК активаторов транскрипции FOXA3, MEIS3, NPAS1 и SIX5 были увеличены в почках и сердце, тогда как убиквитинлигаза IRF2BP1, SUMO-активирующий фермент SAE1, каркасный белок STRN4, переносчик аминокислот SLC1A5. , белок p190RhoGAP (GRLF1), активирующий Rho-GTPase, и регулятор слияния везикул NAPA были подавлены по сравнению с здоровыми животными (рис. 4.5). Достаточно интересно, что делеция влияет на экспрессию примерно 45 генов, которые кодируются на расстоянии до 1 МБ выше и ниже этого CNV (данные не показаны).Более того, кажется, что существует тенденция понижающей модуляции генов ближе к потере числа копий, тогда как более далекие гены, по-видимому, активируются. Уровни экспрессии генов за пределами этого локуса размером 2 Mb демонстрируют относительно высокую индивидуальную вариабельность, что затрудняет интерпретацию данных. На данный момент мы можем только предполагать механизм, лежащий в основе влияния удаленного участка ДНК на всю хромосому. Такие механизмы могут включать физическую диссоциацию активаторов или репрессоров транскрипции, изменение структуры хроматина или изменение положения геномной области в ядре.
Рисунок 4.4. Географическая кластеризация по профилям CNV. Дендрограммы показывают взаимосвязь кластеров CNV, обнаруженных в аутосомах 24 отдельных обезьян cynomolgus. Пол обозначен буквами M для мужчин и F для женщин, а цветные полосы указывают на происхождение (красный = Юго-Восточная Азия; синий = Маврикий; зеленый = Филиппины).
Рисунок 4.5. Связь между CNV и экспрессией генов. Показана область размером 2 Mb на хромосоме 19, содержащая более 45 локусов генов. Две из восьми обезьян cynomolgus несут делецию размером примерно 13 килобаз в центре этой геномной области.Изменение экспрессии генов показано для генов, на которые сильно влияет CNV в носителе, по сравнению с обезьянами, не являющимися носителями, на основании данных микроматрицы для сердца и почек. Красные стрелки указывают на активацию экспрессии генов, а синие стрелки – на отрицательную. Коды отдельных обезьян cynomolgus появляются на оси и .
Точный и эффективный метод определения изменения количества копий
Мотивация: Интеграция нескольких генетических источников для обнаружения вариаций числа копий (CNV) – мощный подход к улучшению идентификации вариантов, связанных со сложными признаками.Хотя было показано, что широко используемые методы, основанные на точках изменения, могут повысить статистическую мощность для идентификации вариантов, по-прежнему сложно эффективно обнаруживать CNV со слабыми сигналами из-за зашумленной природы данных интенсивности генотипирования. Ранее мы разработали modSaRa, нормальную модель на основе среднего, основанную на алгоритме отбора и ранжирования для идентификации вариации количества копий, которая обеспечивает желаемую чувствительность с высокой вычислительной эффективностью. Чтобы повысить статистическую мощность для идентификации вариантов, здесь мы представляем новое усовершенствование, которое объединяет относительную аллельную интенсивность с внешней информацией из эмпирической статистики с моделированием, которую мы назвали modSaRa2.
Полученные результаты: Исследования моделирования показали, что modSaRa2 заметно улучшил как чувствительность, так и специфичность по сравнению с существующими методами анализа данных на основе массивов. Улучшение обнаружения слабого сигнала CNV является наиболее существенным, при этом одновременно улучшается стабильность при изменении размера CNV. Применение нового метода к полногеномному набору данных о меланоме позволило выявить новых кандидатов на делеции, связанные с риском меланомы, на полосах хромосомы 1p22.2 и дупликации в областях 6p22, 6q25 и 19p13, что может облегчить понимание возможных ролей вариантов числа копий зародышевой линии в этиологии меланомы.
Наличие и реализация: http://c2s2.yale.edu/software/modSaRa2 или https://github.com/FeifeiXiaoUSC/modSaRa2.
Дополнительная информация: Дополнительные данные доступны на сайте Bioinformatics онлайн.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
% PDF-1.5 % 799 0 объект > эндобдж xref 799 130 0000000016 00000 н. 0000005071 00000 н. 0000005238 00000 п. 0000005830 00000 н. 0000006318 00000 н. 0000007039 00000 п. 0000007527 00000 н. 0000007641 00000 п. 0000007738 00000 п. 0000008357 00000 н. 0000009034 00000 н. 0000009127 00000 н. 0000009704 00000 п. 0000010564 00000 п. 0000011350 00000 п. 0000011763 00000 п. 0000011913 00000 п. 0000011976 00000 п. 0000012413 00000 п. 0000013272 00000 п. 0000013714 00000 п. 0000013799 00000 п. 0000014173 00000 п. 0000014666 00000 п. 0000015585 00000 п. 0000015751 00000 п. 0000016063 00000 п. 0000016392 00000 п. 0000017288 00000 п. 0000017454 00000 п. 0000017569 00000 п. 0000018721 00000 п. 0000019069 00000 п. 0000020056 00000 п. 0000020932 00000 п. 0000023034 00000 п. 0000023823 00000 п. 0000023906 00000 п. 0000023961 00000 п. 0000029102 00000 п. 0000032267 00000 п. 0000034093 00000 п. 0000037019 00000 п. 0000040871 00000 п. 0000040995 00000 п. 0000041111 00000 п. 0000041189 00000 п. 0000041382 00000 п. 0000041460 00000 п. 0000041653 00000 п. 0000041731 00000 п. 0000044232 00000 п. 0000044310 00000 п. 0000046859 00000 п. 0000046937 00000 п. 0000049239 00000 п. 0000049317 00000 п. 0000051978 00000 п. 0000052056 00000 п. 0000054688 00000 п. 0000054766 00000 п. 0000057482 00000 п. 0000057560 00000 п. 0000060204 00000 п. 0000060282 00000 п. 0000062813 00000 п. 0000062891 00000 п. 0000065413 00000 п. 0000065491 00000 п. 0000068014 00000 п. 0000068092 00000 п. 0000068285 00000 п. 0000068363 00000 п. 0000070886 00000 п. 0000070964 00000 п. 0000073494 00000 п. 0000073572 00000 п. 0000076087 00000 п. 0000076165 00000 п. 0000078804 00000 п. 0000078882 00000 п. 0000081399 00000 н. 0000081477 00000 п. 0000083993 00000 п. 0000084071 00000 п. 0000086597 00000 п. 0000086675 00000 п. 0000089193 00000 п. 0000089271 00000 п. 0000091785 00000 п. 0000091863 00000 п. 0000094524 00000 п. 0000094602 00000 п. 0000094795 00000 п. 0000094873 00000 п. 0000097483 00000 п. 0000097561 00000 п. 0000100099 00000 н. 0000100177 00000 н. 0000102753 00000 п. 0000102831 00000 н. 0000105341 00000 п. 0000105419 00000 п. 0000107934 00000 п. 0000108012 00000 н. 0000108205 00000 н. 0000108283 00000 н. 0000108476 00000 п. 0000108554 00000 п. 0000108747 00000 н. 0000108825 00000 н. 0000109017 00000 н. 0000109095 00000 н. 0000111712 00000 н. 0000111790 00000 н. 0000114321 00000 н. 0000114552 00000 н. 0000114591 00000 н. 0000121855 00000 н. 0000121953 00000 н. 0000122075 00000 н. 0000122201 00000 н. 0000122999 00000 н. 0000123038 00000 н. 0000123836 00000 н. 0000123875 00000 н.