Б п никитина сложи узор: Кубики Никитина Сложи Узор, развивающая дидактическая игра с авторскими заданиями, деревянные чудо-кубики Никитина
Развивающие игровые технологии кубики Б.П.Никитина “Сложи узор” | Статья:
1.Кубики Б.П.Никитина – одна из популярных игровых технологий для детей дошкольного возраста.
Как известно, первые годы жизни ребенка- самые ценные для его будущего и надо как можно полнее использовать их. В дошкольном детстве происходит становление первых форм абстракции, обобщение простых умозаключений, переход от практического мышления к логическому. Происходит развитие восприятия, внимания, памяти, воображения.
Как сказал Б.П.Никитин: « Игрушки, игры – одно из самых сильных воспитательных средств в руках обществаВы думаете, что просто покупаете игрушку? Нет, вы проектируете при этом человеческую личность!»
2.Никитин – педагог-новатор, основоположник многих развивающих игр с кубиками. Таких как:
|
|
3. Кубики Никитина – это прекрасные и увлекательные пособия для развития интеллектуальных способностей детей, в которых всегда есть один простой базовый набор: кубики – из которых необходимо сложить узор; рамки – вкладыши или геометрические фигуры. С их помощью, можно создавать различные задания для малышей разного уровня сложности.
Кубики Никитина – это прекрасные и увлекательные пособия для развития интеллектуальных способностей детей, в которых всегда есть один простой базовый набор: кубики – из которых необходимо сложить узор; рамки – вкладыши или геометрические фигуры. С их помощью, можно создавать различные задания для малышей разного уровня сложности.
4. При игре в кубики, необходимо придерживаться основных правил:
- Сначала взрослый, потом ребенок (выполняя задания вы сможете сами прочувствовать с какими сложностями столкнется ребенок выполняя их)
- Подсказки давать запрещено
- На решение задачи ребенку нужно давать столько времени, сколько требуется
- Если ребенок не справляется, делайте вынужденные перерывы, возвращаясь к простым вариантам
5. Сегодня я познакомлю вас с игрой «Сложи узор». Эту игру очень любят дети, и не удивительно.
6. Из этой маленькой коробочки, рождаются красивые узоры:
7. Буквы
8. Цифры
9. Симпатичные животные и много всего интересного. А самое главное создают все это сами дети.
А самое главное создают все это сами дети.
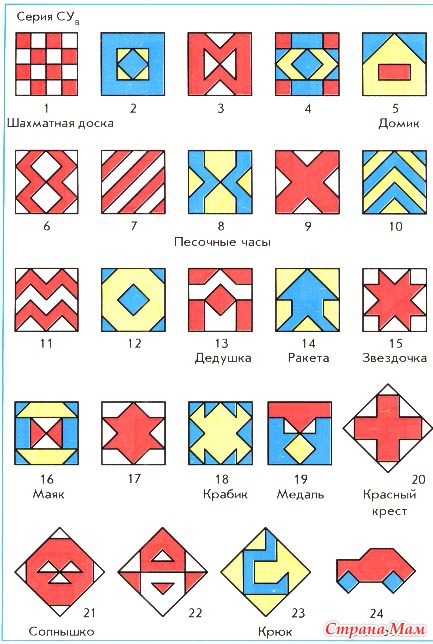
У вас на столе лежит игра СУ. Достаньте кубики, рассмотрите их. Игра состоит из 16 одинаковых кубиков. Каждый кубик окрашен в 4 цвета:
|
|
|
|
и две грани кубика двуцветные: бело-красные и сине-желтые. Из этих кубиков можно сложить огромное количество разных интересных узоров.
10.Задания для игры делятся по количеству использованных кубиков :
На 1 этапе – задания из 4-х кубиков.
На 2 этапе –из 9-ти кубиков.
НА 3 этапе –из 16-ти кубиков с прорисовкой, а в последствии без прорисовки границ рисунка.
Таким образом возрастает сложность заданий от этапа к этапу. В этом и заключается принцип от «простого к сложному», когда предыдущее задание подготавливает малыша к следующему. Все это в совокупности дает возможность ребенку находить решение самостоятельно, без подсказки.
Все это в совокупности дает возможность ребенку находить решение самостоятельно, без подсказки.
В настоящее время разработаны различные пособия и альбомы, которые помогают педагогу в организации игр с кубиками.
Условно организацию игры можно разделить на 3 этапа.
1 этап — МЛАДШАЯ ГРУППА. На данном этапе я использую задания из альбома «СЛОЖИ УЗОР для МАЛЫШЕЙ»
11.У малышей наглядно-образное мышление, поэтому схемы в альбоме «оживают» и кубики превращаются в бабочек, цветочки, зверят.
– Сколько бабочек из кубиков ты видишь на картинке? (4)
– Чем похожи бабочки? (У всех бабочек крылышки квадратные)
Чем отличаются ? (Крылышки разного цвета)
12. В задании можно предложить малышу отгадать загадку:
-Этот фрукт на вкус хорош и на лампочку похож? (груша)
-Круглое, румяное я расту на ветке, любят меня взрослые
и маленькие детки ( яблоко)
Дети отгадывают загадки и подбирают кубик по цвету.
13. Нужно помочь мышонку добраться до сыра
Малыш выкладывает дорожку заданного цвета и просчитывает количество выложенных кубиков.
14. Предлагается ребенку развесить на деревья желтые и красные листочки. Строим для ежика дорожку
С помощью таких заданий –мы учим и закрепляем цвета; формируем представления о временах года, формируем начальные навыки счета.
15. Постепенно сложность задания возрастает. Задания из одноцветных кубиков, переходят в задания двуцветные.
– У медвежонка и котенка тихий час. Укрой их одеяльцами.
Как ты думаешь, чем похожи одеяльца? А чем отличаются?
Покажи на рисунке большие и маленькие треугольники
16. Выполняя такие задания, дети усваивают понятия закономерности и последовательности. 2 этапа игры СУ рассчитан на детей средней группы. В работе с детьми данного возраста, использую задания из альбома «Чудо-Кубики».
17. Учитывая принцип от простого к сложному, задания 2 этапа усложняются, состоят из 9-ти кубиков.
Предлагается выложить узор из кубиков на одеяльца, как показано на картинке, а затем постараться построить свой узор.
18. К примеру поменять цвета на одеяльцах.
К примеру поменять цвета на одеяльцах.
19. Задания данного альбома учат детей симметрично выкладывать узоры на картинке.
Составь из кубиков правую часть ковра симметрично левой.
20. Тренируем внимание. Найди на рисунке такой же корабль и выложи его.
21. Попробуй собрать и остальные корабли.
22. На 3 этапе игры, использую альбом «Чудо-Кубики -2» для детей старшей и подготовительной групп.
Задания – картинки в этом альбоме состоят из 16-ти кубиков с прорисованными гранями. В дальнейшем задания усложняются и картинки предлагаются без прорисовки границ.
23. Детям очень нравиться собирать картинки и составлять узоры из кубиков. Каждый раз выполняя новое задание, радуются что у них получилось справиться с ним, что они смогли его выполнить самостоятельно, без подсказки взрослого.
24. Маргарита с сердечком
25. Ира с лебедем.
Кубики Никитина универсальны. Один и тот же набор кубиков можно использовать на протяжении всего дошкольного детства и в начальной школе, постепенно усложняя игру и задания к ней.
Они могут использоваться в коррекционной и диагностической работе.
Плюс данной технологии еще и в том, что она развивает:
– наглядно действенное мышление
– мыслительные операции: сравнения, анализа, синтеза
– цветовое восприятие
– самоконтроль и самоанализ
– развивает- зрительную память, мелкую моторику, чувство симметрии и т.д.
Кубики Никитина Сложи узор – Развивающая игра известного педагога Б. П. Никитина. В комплект входят 16 одинаковых пластиковых кубиков в картонной коробке, методические рекомендации
Новосибирск
| Компании: | 19 864 |
| Товары и услуги: | 12 990 |
| Статьи и публикации: | 1 143 |
| Тендеры и вакансии: | 114 |
Вход в личный кабинет
А ваша компания есть в справочнике?
- Компании
- Товары и услуги
- Тендеры
- Вакансии
- Статьи и публикации
480 р.
Купить
Развивающая игра известного педагога Б. П. Никитина. В комплект входят 16 одинаковых пластиковых кубиков в картонной коробке, методические рекомендации
Развивающая игра известного педагога Б.П. Никитина. В комплект входят 16 одинаковых пластиковых кубиков в картонной коробке, методические рекомендации родителям и педагогам, примеры узоров. Каждая грань кубика имеет разные цвета, что позволяет составлять множество разнообразных силуэтов и узоров. Кубики помогут развить у детей пространственное воображение, сообразительность и логическое мышление, навыки счета и графические способности, цветоощущение, умение анализировать, синтезировать и комбинировать, навыки классификации, а также воспитать аккуратность, внимание, точность, усидчивость, целеустремленность. Возраст: 2 – 7лет. Производитель: Россия
посмотреть все (248)
Другие товары и услуги компании:
Кубики «Герои сказок» 9 шт.
230 р.
Кубики Сложи узор
Развивающая игра известного педагога Б. П. Никитина
480 р.
Сложи квадрат, 2-я категория сложности. Пособие Никитина
Авторская головоломка Б. Никитина. Два планшета размером А5 с квадратами, по-разному разделенными на 3, 4, и 5 частей
330 р.
Кубики Зайцева
“Кубики Зайцева” — пособие для обучения чтению с двух лет и может использоваться в семье, детском саду и школе.
4 500 р.
Уникуб

590 р.
Логический кубик малый
Логический кубик по Сегену для самых маленьких. Здесь надо вставить всего по одной фигуре с каждой стороны, но для годовалых малышей и этого уже достаточно
115 р.
Товары и услуги других компаний:
Конструктор “Цветной” (65 деталей)
Конструктор “Цветной” (65 деталей)
450 р.
- Семья, дети
Игрушки и игры
Информация о продавце
Игры Почемучек, магазин развивающих игрушек и настольных игр
- +7 (913) 382-59-26
- г. Новосибирск, ул. Красный проспект, 155 (пл. Калинина)
- igry-pochemuchek.ru
Развивающие игры, творчество, настольные игры, игры для всей семьи, учимся играя, игрушки, развивающие игры в Новосибирске, настольные игры в Новосибирске
Татьяна Никитина | Массачусетский университет в Амхерсте | 14 публикаций | 1755 Цитаты
疟原 虫 var 基因 转换 速率 变化 导致 抗原 变异 变异 [英] / Пол Х, Роберт П. , Христодулу З. и др. // Proc Natl Acad Sci U S A
, Христодулу З. и др. // Proc Natl Acad Sci U S A
[…]
宁北芳, 朱淮民 朱淮民
28 июля 2005
TL; DR: PFPMP1) 感染 红 细胞 、 树突 状 组胞 以及 胎盘 的 或 多 个 作用 , 在 黏附 及 免疫 中 起 关键 作 � �ly.
… Прочитайте Moreread Less
Аннотация: 抗原 变异 可 多 种 致病 微 生物 易于 逃避 宿主 免疫 应答。 在 感染 细胞 表面 易于 逃避 宿主 宿主 免疫。 表达 表达 感染 红 表面 恶性 疟原 虫 虫 表面 蛋白 1 (pfpmp1) 感染 、 、 、 红细胞 表面树突状 细胞 胎盘 的 单个 或 多 个 受体 作用 , 在 黏附 免疫 逃避 中 起 关键 的 作用。 个 单倍体 基因组 var 基因 编码 约 60 种 成员 通过 启动 不同 的 var 基因 变异体 约 种 成员 通过 启动 不同 的 的 基因 变异体 变异体 约 种 通过 启动 转录 不同 的 的为抗原变异提供了分子基础。
…читать дальшеЧитать меньше
18 940 цитирований
Журнальная статья•DOI•
Шелтерин: белковый комплекс, формирующий и защищающий теломеры человека
[…]
Титиа де Ланге 1)
Рокфеллеровский университет 1
15 сентября 2005 г.-Genes & Development
TL;DR: Текущие данные утверждают, что шелтерин появляется как белковый комплекс с активностью ремоделирования ДНК, который действует вместе с несколькими ассоциированными факторами репарации ДНК, изменяют структуру теломерной ДНК, тем самым защищая концы хромосом.
…читать дальшечитать меньше
Резюме: Массивы повторов TTAGGG, добавленные теломеразой, определяют концы хромосом человека. Комплекс, образованный шестью специфичными для теломер белками, связывается с этой последовательностью и защищает концы хромосом. По аналогии с другими хромосомными белковыми комплексами, такими как конденсин и когезин, я буду называть этот комплекс шелтерином. Три субъединицы шелтерина, TRF1, TRF2 и POT1, непосредственно распознают повторы TTAGGG. Они связаны между собой тремя дополнительными белками шелтерина, TIN2, TPP1 и Rap1, образуя комплекс, позволяющий клеткам отличать теломеры от участков повреждения ДНК. Без защитной активности шелтерина теломеры больше не скрыты от наблюдения за повреждением ДНК, а концы хромосом неправильно обрабатываются путями репарации ДНК. Как шелтерин предотвращает эти события? Текущие данные утверждают, что шелтерин не является статическим структурным компонентом теломер. Вместо этого шелтерин появляется как белковый комплекс с активностью ремоделирования ДНК, который действует вместе с несколькими ассоциированными факторами репарации ДНК, изменяя структуру теломерной ДНК, тем самым защищая концы хромосом. Шесть субъединиц шелтерина: TRF1, TRF2, TIN2, Rap1, TPP1 и POT1.
Шесть субъединиц шелтерина: TRF1, TRF2, TIN2, Rap1, TPP1 и POT1.
…читать дальшеЧитать меньше
2515 цитирований
Журнальная статья•DOI•
Бисульфитное секвенирование генома арабидопсиса методом дробовика выявило паттерн метилирования ДНК
[…]
1 Shawn J. Feng 2 , Xiaoyu Zhang 3 , Zugen Chen 1 , Barry Merriman 1 , Кристиан Д. Хауденшильд 4 , Sriharsa Pradhan 5 , STANLES F. NELSOSON , , , , , , , , , , , , , , , , , , , , . Якобсен 2 – Показать меньше еще +6•Учреждения (5)Калифорнийский университет, Лос-Анджелес 1 , Медицинский институт Говарда Хьюза 2 , Университет Джорджии 3 , Illumina 4 , 90 New England Biolabs 5
13 марта 2008-Nature
TL;DR: Карта метилированных цитозинов арабидопсиса с разрешением по одной паре оснований создана путем сочетания бисульфитной обработки геномной ДНК со сверхвысокопроизводительным секвенированием с использованием Illumina 1G Genome. Анализатор и технология секвенирования Solexa.
Анализатор и технология секвенирования Solexa.
…читать дальшечитать меньше
Резюме: Цитозиновое метилирование ДНК играет важную роль в регуляции экспрессии генов и подавлении транспозонов и других повторяющихся последовательностей. Недавние геномные исследования Arabidopsis thaliana показали, что многие эндогенные гены метилированы либо в пределах своих промоторов, либо в пределах своих транскрибируемых областей, и что метилирование генов сильно коррелирует с уровнями транскрипции. Однако растения имеют разные типы метилирования, контролируемые разными генетическими путями, и подробная информация о статусе метилирования каждого цитозина в любом данном геноме отсутствует. С этой целью мы создали карту метилированных цитозинов арабидопсиса с разрешением по одной паре оснований, объединив обработку бисульфитом геномной ДНК с высокопроизводительным секвенированием с использованием анализатора генома Illumina 1G и технологии секвенирования Solexa. Этот подход, названный BS-Seq, в отличие от предыдущих методов на основе микрочипов, позволяет чувствительно измерять метилирование цитозина в масштабе всего генома в определенных контекстах последовательностей. Здесь мы описываем метилирование ранее недоступных компонентов генома и анализируем состав и распределение последовательности метилирования ДНК. Мы также описываем влияние различных мутантов метилирования ДНК на паттерны метилирования всего генома и демонстрируем, что наша недавно разработанная конструкция библиотеки и вычислительные методы могут быть применены к большим геномам, таким как геном мыши.
Этот подход, названный BS-Seq, в отличие от предыдущих методов на основе микрочипов, позволяет чувствительно измерять метилирование цитозина в масштабе всего генома в определенных контекстах последовательностей. Здесь мы описываем метилирование ранее недоступных компонентов генома и анализируем состав и распределение последовательности метилирования ДНК. Мы также описываем влияние различных мутантов метилирования ДНК на паттерны метилирования всего генома и демонстрируем, что наша недавно разработанная конструкция библиотеки и вычислительные методы могут быть применены к большим геномам, таким как геном мыши.
… Прочитайте Moreread Less
1 925 Цитации
Журнал. 1)
Рокфеллеровский университет 1
04 ноября 2008 г. – Ежегодный обзор генетики
TL;DR: Недавние эксперименты показали, как шелтерин подавляет сигнальные пути киназы ATM и ATR и скрывает концы хромосом от негомологичного соединения концов и гомологии- адресный ремонт.
…читать дальшечитать меньше
Резюме: Геномы прокариот и эукариотических органелл обычно кольцевые, как и большинство плазмид и вирусных геномов. Напротив, ядерные геномы эукариот организованы на линейных хромосомах, что требует механизмов для защиты и репликации концов ДНК. Эукариоты решают эти проблемы с появлением теломер, защитных нуклеопротеиновых комплексов на концах линейных хромосом и теломеразы, фермента, поддерживающего ДНК в этих структурах. Теломеры млекопитающих содержат специфический белковый комплекс, шелтерин, который защищает концы хромосом от всех аспектов реакции на повреждение ДНК и регулирует поддержание теломер теломеразой. Недавние эксперименты, обсуждаемые здесь, показали, как шелетин репрессирует пути передачи сигналов киназы ATM и ATR и скрывает концы хромосом от негомологичного соединения концов и направленной по гомологии репарации.
…читать дальшеЧитать меньше
1609 цитирований
Журнальная статья•DOI•
Теломеры и болезни человека: старение, рак и другие болезни
[. ..]
..]
Мария А. Бласко
01 августа Nature Reviews Genetics
TL;DR: изменение функционирования как теломеразы, так и белков, взаимодействующих с теломерами, наблюдается при некоторых синдромах преждевременного старения человека и при раке, а недавние результаты показывают, что изменения, влияющие на теломеры на уровне структуры хроматина, также могут иметь роль в заболеваниях человека.
…читать дальшечитать меньше
Резюме: Длина теломер и активность теломеразы являются важными факторами патобиологии заболеваний человека. Возрастные заболевания и синдромы преждевременного старения характеризуются короткими теломерами, которые могут поставить под угрозу жизнеспособность клеток, в то время как опухолевые клетки могут предотвращать потерю теломер за счет аномальной активизации теломеразы. Измененное функционирование как теломеразы, так и белков, взаимодействующих с теломерами, присутствует при некоторых синдромах преждевременного старения человека и при раке, а недавние открытия показывают, что изменения, влияющие на теломеры на уровне структуры хроматина, также могут играть роль в заболеваниях человека. Эти результаты вдохновили на разработку ряда потенциальных терапевтических стратегий, основанных на теломеразе и теломерах.
Эти результаты вдохновили на разработку ряда потенциальных терапевтических стратегий, основанных на теломеразе и теломерах.
…читать дальшечитать меньше
1,434 цитирования
Расстояние между нуклеосомами периодически модулирует укладку нуклеосомной цепи и топологию ДНК в кольцевых массивах нуклеосом
1. Джоти Ю., Хикима Т., Нишино Ю., Камада Ф., Хихара С., Таката Х., Исикава Т. и Маэсима К. (2012) Хромосомы без 30-нм хроматинового волокна. ядро 3, 404–410 10.4161/нукле.21222 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2. Оу Х. Д., Фан С., Диринк Т. Дж., Тор А., Эллисман М. Х. и О’Ши К. С. (2017) ChromEMT: визуализация трехмерной структуры хроматина и уплотнения в интерфазных и митотических клетках. Наука 357, eaag0025 10.1126/наука.aag0025 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3.
Hsieh T.H., Weiner A., Lajoie B., Dekker J., Friedman N. и Rando O.J. (2015) Картирование укладки хромосом с разрешением нуклеосом в дрожжах с помощью Micro-C.
4. Григорьев С. А., Баском Г., Баквалтер Дж. М., Шуберт М. Б., Вудкок С. Л. и Шлик Т. (2016) Иерархическое зацикливание зигзагообразных цепей нуклеосом в метафазных хромосомах. проц. Натл. акад. науч. США 113, 1238–1243 гг. 10.1073/пнас.1518280113 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Риска В. И., Денни С. К., Стрейт А. Ф. и Гринлиф В. Дж. (2017) Вариабельная структура хроматина, выявленная с помощью in situ пространственно коррелированного картирования расщепления ДНК. Природа 541, 237–241 10.1038/природа20781 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Кай С., Бок Д., Пилхофер М. и Ган Л. (2018) in situ структур моно-, ди- и тринуклеосом в гетерохроматине человека. Мол. биол. Клетка 29, 2450–2457 гг. 10.1091/mbc.E18-05-0331 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Вудкок С.Л., Григорьев С.А., Горовиц Р.А. и Уитакер Н. (1993)Модель укладки хроматина, которая включает изменчивость линкера, генерирует волокна, напоминающие нативные структуры. проц. Натл. акад. науч. США 90, 9021–9025
10.1073/пнас.90.19.9021
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Вудкок С.Л., Григорьев С.А., Горовиц Р.А. и Уитакер Н. (1993)Модель укладки хроматина, которая включает изменчивость линкера, генерирует волокна, напоминающие нативные структуры. проц. Натл. акад. науч. США 90, 9021–9025
10.1073/пнас.90.19.9021
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Леуба С.Х., Ян Г., Роберт С., Самори Б., ван Холде К., Златанова Дж. и Бустаманте С. (1994) Трехмерная структура протяженных хроматиновых волокон, выявленная с помощью сканирующей силовой микроскопии в постукивающем режиме. проц. Натл. акад. науч. США 91, 11621–11625 10.1073/пнас.91.24.11621 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Григорьев С. А. (2018) Укладка хроматина более высокого порядка: перспектива с углами линкерной ДНК. Биофиз. Дж. 114, 2290–2297. 10.1016/j.bpj.2018.03.009 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10.
Перишич О., Коллепардо-Гевара Р. и Шлик Т. (2010) Моделирование исследования структуры волокон хроматина в зависимости от длины линкера ДНК. Дж. Мол. биол. 403, 777–802
10.1016/j.jmb.2010.07.057
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Мол. биол. 403, 777–802
10.1016/j.jmb.2010.07.057
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Видом Дж. (1992) Связь между спиральным поворотом ДНК и упорядоченным расположением нуклеосом во всех эукариотических клетках. проц. Натл. акад. науч. США 89, 1095–1099 гг. 10.1073/пнас.89.3.1095 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Лор Д. (1986) Зависимость структуры хроматина курицы и дрожжей от соли. Влияние на межнуклеосомную организацию и отношение к активному хроматину. Дж. Биол. хим. 261, 9904–9914 [PubMed] [Академия Google]
13. Wang J.P., Fondufe-Mittendorf Y., Xi L., Tsai G.F., Segal E. и Widom J. (2008) Преимущественно квантованные длины линкерной ДНК в Saccharomyces cerevisiae . PLoS-компьютер. биол. 4, е1000175 10.1371/journal.pcbi.1000175 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14.
Брогаард К., Си Л., Ван Дж. П. и Видом Дж. (2012) Карта положений нуклеосом в дрожжах с разрешением пар оснований. Природа
486, 496–501
10.1038/природа11142
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Природа
486, 496–501
10.1038/природа11142
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Череджи Р.В., Рамачандран С., Брайсон Т.Д. и Хеникофф С. (2018) Точное полногеномное картирование одиночных нуклеосом и линкеров in vivo . Геном биол. 19, 19 10.1186/с13059-018-1398-0 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Вонг Л. Н., Си Л., Себесон А. С., Сюн Б., Ван Дж. П. и Ван Х. (2016) Изучение организации нуклеосом в эмбриональных стволовых клетках мыши с помощью химического картирования. Клетка 167, 1555–1570.e15 10.1016/j.cell.2016.10.049[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Валуев А., Джонсон С.М., Бойд С.Д., Смит С.Л., Файр А.З. и Сидоу А. (2011)Детерминанты организации нуклеосом в первичных клетках человека. Природа 474, 516–520 10.1038/природа10002 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18.
Балди С., Кребс С., Блюм Х. и Беккер П. Б. (2018) Полногеномное измерение регулярности и расстояния между локальными массивами нуклеосом с помощью секвенирования нанопор. Нац. Структура Мол. биол. 25, 894–901
10.1038/с41594-018-0110-0
[PubMed] [CrossRef] [Google Scholar]
Нац. Структура Мол. биол. 25, 894–901
10.1038/с41594-018-0110-0
[PubMed] [CrossRef] [Google Scholar]
19. Лоуари П. Т. и Видом Дж. (1998) Новые правила последовательности ДНК для высокоаффинного связывания с октамером гистонов и ориентированного на последовательность позиционирования нуклеосом. Дж. Мол. биол. 276, 19–42 10.1006/jmbi.1997.1494 [PubMed] [CrossRef] [Google Scholar]
20. Робинсон П. Дж., Файрал Л., Хюинь В. А. и Родс Д. (2006) ЭМ-измерения определяют размеры хроматинового волокна «30 нм»: свидетельство компактной встречно-штыревой структуры. проц. Натл. акад. науч. США 103, 6506–6511 10.1073/пнас.0601212103 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Routh A., Sandin S. и Rhodes D. (2008)Длина нуклеосомных повторов и стехиометрия линкерных гистонов определяют структуру волокон хроматина. проц. Натл. акад. науч. США 105, 8872–8877 10.1073/пнас.0802336105 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Вонг Х., Виктор Дж. М. и Моцциконаччи Дж. (2007) Полноатомная модель хроматинового волокна, содержащего линкерные гистоны, обнаруживает универсальную структуру, регулируемую длиной нуклеосомных повторов. PLoS один
2, е877
10.1371/journal.pone.0000877
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Вонг Х., Виктор Дж. М. и Моцциконаччи Дж. (2007) Полноатомная модель хроматинового волокна, содержащего линкерные гистоны, обнаруживает универсальную структуру, регулируемую длиной нуклеосомных повторов. PLoS один
2, е877
10.1371/journal.pone.0000877
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Коллепардо-Гевара Р. и Шлик Т. (2014)Полиморфизм волокон хроматина, вызванный вариациями длин линкеров ДНК. проц. Натл. акад. науч. США 111, 8061–8066 10.1073/пнас.1315872111 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Коррелл С. Дж., Шуберт М. Х. и Григорьев С. А. (2012) Короткие нуклеосомные повторы накладывают вращательные модуляции на укладку хроматиновых волокон. EMBO J. 31, 2416–2426 10.1038/эмбодж.2012.80 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
25. Норузи Д., Журкин В.Б. (2015) Топологический полиморфизм двухстартового хроматинового волокна. Биофиз. Дж. 108, 2591–2600. 10.1016/j.bpj.2015.04.015 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Баквалтер Дж. М., Норузи Д., Арутюнян А., Журкин В. Б. и Григорьев С. А. (2017) Регуляция укладки хроматина конформационными вариациями нуклеосомной линкерной ДНК. Нуклеиновые Кислоты Res. 45, 9372–9387
10.1093/нар/gkx562
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Баквалтер Дж. М., Норузи Д., Арутюнян А., Журкин В. Б. и Григорьев С. А. (2017) Регуляция укладки хроматина конформационными вариациями нуклеосомной линкерной ДНК. Нуклеиновые Кислоты Res. 45, 9372–9387
10.1093/нар/gkx562
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
27. Никитина Т., Норузи Д., Григорьев С. А., Журкин В. Б. (2017) Топология ДНК в хроматине определяется расстоянием между нуклеосомами. науч. Доп. 3, е1700957 10.1126/sciadv.1700957 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Комптон Дж. Л., Беллард М. и Чамбон П. (1976) Биохимические доказательства изменчивости длины повторов ДНК в хроматине высших эукариот. проц. Натл. акад. науч. США 73, 4382–4386 10.1073/пнас.73.12.4382 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29.
Лор Д., Корден Дж., Татчелл К., Ковачич Р.Т. и Ван Холде К.Е. (1977) Сравнительная субъединичная структура хроматина HeLa, дрожжей и куриных эритроцитов. проц. Натл. акад.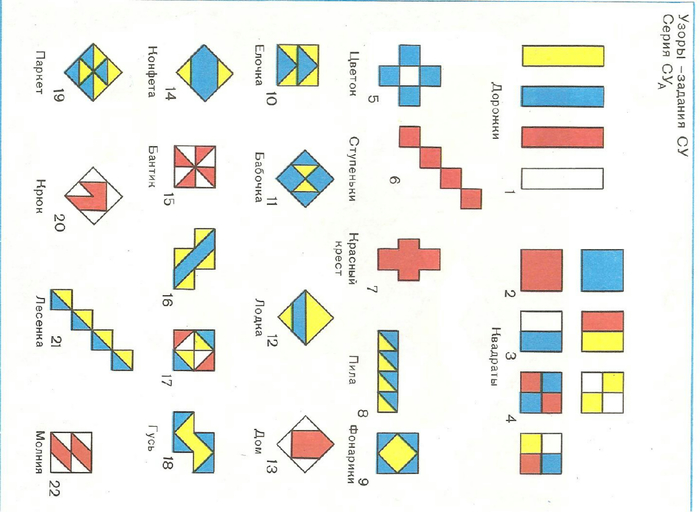 науч. США 74, 79–83
10.1073/пнас.74.1.79
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
науч. США 74, 79–83
10.1073/пнас.74.1.79
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Бауэр В. Р., Крик Ф. Х. и Уайт Дж. Х. (1980) Сверхспиральная ДНК. науч. Являюсь. 243, 100–113 [PubMed] [Google Scholar]
31. Шур М., Пуллибланк Д.Э. и Виноград Дж. (1977) Проблемы упаковки эукариотической и прокариотической ДНК и конформации in vivo, возникающие из-за неоднородности плотности суперспирали. Нуклеиновые Кислоты Res. 4, 1183–1205 гг. 10.1093/нар/4.5.1183 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Норузи Д. и Журкин В. Б. (2018) Динамика хроматиновых волокон: сравнение моделирования методом Монте-Карло с силовой спектроскопией. Биофиз. Дж. 115, 1644–1655 гг. 10.1016/j.bpj.2018.06.032 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33.
Шварц П.М., Фелтхаузер А., Флетчер Т.М. и Хансен Дж. К. (1996) Обратимая самоассоциация олигонуклеосом: зависимость от двухвалентных катионов и основных доменов гистонового хвоста. Биохимия
35, 4009–4015
10.1021/bi9525684
[PubMed] [CrossRef] [Google Scholar]
Биохимия
35, 4009–4015
10.1021/bi9525684
[PubMed] [CrossRef] [Google Scholar]
34. Григорьев С. А., Арья Г., Коррелл С., Вудкок С. Л. и Шлик Т. (2009) Данные о гетероморфных хроматиновых волокнах на основе анализа нуклеосомных взаимодействий. проц. Натл. акад. науч. США 106, 13317–13322 10.1073/пнас.0903280106 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Виан Л., Пковска А., Рао С.С.П., Киффер-Квон К.Р., Юнг С., Баранелло Л., Хуанг С.К., Эль Хаттаби Л., Дозе М., Пруэтт Н., Санборн А.Л., Канела А., Маман Ю. , Оксанен А., Реш В. и соавт. (2018) Энергетика и физиологическое воздействие экструзии когезина. Клетка 173, 1165–1178 гг. е1120 10.1016/j.cell.2018.03.072 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Yan Y., Ding Y., Leng F., Dunlap D. и Finzi L. (2018)Опосредованные белками петли в сверхскрученной ДНК создают большие топологические домены. Нуклеиновые Кислоты Res. 46, 4417–4424 10.1093/нар/gky153 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Филлипс-Креминс Дж. Э., Саурия М. Э., Саньял А., Герасимова Т. И., Лажуа Б. Р., Белл Дж. С., Онг С. Т., Хукуэй Т. А., Го К., Сунь Ю., Бланд М. Дж., Вагстафф В., Далтон С., Макдевитт Т. С., Сен Р., Деккер Дж., Тейлор Дж. и Корсес В.Г. (2013) Подклассы архитектурных белков формируют трехмерную организацию геномов во время приверженности линии. Клетка
153, 1281–1295
10.1016/j.cell.2013.04.053
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Филлипс-Креминс Дж. Э., Саурия М. Э., Саньял А., Герасимова Т. И., Лажуа Б. Р., Белл Дж. С., Онг С. Т., Хукуэй Т. А., Го К., Сунь Ю., Бланд М. Дж., Вагстафф В., Далтон С., Макдевитт Т. С., Сен Р., Деккер Дж., Тейлор Дж. и Корсес В.Г. (2013) Подклассы архитектурных белков формируют трехмерную организацию геномов во время приверженности линии. Клетка
153, 1281–1295
10.1016/j.cell.2013.04.053
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Кулаева О. И., Низовцева Е. В., Поликанов Ю. С., Ульянов С. В., Студицкий В. М. (2012) Дистанционная активация транскрипции: механизмы действия энхансеров. Мол. Клетка. биол. 32, 4892–4897 10.1128/МКБ.01127-12 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Люгер К. и Хансен Дж. К. (2005)Динамика нуклеосом и хроматиновых волокон. Курс. мнение Структура биол. 15, 188–196 10.1016/ж.сби.2005.03.006 [PubMed] [CrossRef] [Google Scholar]
40.
Треметик Д. Дж. (2007) Структуры хроматина более высокого порядка: неуловимое 30-нм волокно. Клетка
128, 651–654
10.1016/j.cell.2007.02.008
[PubMed] [CrossRef] [Google Scholar]
Клетка
128, 651–654
10.1016/j.cell.2007.02.008
[PubMed] [CrossRef] [Google Scholar]
41. Шлик Т., Хейс Дж. и Григорьев С. (2012) На пути к сближению экспериментальных исследований и теоретического моделирования хроматинового волокна. Дж. Биол. хим. 287, 5183–5191 10.1074/jbc.R111.305763 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Шальх Т., Дуда С., Сарджент Д. Ф. и Ричмонд Т. Дж. (2005) Рентгеновская структура тетрануклеосомы и ее значение для хроматинового волокна. Природа 436, 138–141 10.1038/природа03686 [PubMed] [CrossRef] [Google Scholar]
43. Сонг Ф., Чен П., Сунь Д., Ван М., Донг Л., Лян Д., Сюй Р. М., Чжу П. и Ли Г. (2014) Крио-ЭМ исследование хроматинового волокна выявляет двойную спираль скручены тетрануклеосомными единицами. Наука 344, 376–380 10.1126/наука.1251413 [PubMed] [CrossRef] [Академия Google]
44.
Экундайо Б., Ричмонд Т.Дж. и Шальх Т. (2017) Учет структурной неоднородности волокон хроматина. Дж. Мол. биол. 429, 3031–3042
10.1016/j.jmb.2017.09.002
[PubMed] [CrossRef] [Google Scholar]
429, 3031–3042
10.1016/j.jmb.2017.09.002
[PubMed] [CrossRef] [Google Scholar]
45. Низовцева Е. В., Клаувелин Н., Тодолли С., Поликанов Ю. С., Кулаева О. И., Венгжинек С., Олсон В. К., Студицкий В. М. (2017) Свободные от нуклеосом участки ДНК по-разному влияют на отдаленные коммуникации в хроматине. Нуклеиновые Кислоты Res. 45, 3059–3067 10.1093/нар/gkw1240 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Воган Р. М., Диксон Б. М., Уелихан М. Ф., Джонстон А. Л., Корнетт Э. М., Чик М. А., Аушерман К. А., Коулз М. В., Сан З. В. и Ротбарт С. Б. (2018) Структура хроматина и его химические модификации регулируют селективность субстрата убиквитинлигазы UHRF1. проц. Натл. акад. науч. США 115, 8775–8780 10.1073/пнас.1806373115 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47.
Бирнбойм Х.С. и Доли Дж. (1979) Процедура быстрой щелочной экстракции для скрининга рекомбинантной плазмидной ДНК. Нуклеиновые Кислоты Res. 7, 1513–1523 гг.
10. 1093/нар/7.6.1513
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
1093/нар/7.6.1513
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Каррутерс Л.М., Ширф В.Р., Демелер Б. и Хансен Дж. К. (2000) Анализ скорости осаждения макромолекулярных ансамблей. Методы Энзимол. 321, 66–80 10.1016/С0076-6879(00)21187-7 [PubMed] [CrossRef] [Google Scholar]
49. Schuck P. (2000) Анализ распределения макромолекул по размерам с помощью ультрацентрифугирования скорости седиментации и моделирования уравнения Ламма. Биофиз Дж. 78, 1606–1619. 10.1016/S0006-3495(00)76713-0 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Макде Р. Д., Ингланд Дж. Р., Йеннавар Х. П. и Тан С. (2010) Структура хроматинового фактора RCC1, связанного с сердцевиной нуклеосомы. Природа 467, 562–566 10.1038/природа09321 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51.
Олсон В.К., Горин А.А., Лу Х.Дж., Хок Л.М. и Журкин В.Б. (1998) Деформируемость, зависящая от последовательности ДНК, полученная из кристаллических комплексов белок-ДНК.